HA蛋白位點變異影響H7N9亞型流感病毒特性的研究進展
2021-08-23 07:00:38嚴雅瑤劉秀梵
畜牧獸醫學報 2021年8期
嚴雅瑤,顧 敏,2,3 *,劉秀梵,2,3
(1.揚州大學農業部畜禽傳染病學重點開放實驗室,揚州 225009;2. 江蘇高校動物重要疫病與人獸共患病防控協同創新中心,揚州 225009; 3.江蘇省人獸共患病學重點實驗室,揚州 225009)
2013年3月,一種源自禽類的新型H7N9亞型流感病毒在人群中暴發開來,迄今為止,已出現至少五波流行高峰。其中,2016—2017年的第五波疫情出現最早、報告的人感染病例數最多、區域分布也最廣,并且在此期間還分離到了對雞呈高致病性的H7N9禽流感病毒[1-3]。截至2021年7月7日,全球共報告1 568人感染H7N9病毒,死亡616例,患者數量超過了H5N1、H9N2等其他能夠感染人的禽流感病毒亞型的總和[4]。同時,H7N9病毒仍在不斷地變異,例如與家禽群體中的H9N2、H5N6亞型病毒進一步發生基因重配而產生新型H7N2、H7N6病毒,對家禽養殖及公共衛生安全構成持續威脅[5-6]。自2017年下半年以來,伴隨著特異性禽流感疫苗的有效使用,H7N9亞型禽流感病毒的分離率急劇下降;相對應地,大多通過活禽或活禽市場暴露而感染的人的散發病例也驟減[7-8]。然而,盡管當前對家禽低致病性的H7N9病毒已基本分離不到,但高致病性H7N9禽流感病毒仍在我國部分地區存在,且部分毒株表現出明顯的抗原性變異,使防控工作面臨嚴峻挑戰[9-10]。此外,雖然目前H7N9病毒尚不具備在人際間有效傳播的能力,但病毒有可能通過關鍵基因位點的突變或基因片段的重配進而提高其在人類宿主中的適應性,所以密切監測與病毒生物學特性相關的氨基酸變異尤為重要。
1 H7N9亞型流感病毒概述
流感病毒屬于正黏病毒科,是具有囊膜、分節段的負鏈RNA病毒,大小為80~120 nm,根據其核蛋白(NP)和基質蛋白(M)的抗原性不同,可分為A、B、C、D 4個型[11]。其中,威脅畜禽和人類健康最為嚴重的是A型流感病毒(influenza A virus, IAV),其基因組包含8條獨立的基因片段,總長度約13 kb,可編碼多達17種蛋白,包括人們所熟知的10種核心蛋白(PB2、PB1、PA、HA、NP、NA、M1、M2、NS1、NS2),還有近年來陸續發現的附屬蛋白(accessory proteins),如:PB1-F2、PB1-N40、PA-X、M42、NS3、PA-N155和PA-N182[12]等。上述蛋白中,HA與NA是IAV囊膜表面最重要的兩種抗原性糖蛋白,在IAV的生命周期中發揮重要作用:HA主要介導病毒與宿主細胞的結合及包裝復制,NA主要負責切割受體結合、釋放病毒粒子,兩者維持著功能上的動態平衡,同時HA和NA也是進一步劃分H1~H18、N1~N11亞型的依據[13]。
