叢枝菌根真菌對汞脅迫下水稻葉片生理和光合特性的影響
2021-08-23 10:29:56李信茹蘇海磊米屹東王凡凡柳慧芳沈亞琴
環境科學研究 2021年8期
李信茹, 蘇海磊, 周 民,2, 米屹東,2, 王凡凡, 柳慧芳, 沈亞琴, 魏 源*
1.中國環境科學研究院, 環境基準與風險評估國家重點實驗室, 北京 100012 2.河海大學環境學院, 江蘇 南京 210098
中國是世界上最大的稻米生產和消費國,水稻種植面積可達 3 018.9×104hm2,總產量達2.1×108t/a,約占我國糧食總產量的1/3[1]. 提高水稻產量及其抗逆性對國家糧食安全保障具有重要意義. 汞是環境中毒性最強的重金屬之一,在自然界中主要以單質汞、無機汞和有機汞的形態存在,其中有機汞尤其是甲基汞的毒性最強[2]. 甲基汞和無機汞都具有很強的腎臟毒性、生殖系統毒性、免疫系統毒性和神經系統毒性[3]. 在水-土-大氣的環境介質中,即使是低劑量的汞也能通過生物放大效應沿食物鏈進入人體,嚴重危害人體健康[4]. 水稻對汞具有較強的富集能力,水稻特殊的淹水生長環境有利于硫酸鹽還原菌(SRB)等甲基化微生物將無機汞還原為甲基汞,極大地增加了汞在水稻中的毒性[5]. 研究表明,我國貴州萬山汞礦區內稻米中總汞含量為 0.002 8~0.295 5 mg/kg,平均值可達 0.050 8 mg/kg,甲基汞含量最高可達 0.019 0 mg/kg[6-7],均高于GB 2762—2005《食品中污染物限量》規定的糧食污染物標準限值(0.02 mg/kg). 食用稻米逐漸成為我國貴州、陜西和四川等汞礦區居民汞攝入的最主要途徑. 我國汞礦資源豐富,汞礦開采歷史悠久,對土壤尤其是西南礦區農田造成了嚴重的汞污染,因此,增強水稻對汞的耐受性,減少水稻籽粒中汞富集含量對減輕人體汞暴露風險具有重要意義.
AM真菌能與80%的陸生植物形成良好的共生體系[8]. AM真菌侵染植物后形成的菌絲體極大地增加了植物根系與土壤的接觸面積,促進了宿主植物吸收營養物質和水分[9-10],減少植物病害,提高植物抗病性,保證甚至促進植物在逆境中的生長發育[11-13]. 有研究表明,AM真菌能有效降低As、Cd對水稻生長的毒害作用[14-15],通過接種AM真菌,水稻生物量明顯增加,凈光合速率和葉綠素含量均有增加,水稻地上部重金屬含量明顯降低,有效降低了重金屬通過食物鏈傳遞的潛在危害.
目前有關AM真菌的研究大多集中在As、Cd等重金屬,有關Hg、水稻和AM真菌三者之間關系的研究較少. 為此,該文通過接種AM真菌進行溫室盆栽試驗,設置低、中、高3個Hg投加量梯度,選取接種和未接種AM真菌的水稻作為研究對象,測定水稻光合生理指標和抗氧化酶系指標,比較不同Hg投加量下AM真菌對水稻生長的影響,旨在探索AM真菌對不同Hg投加量下水稻生長的影響,以期為更好地將AM真菌應用在水稻生長及提高水稻抗逆性方面提供理論指導.
1 材料與方法
1.1 供試材料
所用水稻品種為鎮糯20,來自安徽省銅陵市義安區農業農村局的友情饋贈.
選用菌種為摩西球囊霉(BGC BJ05A)和幼套球囊霉(BGC XJ04B)混合菌種,購自北京市農林科學院植物營養與資源研究所,每10 g菌種中含282個孢子,接種菌劑為含有孢子和菌絲的菌根真菌及寄主植物根段的根際混合土壤,并按照10 g/kg AM真菌菌劑的比例進行接種.
土壤采自江蘇省常熟市某農田. 將土壤風干,過2 mm篩,高壓滅菌鍋121 ℃下滅菌2 h備用,土壤理化性質見表1. 試劑盒購自青島科創質量檢測公司.