在眾多IAV亞型中,H7N9因其同時對畜禽養殖與公眾健康造成了嚴重危害而備受關注。該始于2013年的新型H7N9病毒,推測其可能是H7、N9和H9N2亞型禽流感病毒的三源重配產物:H7和N9基因為歐亞進化譜系的野鳥來源,而內部片段則完全由中國家禽中廣泛流行的H9N2提供,三者重組誕生了原始型H7N9,而后進一步在家禽中暴發開來,并伴隨有人類感染病例的出現[14-15]。在地區分布上,H7N9病毒首次出現在長江三角洲及珠江三角洲等東南沿海地區,隨著病毒的傳播蔓延,近幾年西部內陸及東北地區也陸續出現感染病例[10,16]。在時間上,H7N9病毒自出現以來,每年冬季至次年春季均會在人群中形成一次流行高峰,截至目前已引起至少五波流行,其中以第五波(2016—2017年)的感染人數最多、感染區域最廣。同時,第五波流行期間還出現了對家禽呈高致病性(highly pathogenic,HP)的H7N9病毒,其HA裂解位點處早期主要表現為 “KRTA”的特異性氨基酸插入,雖保留與此前低致病性(low pathogenicity,LP)H7N9病毒相類似的雙受體結合特性,但可能在哺乳動物中表現出更強的致病性、傳播性以及更廣泛的組織嗜性[1,7]。并且,不斷有證據表明,HP H7N9仍在持續地演化與變異。例如,H7N9感染的禽類宿主范圍在不斷擴大,除雞以外,鴨、鵪鶉體內也分離到少量H7N9毒株[8,17-18]。HP H7N9病毒還可以與其他NA亞型的禽流感病毒發生基因重配而產生H7N2、H7N3、H7N6等新型重組H7Ny病毒[19]。另外,HA蛋白上關鍵氨基酸位點的變異及其導致的N-糖基化(N-linked glycosylation, NLG)狀態改變能夠通過對蛋白的空間構象與生物活性的調節來影響H7N9病毒的生物學特性[20-21]。
2 HA蛋白的結構特點
2.1 HA的一級結構
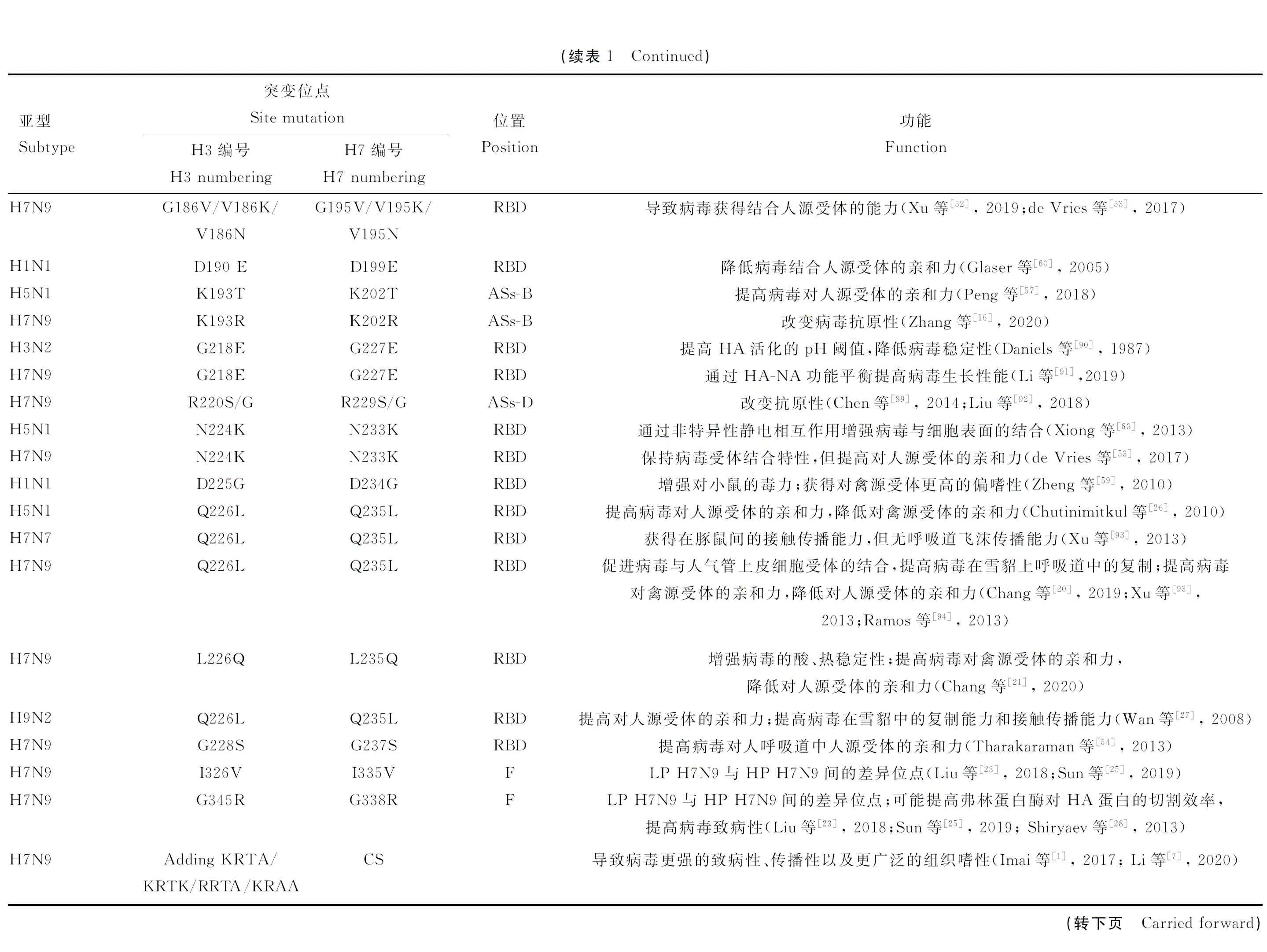
HA蛋白位于IAV囊膜表面,由3個HA1-HA2復合物單體結合而成同源三聚體。各亞型HA蛋白在結構上大體相似,但一級結構的片段長度略有不同。H7N9病毒的HA單體由561或565個氨基酸組成,從N端開始的18個疏水性氨基酸組成信號肽,緊接著321或325個氨基酸殘基組成HA1,隨后由222個氨基酸構成HA2[22](圖1A)。對于HA2,其N端融合肽(fusion peptide, FP)區域為疏水性氨基酸,可與病毒囊膜的脂質雙層緊密相連;緊接著仍由一系列疏水性氨基酸構成跨膜區;而C末端的氨基酸多數為親水性,可透過脂質雙分子層進入病毒粒子內部而構成胞內尾區。序列分析發現,LP和HP H7N9病毒HA基因長度的差異由裂解位點處(cleavage site,CS)氨基酸基序的差異所導致,前者為PKG----R↓G,而后者至少存在7種模式:PKRKRTAR↓G、PKGKRTKR↓G、PKGKRTAR↓G、PKGKRIAR↓G、PKRRRTAR↓G、PKRKRAAR↓G、PKRKRIAR↓G,其中占比較高的為PKG/RKRTAR↓G[2, 23-24]。此外,相較于LP H7N9,HP H7N9病毒的HA上還有諸如HA1中L235Q(H3編號為L226Q)、G338R等明顯氨基酸替換[25]。其中,235位是重要的抗原表位并能夠影響LP與HP H7N9病毒的交叉反應性,且在其他多個HA亞型中均被證實與流感病毒的跨種傳播能力密切相關[21,26-27]。而G338R突變增加了裂解位點處連續堿性氨基酸的數目,可以潛在地提高弗林蛋白酶的切割效率,從而對病毒的感染性及致病力造成影響[23,28]。