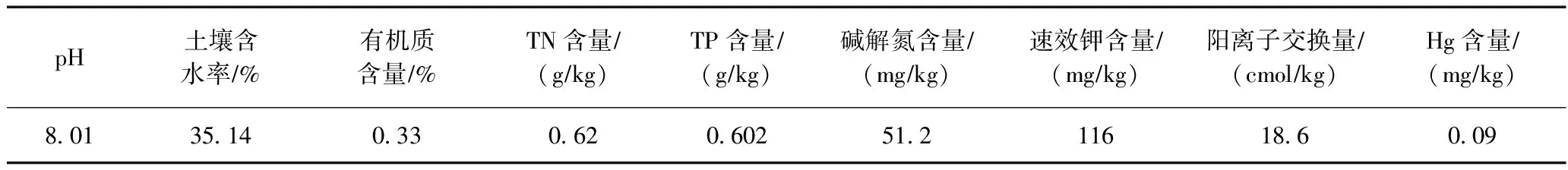
表1 供試土壤理化性質
1.2 試驗設計
試驗地點位于中國環境科學研究院溫室,溫度為22~30 ℃,濕度為50%~90%,恒溫箱人為光照(10 000 lx)12 h. 選擇顆粒飽滿的水稻種子,在氣候箱內進行恒溫育苗,育苗時間為20~30 d,育苗結束后,挑選長勢相近的水稻苗備用. 水稻苗移栽前用10%的過氧化氫滅菌,再用清水洗凈. 種植容器為高20 cm、直徑16 cm的不漏水圓形聚乙烯花盆,使用前用10%的過氧化氫及75%的酒精擦拭2~3次. 每盆裝2.5 kg土,提前加入配制好的氯化汞(HgCl2)老化1個月. 老化結束后,水稻移栽同時,向每盆接種組加入25 g AM真菌〔土樣(kg)與AM真菌(g)質量比為1∶10〕,AM真菌放置于土面之下10~15 cm處. 試驗共設置4個Hg投加量梯度,分別為0、0.1、1.0和2.0 mg/kg. 每個梯度分別設置AM真菌接種組和未接種組,每組各設置3組平行,共24盆,定期澆Hoagland營養液.
1.3 試驗指標測定
1.3.1菌根侵染率
水稻生長到第30天時取水稻根系進行菌根侵染率的測定. 采用曲利苯藍染色法測定菌根侵染率[16].
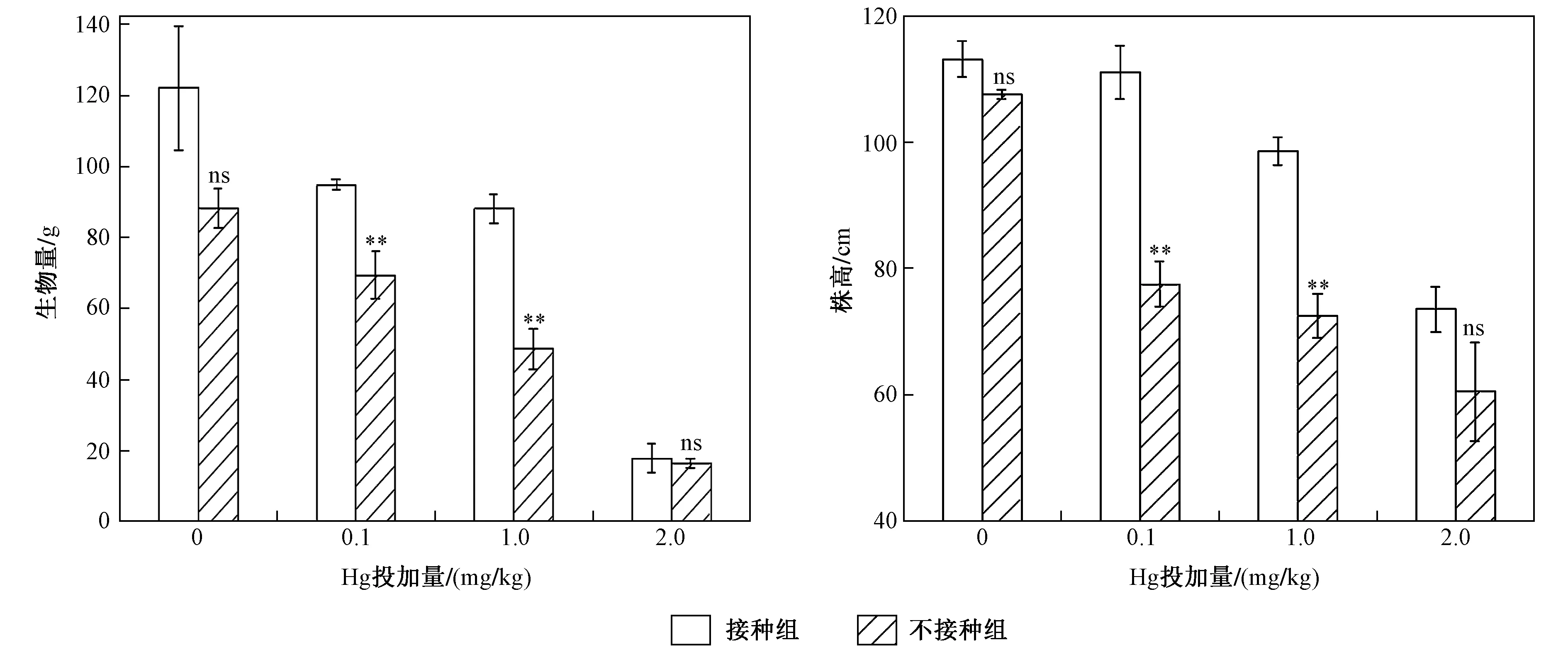
注: 數據為3次重復的平均值±標準偏差. ns表示無顯著差異,*為P<0.05,表示差異顯著;** 為P<0.01,表示差異非常顯著; *** 為P<0.001,表示差異極顯著. 下同.圖1 不同Hg投加量下接種AM真菌對水稻植物株高和生物量的影響Fig.1 Effects of rice biomass and height inoculation with/without AM fungi on under different Hg addition levels
1.3.2水稻株高、生物量的測定
水稻生長到第150天時,收獲水稻,并用卷尺測定水稻株高,70 ℃下烘干至恒質量,稱量水稻生物量.
1.3.3葉綠素相對含量測定及光合指標測定
水稻生長到第60天時采集水稻葉片進行水稻葉片葉綠素相對含量及光合指標的測定.
采用手持式葉綠素測定儀(托普TYS-B,浙江托普云農科技股份有限公司)測定葉片總葉綠素含量. 每盆水稻選取3片頂葉測定葉綠素相對含量(SPAD),重復測定4次,取平均值.
采用光合作用測定儀(托普3051D,浙江托普云農科技股份有限公司)測定凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、水分利用效率(WUE). 光合指標測定時間為10:00—12:00.
1.3.4水稻葉片抗氧化酶系及MDA含量測定
水稻生長到第60天時采集水稻葉片進行水稻葉片抗氧化酶系的活性及MDA含量的測定.
水稻SOD酶、POD酶、CAT酶、APX酶的活性及MDA含量均采用植物酶聯免疫分析法測定. 具體試驗步驟按酶試劑盒說明進行.
1.3.5水稻葉片可溶性糖和可溶性蛋白含量測定
水稻生長到第60天時采集水稻葉片進行水稻葉片可溶性糖和可溶性蛋白含量測定.
水稻葉片可溶性糖含量采用蒽酮法測定[17]. 水稻葉片可溶性蛋白含量采用考馬斯亮藍法測定[18].
1.4 數據處理方法
所有統計分析數據采用Origin 2018進行均值、標準誤計算及制圖;采用統計分析軟件SPSS 25對數據進行單因素方差分析.
2 結果與分析
2.1 AM真菌接種對水稻生長的影響
不同Hg投加量下AM真菌接種組均在一定程度上被侵染,當Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,接種組菌根侵染率分別為24.04%、19.45%、16.40%和15.54%. 結果顯示,AM真菌與水稻已形成共生體系,并通過這一共生體系對水稻產生直接或間接影響.
不同Hg投加量下水稻植株生物量與株高如圖1所示. 由圖1可見,在不同Hg投加量下接種AM真菌均可以提高水稻生物量及株高,當Hg投加量分別為0、0.1、1.0、2.0 mg/kg時,水稻生物量分別增加了38.66%、36.89.%、81.68%和8.51%,水稻株高分別增加了5.12%、43.23%、35.86%和22.49%. 顯著性差異分析結果表明,外源Hg投加量為0和2.0 mg/kg時,AM真菌對水稻株高和生物量并沒有顯著影響,而當Hg投加量為0.1和1.0 mg/kg時,AM真菌能顯著提高水稻生物量和株高.
2.2 AM真菌對不同Hg投加量下水稻光合生理特性的影響
如圖2所示,與未接種AM真菌組相比,接種AM真菌導致水稻葉片凈光合速率增加,且不同Hg投加量下接種組凈光合速率的增幅不同. 與對照組相比,Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,凈光合速率增幅分別為15.11%、48.18%、135.85%和134.88%,其中Hg投加量為1.0和2.0 mg/kg時凈光合速率增幅最大. 與未接種AM真菌組相比,接種AM真菌增加了水稻葉片氣孔導度,且不同Hg投加量下氣孔導度增幅不同. 與對照組相比,Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,氣孔導度增幅分別為40.00%、200.00%、133.33%和42.86%,其中Hg投加量為0.1 mg/kg時氣孔導度增幅最大. 與未接種AM真菌組相比,接種AM真菌導致水稻葉片水分利用效率增加,且不同Hg投加量下水分利用效率增幅不同. 與對照組相比,Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,水分利用效率增幅分別為76.93%、5.08%、35.12%和74.79%. 其中Hg投加量為0 mg/kg時水分利用效率增幅最大,Hg投加量為0.1 mg/kg時水分利用效率變化最小. 與未接種AM真菌組相比,接種AM真菌對水稻葉片胞間CO2濃度的影響各有不同. 其中,與未接種組相比,未添加Hg的接種組胞間CO2濃度有所增加,而在Hg脅迫條件下,水稻胞間CO2濃度均較未接種組有所降低,Hg投加量分別為0.1、1.0、2.0 mg/kg時,胞間CO2濃度降幅依次為14.58%、1.94%、9.86%.