A. H7N9病毒HA蛋白的一級結構:圖中氨基酸計數使用H7編號(包含信號肽),裂解位點處HP H7N9相較于LP H7N9存在“KRTA”的4個氨基酸插入;B. H7N9病毒HA蛋白的三維結構:圖中僅顯示HA單體結構,并標記出影響病毒生物學特性的部分關鍵氨基酸位點;該結構以A/Shanghai/02/2013(H7N9) 病毒的HA蛋白結構(PDB ID: 6IDD)為模板進行同源建模,使用PyMOL軟件進行展示A. The primary structure of HA protein of H7N9 virus. The amino acid sites were according to H7 numbering, including the signal peptide. As compared with LP H7N9, HP H7N9 possessed the 4-amino-acid insertion of KRTA at the cleavage site; B. The three-dimensional structure of HA protein of H7N9 virus. The structure of HA monomer was shown, and some key amino acid sites affecting the viral biological properties were labelled. The structure was homology modelled with the HA protein template of A/Shanghai/02/2013(H7N9) virus (PDB ID: 6IDD), and presented with the software of PyMOL圖1 H7N9亞型流感病毒HA蛋白的結構Fig.1 HA protein structure of H7N9 subtype influenza virus
2.2 HA的空間結構
HA是一種典型的I型蛋白,其三聚體組裝成一個中心螺旋線圈,形成桿狀莖部以及含有唾液酸(sialic acid, SA)結合位點的球狀頭部,三聚體高約135 ?,直徑35~70 ?[29]。頭部由HA1的大部分組成,變異率較高,包含受體結合區(receptor binding domain, RBD)、痕酯酶區(vestigial esterase domain, VE)。莖部較為保守,由HA1的N端、C端部分和HA2的整體共同組成,且一直延伸至膜內(圖1B)。
2.2.1 頭部(head) HA蛋白的頭部是由HA1第53—275位氨基酸(H3編號)折疊成8條反平行鏈而組成的空間構象,其遠端是RBD區,130-loop(135~138)、190-helix(190~198)和220-loop(221~228) 這3個二級結構元件構成口袋邊緣;保守性的氨基酸殘基Y98、W153、H183和Y195組成了基底部;鄰近的150-loop作為RBD結構的一部分,有助于保持結構完整性及與SA形成相互作用[30-31]。