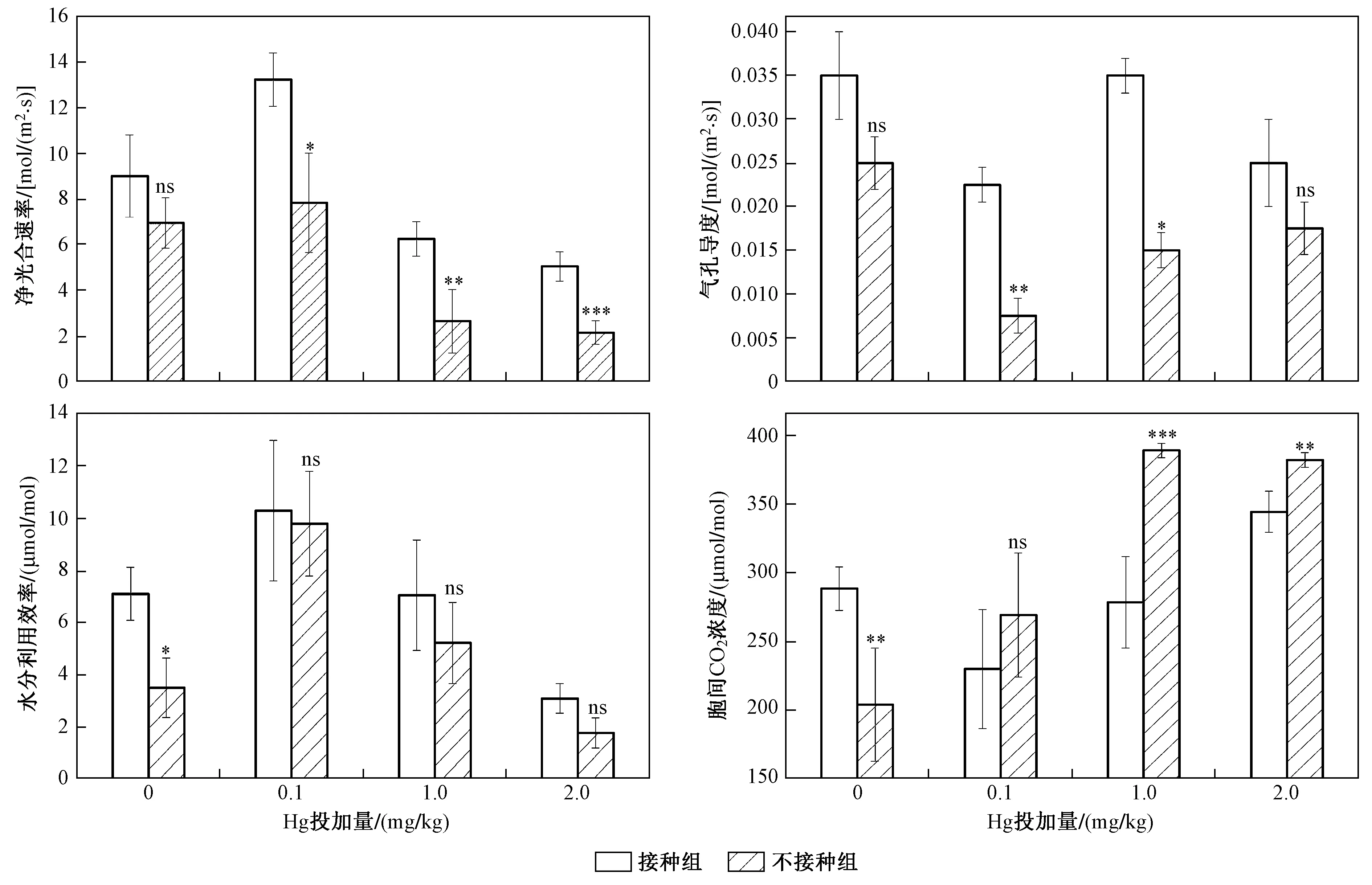
圖2 不同Hg投加量下接種和未接種AM真菌對水稻光合特性的影響Fig.2 The photosynthetic characteristics of rice inoculated with/without AM fungi under different Hg addition levels
對于葉綠素相對含量(SPAD),在不同Hg投加量下,AM真菌均能增加水稻葉片葉綠素相對含量,當Hg投加量為0、0.1、1.0和2.0 mg/kg時,葉綠素相對含量增幅依次為8.17%、28.79%、17.92%和12.96%. 當Hg投加量為0.1 mg/kg時,接種AM真菌后,水稻葉綠素相對含量增幅最大(見圖3),說明AM真菌極有可能通過促進水稻葉片光合色素的分泌來促進光合速率.
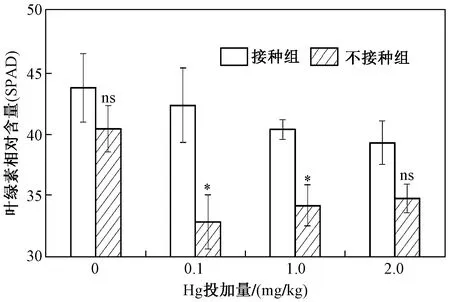
圖3 不同Hg投加量下接種和未接種AM真菌對水稻葉綠素相對含量的影響Fig.3 The chlorophyll content of rice inoculated with/without AM fungi under different Hg addition levels
2.3 AM真菌對不同Hg投加量下水稻葉片抗氧化酶活性和膜系統的影響
在不同Hg投加量下接種AM真菌對水稻葉片抗氧化酶活性的影響如圖4所示. 由圖4可見,與未接種組相比,接種AM真菌能提高水稻葉片SOD活性,且不同Hg投加量下接種AM真菌對水稻葉片SOD活性的影響不同. 其中,Hg投加量為0.1 mg/kg時,SOD活性增幅最大,為19.06%,其次是Hg投加量分別為1.0、0和2.0 mg/kg時,增幅依次為8.24%、1.07%、1.00%. 與未接種組相比,接種AM真菌能提高水稻葉片POD活性,且不同Hg投加量下接種AM真菌對水稻葉片POD活性的影響不同. 其中,Hg投加量為0 mg/kg時AM真菌能顯著增加POD活性,增幅為15.82%,Hg投加量為0.1 mg/kg時水稻葉片POD活性增加最少,增幅為2.17%,當Hg投加量分別為1.0和2.0 mg/kg時,POD活性增幅依次為5.97%和11.88%. 與未接種組相比,接種AM真菌能提高水稻葉片CAT活性,但增長差異不明顯,Hg投加量為0、0.1、1.0和2.0 mg/kg時,水稻葉片CAT活性增幅依次為4.28%、3.00%、9.96%和4.73%. 對于APX,接種AM真菌能顯著提高水稻葉片APX活性,當Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,與未接種組相比,接種AM真菌時水稻葉片APX活性的增幅依次為33.03%、11.05%、13.45%和23.58%,其中,Hg投加量為0.1 mg/kg時,APX活性增幅最少,Hg投加量為0 mg/kg時增幅最大.

圖4 不同Hg投加量下接種和未接種AM真菌對水稻抗氧化酶系的影響Fig.4 The antioxidant enzyme system of rice inoculated with/without AM fungi under different Hg addition levels
對于MDA,接種AM真菌能降低水稻葉片MDA含量,當Hg投加量分別為0、0.1、1.0和2.0 mg/kg時,與未接種組相比,接種AM真菌時水稻葉片MDA含量的降幅依次為10.97%、10.39%、1.55%和2.17%,其中Hg投加量為1.0和2.0 mg/kg時,其降幅最小,未添加Hg的試驗組和Hg投加量為0.1 mg/kg時的降幅最大(見圖5). 抗氧化酶系活性能表征水稻葉片細胞受重金屬毒害作用的強弱,一般來說,重金屬脅迫下,植物葉片氧化還原穩態被破壞,為了抵御這一毒害作用,水稻抗氧化酶系活性會增加,以緩解已被破壞的氧化還原穩態. 該研究證實,AM真菌能促進抗氧化酶系活性增加,繼而緩解細胞毒害,調節葉片細胞體內氧化還原穩態.