各HA亞型的RBD構象較為相似,但也存在一定差異,比如H7亞型的RBD相較于H1、H3、H5等亞型,在150-loop中存在兩個連續氨基酸(Asn、Thr)的插入,使150-loop突出到結合位點的一側,而這種構象有可能影響病毒受體結合特異性[22,32]。此外,一些H7亞型病毒(如H7N1)在150-loop的第158位新增一個NLG修飾,形成空間干擾從而阻礙受體的結合,最終降低病毒對受體的親和力[33]。而其他亞型(如H5N6)HA第158 位出現NLG缺失后,卻能夠獲得結合人源受體的能力,并且增加對小鼠的致病性[34]。因此,RBD的氨基酸差異及NLG修飾對流感病毒受體結合特性具有較大影響。
位于HA1的RBD和HA2的膜近端莖部之間的則是VE區,因其與C型流感病毒脂酶融合蛋白上的9-O-乙酰脂酶同源性較高而得名[35],具體定位于第53—115及265—275位氨基酸(H3編號)(圖1B)。盡管有關VE區的功能研究尚較少,但其高度保守的特性可作為除HA頭部、莖部之外的第3個用于治療性單抗結合的靶點區域,目前已在H5N1病毒的相關研究中取得一定成效[36]。
同時,HA頭部具有較多抗原表位(antigenic sites, ASs)。通常所述的5大抗原區(H3編號)主要包括:位于膜遠端的A區(133—137,140—146)和B區(156—160,187—198),位于膜近端的C區(53、54、275、278),位于HA三聚體交界處的D區(174、207)及位于VE結構域的E區(63、78、81、83);其中A、B抗原區因與RBD的部分區域(A區與130-loop、B區與190-helix)存在重疊而導致其抗原性相對于C、D、E區有所增強,此外,A、B、E區的抗原位點多為線性表位,而C、D區的抗原位點多呈構象表位[37]。但HA抗原位點處氨基酸的組成及數量并非一成不變,處于上述5個抗原區之外的氨基酸變異,比如G132R、G137E(H3編號分別為G124R、G129E),同樣可能引起抗原性的改變而使其成為新的潛在抗原位點[38]。
2.2.2 莖部(stem) HA莖部由部分HA1和全部HA2組成,其膜外螺旋樣結構支撐膜遠端的HA頭部,并延伸至膜內從而將HA蛋白錨定于病毒囊膜表面。膜外的HA莖部也包含較多抗原位點且在各HA亞型中高度保守,因此可作為中和抗體結合的靶點,但各亞型莖部之間也具有一定相似性,如group 1的H5、H9,group 2的H3、H7,故部分抗體(如CR6216)可以中和group 1中的某些病毒、部分抗體(如CR8020)可以中和group 2中的某些病毒或對group 1和group 2病毒的攻擊均具有保護作用(如C05)[39-41]。這些靶向HA莖部的抗體通過抑制病毒囊膜與細胞膜的融合來阻斷病毒遺傳物質進入宿主細胞內,從而抑制病毒的復制。此外,位于膜近端長螺旋上的D112G(HA2編號)會降低HA蛋白的穩定性,而位于膜遠端短螺旋上的K58I(HA2編號)則會增加HA蛋白的穩定性,這些突變可為開發穩定性更好的疫苗提供理論參考[42]。HA莖部的跨膜區和胞內區更為保守,其中的半胱氨酸可作為重要的酰化位點與棕櫚酸結合,而經修飾后的棕櫚酰化HA蛋白對于膜融合及病毒出芽均至關重要[43-44]。同時,莖部還存在NLG位點,具有高度保守性及一定的亞型特異性;它們對于HA蛋白的折疊和構象至關重要,當去除莖部關鍵的NLG位點會導致HA蛋白的三聚化、折疊及轉運受損,同時還會改變HA對pH變化的敏感性[45-47]。