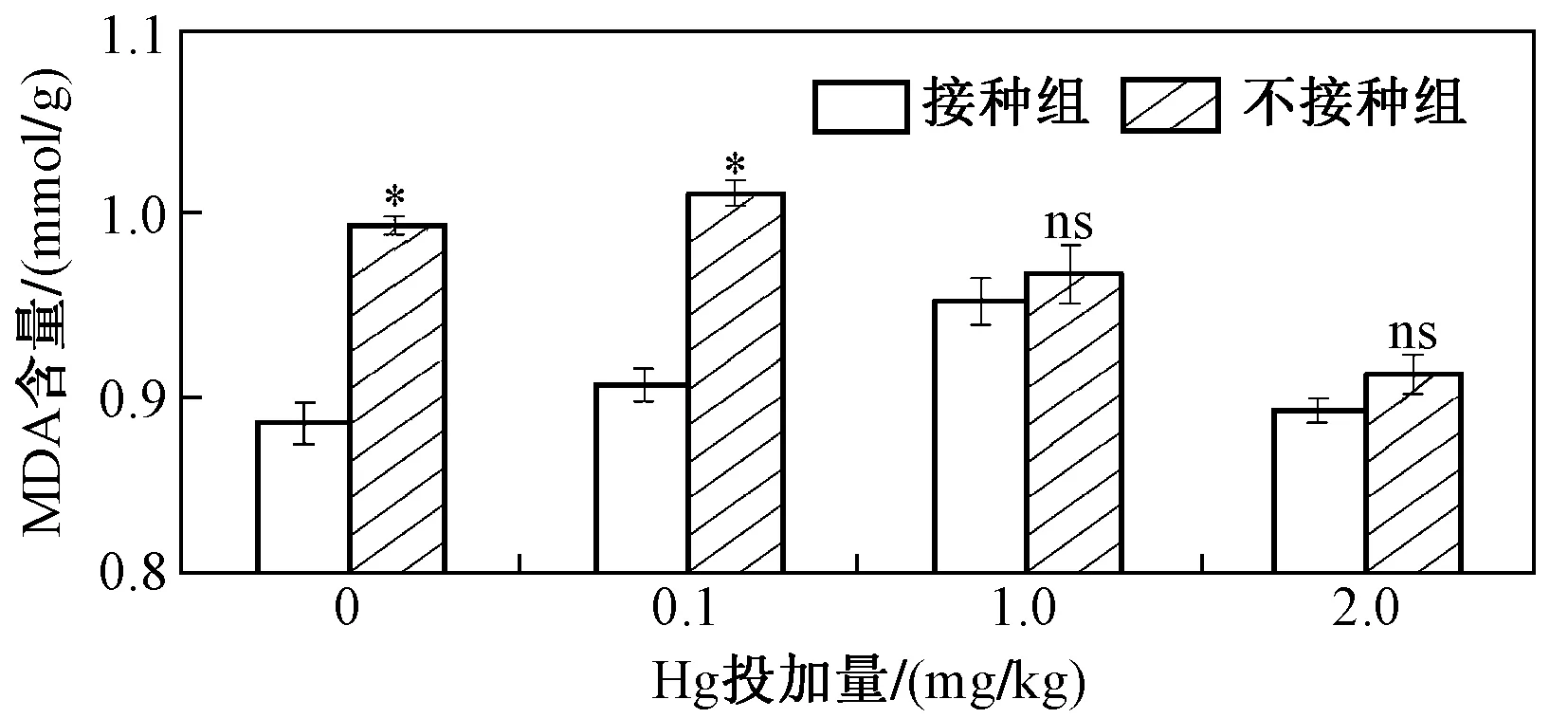
圖5 不同Hg投加量下接種和未接種AM真菌對水稻MDA含量的影響Fig.5 The content of MDA of rice inoculated with/without AM fungi under different Hg addition levels
2.4 AM真菌對不同Hg投加量下水稻葉片可溶性糖和可溶性蛋白的影響
如圖6所示,Hg投加量下接種AM真菌對水稻葉片可溶性糖含量有一定影響,當Hg投加量為0 mg/kg 時,AM真菌降低了水稻葉片可溶性糖含量,在外源添加Hg后,AM真菌均能促進水稻分泌可溶性糖. 接種AM真菌條件下,Hg投加量分別為0.1、1.0和2.0 mg/kg時,水稻可溶性糖含量增幅依次為43.14%、60.69%和21.21%. 同時,不同Hg投加量下,AM真菌對水稻可溶性蛋白含量的影響各有不同. 當Hg投加量為0、1.0 mg/kg時,AM真菌能促進水稻分泌可溶性蛋白,而當Hg投加量為0.1和2.0 mg/kg時,AM真菌能抑制水稻葉片分泌可溶性蛋白. 不同Hg投加量下,AM真菌對水稻可溶性糖和可溶性蛋白含量的影響不同,影響程度也各異,這可能與Hg投加量相關[19],水稻本身對外界脅迫具有一定的適應能力,這一適應能力與水稻自身的抗性及外界脅迫強弱相關,因此,水稻可溶性糖和可溶性蛋白可能對Hg的濃度更為敏感,在Hg脅迫下,水稻葉片可溶性糖和可溶性蛋白含量的影響需要更加關注Hg濃度的影響.