3 HA蛋白的主要功能
3.1 HA蛋白RBD介導的受體結合特性
HA蛋白的受體結合特性是影響流感病毒組織嗜性及宿主范圍的主要因素,而SA是存在于宿主細胞表面與病毒感染相關的重要結構。SA作為一類9-碳單糖神經氨酸衍生物,常存在于N-聚糖、寡聚糖和神經節苷脂的末端。在自然界中,SA通常通過其C5位的氨基與乙酰基或羥乙酰基的酰化反應來穩定其修飾,從而分為N-乙酰神經氨酸 (N-acetylneuraminic acid, Neu5Ac)與N-羥乙酰神經氨酸 (N-glycolylneuraminic acid, Neu5Gc)[48]。此外,末端SA(SA-1)的C2與其鄰近的倒數第二個半乳糖(Gal-2)的C3或C6連接,分別生成SA-α2, 3-Gal和SA-α2, 6-Gal 兩種SA受體,這種SA空間構象的差異直接影響到流感病毒受體結合的偏嗜性[49]。
不同流感病毒對特異性SA受體的偏嗜性明顯不同,人流感病毒更偏向于識別SA-α2, 6-Gal,禽流感病毒和馬流感病毒親嗜SA-α2, 3-Gal,而豬流感病毒對α2, 3和α2, 6兩種SA受體親和力均較高[50]。同時,不同種屬動物受體的結合構象也有所差異,禽類受體的結合在SA-1和Gal-2之間呈現反式構象,而人源受體的結合均采用順式構象;這種受體結合的特異性,部分由HA蛋白受體結合口袋的結構特征決定,是影響宿主嗜性的重要影響因素,包括種間適應及傳播[51]。研究指出,HA蛋白受體結合口袋結構的細微變化即可導致受體結合偏好性的改變,比如,H7N9亞型HA蛋白上186、193、226、228位(H3編號)是病毒獲得結合SA-α2, 6-Gal的關鍵[52-54]。而其他亞型HA上的81、98、145、190、193、222、224、225、226、228和238位(H3編號)氨基酸也被報道可能通過影響RBD口袋區域的結構而影響其與SA受體的結合力及偏嗜性[53,55-63]。
3.2 HA蛋白莖部介導的雙膜融合
HA蛋白的另一重要功能是在低pH環境下誘導流感病毒囊膜與宿主細胞膜的融合,這也是病毒感染細胞過程中進行脫殼所必須的。首先,HA蛋白以未經修飾的HA0(HA前體蛋白)形式存在,雖能識別宿主細胞表面的受體,但不能與宿主細胞膜發生融合,導致病毒也不具備感染性[64-65]。而IAV發生感染的先決條件則是HA的可裂解特性,即HA0裂解為HA1和HA2,因此HA CS處的氨基酸組成和宿主體內的蛋白酶分布將對病毒的致病性和組織嗜性產生重要影響[65]。HP病毒在HA CS處具有連續的多堿性氨基酸,該序列特征在高爾基體反面網絡(trans-Golgi network,TGN)上被宿主體內泛嗜性的蛋白酶(如弗林蛋白酶、PC6)所識別并發生裂解[66-67]。相比之下,LP病毒在HA CS處通常僅含單個堿性氨基酸,主要由局限于機體特定部位(如呼吸道和胃腸道的黏膜表面)的胰蛋白酶樣蛋白酶所切割[67-69]。當HA RBD與SA受體結合后,經切割產生的具有膜融合能力的HA1-HA2復合物便通過內吞作用進入細胞。在內小體低pH環境下,HA蛋白經歷一系列構象變化,具體包括HA1-HA2構象改變、融合肽的暴露、莖部螺旋的重塑;其中,融合肽暴露后與跨膜區形成鉤狀結構插入宿主細胞膜中,拉近其與病毒囊膜的距離,從而介導雙膜融合和融合孔的形成,最終病毒的遺傳物質得以釋放到宿主細胞中實現病毒的轉錄和復制[70-72]。由于誘導雙膜融合主要由HA莖部介導,其高度保守的氨基酸序列維持著HA蛋白的穩定性及病毒的包裝復制。有研究表明,H3N2亞型F63P及F70P雙突變(HA2 編號)可通過影響HA莖部構象進而抑制雙膜融合[73]。此外,H7N1亞型12 N、28 N(H7編號,去信號肽)被證明是HA蛋白融合活性的關鍵位點,其變異會影響HA蛋白的胞內轉運乃至病毒的感染特性[47,73-74]。
3.3 HA蛋白的酸、熱穩定性
HA蛋白的酸、熱穩定性具體表現為病毒在不同pH、溫度等環境下抵抗失活的能力,是影響IAV宿主范圍、傳播性及致病潛力的重要因素[75]。其中,溫度作為一種非生理性誘發因素可允許HA蛋白在中性環境下發生雙膜融合;而一定pH閾值所觸發的HA構象改變(HA的活化)則通常定義為HA的酸穩定性,且有研究表明HA蛋白酸穩定性具有一定的宿主差異性及亞型特異性[76]。例如,人的H1N1、H2N2、H3N2大流感病毒pH閾值(5.1~5.5)普遍低于野鳥源IAV(5.5~6.1),而致人感染的禽流感病毒(如H5N1、H5N6、H7N9)pH閾值則普遍較高(5.6~5.8),提示較高的HA活化pH閾值(即HA低穩定性)有利于病毒在鳥類宿主中有效復制及傳播,而HA穩定性較高的IAV則更加適應于哺乳動物體內[77-80]。值得注意的是,HA蛋白的某些氨基酸突變可通過改變HA蛋白活化的pH閾值來影響病毒的穩定性,進而影響病毒的復制及致病力。例如,有研究發現,H7亞型HA2中E64K突變可通過提高HA活化的pH閾值來降低病毒的穩定性及其在小鼠體內的復制性能[25]。另外,H5亞型HA蛋白上的K387I突變(H3編號),在降低pH閾值的同時也增加了病毒在小鼠中的復制能力及毒力[81]。
3.4 HA蛋白的抗原性變異
HA蛋白是IAV引起中和抗體反應的主要抗原,由于病毒RNA聚合酶缺乏校對功能,在選擇壓力的作用下,極可能出現基因重組及突變,從而重配出新型病毒(如H7N9、H7N2)或已有病毒的不同致病型(如LP H7N9、HP H7N9),并可能造成持續性的流感疫情[2,5,14]。而歷史上引起大流行的流感病毒(如1918年的H1N1、1957年的H2N2),它們在實現人與人之間有效傳播的同時,還能通過更新其抗原性來不斷適應人群,給人類健康造成了極大威脅。
H7N9病毒在從LP到HP的演變過程中其抗原性呈現明顯變異,但關鍵位點尚未明確,故Chang等[20-21]基于1株早期LP病毒A/Anhui/1/2013(H7N9),通過在含有對應抗血清的雞胚中進行連續傳代的方式來模擬病毒在自然界中抗體壓力下的抗原演變,最終鑒定出HA蛋白上A143T、A169T和L235Q 這3個決定性的氨基酸突變,并被證實除了可以改變抗原性也對病毒穩定性和受體結合特性產生了一定影響。值得注意的是,A143T和A169T的突變還同時導致141與167 位分別增加為NLG位點,且該變異均以較高比例出現于近兩年來的HP H7N9自然分離株中[82]。