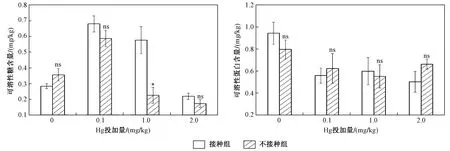
圖6 不同Hg投加量下接種和未接種AM真菌對水稻可溶性蛋白和可溶性糖的影響Fig.6 The content of soluble proteins and soluble sugars of rice inoculated with/without AM fungi under different Hg addition levels
3 討論
3.1 AM真菌對不同Hg投加量下水稻生物量和株高的影響
植物生物量和株高是植物生長狀況評價最直觀的指標,能夠直接反映植物對逆境的抗性. 該研究發現,在各Hg投加量下,接種AM真菌均增加了水稻生物量和株高,這與Alam等[20]的研究結果一致,他們發現在砷污染土壤中接種AM真菌能增加小扁豆生物量. 研究證實,AM真菌可顯著促進Hg脅迫下水稻的生長,其機制可能是AM真菌與水稻共生體形成后,數量繁多的根外菌絲極大地擴大了植物根系表面,有利于水稻根系從土壤根系吸收水分、養分等營養物質[21].
3.2 AM真菌對不同Hg投加量下水稻光合生理特性的影響
植物光合作用與植物體內有機物的積累情況息息相關,一般來說,植物體內的有機物積累量越多,說明植物光合作用越強,植物將CO2同化為有機物的能力就越強[22]. 一般認為,影響葉片光合作用的因素分為氣孔因素和非氣孔因素[23]. 氣孔因素是指由于氣孔導度降低、CO2進入葉肉細胞受阻,從而導致葉肉細胞光合速率降低的現象[24]. 當光合速率降低、胞間CO2濃度降低且氣孔導度降低時,氣孔因素是影響葉片光合作用的主要因素[25];反之,當光合速率降低、胞間CO2濃度升高或不變、氣孔導度升高時,非氣孔因素是影響葉片光合作用的主要因素. 該研究發現,在未接種AM真菌的條件下,當Hg投加量大于0.1 mg/kg時,水稻葉片光合速率、氣孔導度均隨Hg投加量的增加而逐漸降低,氣孔導度隨之升高,說明此時影響水稻葉片光合速率的最主要因素是非氣孔因素. 接種AM真菌后,同樣是Hg投加量大于0.1 mg/kg 時,水稻葉片光合速率、胞間CO2濃度和氣孔導度均隨Hg投加量的增加而逐漸降低,說明由于氣孔導度降低,CO2進入葉片受阻,繼而影響葉片光合作用,此時影響葉片光合作用的最主要因素是氣孔因素. 同時,該研究發現,接種AM真菌后,水稻葉片光合速率、水分利用效率均呈先增后減的趨勢,并在Hg投加量為0.1 mg/kg時凈光合速率最高,這與孫永娣等[26]的研究結果一致,他們發現,隨著Hg投加量的升高,茅蒼木的光合速率也呈先增再減的趨勢,并在Hg投加量為1.0 mg/kg時達到峰值. 這說明低濃度的Hg能促進植物光合速率的增加[27].
與未接種組相比,接種組水稻葉片光合速率、氣孔導度和水分利用效率均增加,說明AM真菌能促進水稻同化CO2生成有機物,增強水稻抗逆性. 有研究[28-29]表明,Hg脅迫條件下,植物葉片葉綠素含量會明顯降低. 隨著Hg投加量的增加,水稻葉綠素含量逐漸降低,接種AM真菌后,水稻葉綠素含量明顯增高,結合顯著性差異分析結果可知,當Hg投加量為0.1和1.0 mg/kg時,AM真菌能顯著影響水稻葉綠素相對含量,說明AM真菌能通過促進光合色素分泌來提高水稻葉片光合速率. 在復合污染場地上,AM真菌同樣具有較高的提高植物抗逆性的能力,王宇濤等[30]發現,在Cd、Cu復合污染下,接種AM真菌能提高玉米的株高、生物量以及葉綠素含量,說明AM真菌提高植物的抗逆性的方式之一可能是增加葉綠素含量,緩解重金屬對光合作用的抑制[31].
3.3 AM真菌對不同Hg投加量下水稻抗氧化酶系的影響
MDA是膜質過氧化最重要的產物,通過MDA可以了解膜質過氧化的程度,以間接測定膜系統受損程度和植物抗逆性. 該研究發現,Hg脅迫條件下,接種AM真菌均降低了水稻葉片MDA含量,說明AM真菌能有效緩解Hg對細胞膜系統的毒害作用,增加植物抗逆性. 這與彭昌琴等[32]的研究結果一致,他們發現,AM真菌接種組及未接種組中鳳仙花中MDA的含量基本都隨著Cd濃度的增加而增加,但在同一Cd濃度下,AM真菌接種能降低MDA含量,說明AM真菌在每種Cd濃度下均能降低植物細胞膜質過氧化作用,繼而緩解重金屬毒害.