另外,不同時期的H7N9病毒間也存在抗原性差異。自2013年出現以來,H7N9病毒相繼經歷了從LP→LP與HP共存→HP的演變過程,盡管伴隨著H7-Re1、H7-Re2等禽流感疫苗的有效使用以及相關干預措施的有效執行,禽群和人群中H7N9的分離率均急劇下降,但HP H7N9病毒仍能在部分地區被檢測到。中國動物衛生與流行病學中心、華南農業大學的相關調查分析均指出,禽群中2019年HP H7N9病毒的HA基因與早期HP分離株不僅在遺傳進化上有明顯分歧,疫苗免疫保護效力試驗也表明H7-Re2、rGD76等疫苗株已不能對流行株提供完全保護[9,16]。甘肅疾控中心還發現,2019年,人源HP H7N9病毒與疫苗株SF003相比,其HA1結構域上具有多達15個位點的氨基酸突變,其中,R65K、G132R、V143T、S152P等多個潛在抗原位點已出現明顯變異[10,38]。因此,持續關注HA的抗原性變異是評估疫苗有效性的重要前提。目前,中國農業農村部官方公布的H7N9疫苗株已更新為H7-Re3、rLN79。此外,由于HA蛋白抗原區和受體結合區在空間結構上存在部分重疊,基于血凝抑制(hemagglutinin inhibition, HI)試驗進行的抗原差異性評估也可能會受到受體親和力的干擾。分別有研究指出,H7N9病毒HA的L235Q突變、H1N1病毒HA的E158K突變、H3N2病毒的N145K突變均是因其結合受體的能力發生改變而間接影響了與抗血清的HI滴度,但卻均曾被誤認為是位點突變直接導致了病毒的抗原性變異[62,83-84]。因此,在進行病毒抗原性分析時,也應同時考慮對受體親和力的影響。
鑒于HA蛋白的上述重要功能,眾多學者對其開展了相關研究并鑒定出了一系列關鍵氨基酸位點,被證明不僅會影響HA蛋白的結構與功能,進而也會改變病毒的部分生物學特性。基于此,本文在重點關注H7N9亞型的同時也兼顧其他部分亞型,總結了近年來HA蛋白上涉及IAV穩定性、抗原性、復制性能、受體結合特性、傳播性、致病性等關鍵氨基酸位點(表1),以期為更深入地解析HA基因變異所導致IAV特性改變的相關機制提供參考和借鑒。
4 展 望
目前,國內LP H7N9流感病毒已基本消失,但HP H7N9病毒仍在國內部分地區持續存在,并且有研究表明HP H7N9毒株在實驗動物體內感染過程中出現毒力和傳播能力增強的現象,對家禽及人類健康的威脅依然嚴峻[1-2,8,18]。HA蛋白是H7N9流感病毒表面最主要的糖蛋白和抗原組分,其基因變異所致蛋白結構、功能的變化可能會進而對病毒特性產生重要影響,包括可能產生具有抗原逃逸的變異型子代病毒以維持其在自然界的感染與擴散,或可能獲得更強的結合人源受體的能力以促進其在哺乳動物體內的適應與傳播等。迄今為止,H7N9病毒的持續變異已使疫苗歷經H7-Re1→H7-Re2→H7-Re3、rGD76→rLN79的不斷更新,而疫苗研發相對滯后于病毒變異的現狀迫切要求進一步開發成熟的通用型疫苗。其中, HA莖部由于其結構的相對保守性以及具有激發廣泛保護性免疫的潛能,成為了通用疫苗設計的理想靶標之一。因此,持續監測HA基因的變異、深入解析HA蛋白的功能,必將為進一步揭示H7N9病毒的遺傳演化規律、探究其感染與致病的相關分子機制及疫情的防控奠定重要基礎。