Hg、Cd等重金屬會促進植物體內活性氧的產生,破壞植物細胞氧化還原穩態,最終導致細胞損傷[33-34]. 抗氧化酶系能清除細胞內多余的活性氧,恢復細胞穩態,通過測定抗氧化酶活性,能了解植物細胞所受到的重金屬毒害作用強弱. 沈亞琴等[35]以玉米為研究對象,同樣研究了Sb脅迫下玉米的生長及抗氧化酶系活性,結果表明,SOD、POD、CAT的活性較未接種組均顯著增加,單因素方差分析結果顯示,AM真菌對玉米SOD、POD、CAT的活性均有顯著影響. 同樣,該研究結果證實,AM真菌能有效增加水稻葉片SOD、POD、CAT、APX的活性,說明AM真菌能通過提高植物抗氧化酶系活性來緩解重金屬毒害作用,進而提高植物抗逆性. 顯著性差異結果分析表明,不同Hg投加量下,AM真菌對水稻抗氧化酶活性的影響更為顯著,說明AM真菌更有可能通過提高抗氧化酶活性提高植物抗逆性. Neera等[36]研究表明,在Cd污染條件下接種AM真菌,毛竹抗氧化酶系活性有所增加,且隨著Cd濃度的增加,AM真菌對毛竹抗氧化酶系活性影響更為顯著. 陳志鵬等[37]以紫花苜蓿為研究對象,通過接種AM真菌,研究Sb脅迫下紫花苜蓿的生長狀況,結果表明,不同Sb濃度下AM真菌對抗氧化酶系的活性影響各有不同,說明AM真菌對重金屬Sb的作用受Sb濃度的影響,這與筆者所得結果類似,盡管AM真菌促進了水稻抗氧化酶系活性的增加,但是在不同Hg投加量下,AM真菌對水稻抗氧化酶系活性影響的顯著性各有不同,因此,AM真菌對水稻的抗氧化酶系活性影響是否存在某一最適汞濃度仍需進一步分析.
3.4 AM真菌對可溶性糖和可溶性蛋白的影響
植物體內的可溶性蛋白大多數是參與代謝的酶類,是用來了解植物體總代謝的重要指標[38],可溶性蛋白能幫助植物維持細胞較低的滲透勢,增加細胞的持水能力,這一特性在低溫及干旱條件下更為明顯[39-41]. 可溶性糖是主要的滲透調節物質,能夠提高細胞膜和原生質體的穩定性,并在植物細胞體內無機離子濃度過高時起到保護酶類的作用[42-43]. 趙匠等[44]研究了鹽脅迫條件下AM真菌對黃檗的影響,結果顯示,AM真菌能通過促進黃檗可溶性蛋白分泌來提高黃檗抗逆性. 孫晨瑜等[45]的研究也發現,AM真菌的施加能有效增加植物機制根系分泌物中可溶性糖和可溶性蛋白含量,而水培條件下這一促進作用并不明顯,這可能因為AM真菌是好氧微生物,水培條件下AM真菌生長及侵染情況有待進一步研究,結合筆者研究中AM真菌對水稻可溶性糖和可溶性蛋白的影響并不顯著這一結論進行分析,表明水稻的淹水環境對AM真菌生長確實產生了一定影響,但影響程度還需要做進一步研究.
4 結論
a) 在不同Hg投加量下,接種AM真菌均能有效增加水稻株高及生物量,當Hg投加量為0.1 mg/kg時,水稻株高和生物量較未接種組的增幅最大.
b) 在不同Hg投加量下,接種AM真菌能有效提高水稻光合速率. 接種AM真菌后,當Hg投加量大于0.1 mg/kg時,影響水稻葉片光合作用的因素從非氣孔因素轉變為氣孔因素. 在Hg投加量為0.1 mg/kg時,接種及不接種AM真菌條件下,水稻葉片光合速率均最強,表明水稻對低濃度Hg具有一定耐受性.
c) 隨著Hg投加量的增加,接種AM真菌條件下水稻抗氧化酶系活性均有所增加. 接種AM真菌能通過提高抗氧化酶系活性來降低細胞內活性氧含量,繼而維持細胞穩態,增強水稻對Hg的抗逆性.
d) 接種AM真菌能有效增加水稻可溶性蛋白和可溶性糖含量、降低MDA含量. 在Hg投加量為1.0 mg/kg時,接種AM真菌對水稻可溶性糖含量的促進作用最明顯,而可溶性蛋白含量的改變趨勢并不明顯. 當Hg投加量為0.1 mg/kg時,MDA含量的降幅最明顯,表明接種AM真菌能減少細胞質膜損傷,降低重金屬對水稻植物的毒害作用.
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
