利用單堿基編輯系統(tǒng)定點(diǎn)編輯哈薩克羊MSTN基因的研究
2021-08-23 07:00:56姚旭東蒙亞琦任秀美奧郭延華張譯元王立民
畜牧獸醫(yī)學(xué)報(bào) 2021年8期
姚旭東,蒙亞琦,任秀美奧,郭延華,唐 紅,張譯元,王立民*,周 平*
(1.石河子大學(xué)動物科技學(xué)院,石河子 832000;2. 新疆農(nóng)墾科學(xué)院 省部共建綿羊遺傳改良與健康養(yǎng)殖國家重點(diǎn)實(shí)驗(yàn)室,石河子 832000)
哈薩克羊是新疆最古老的地方綿羊品種之一,具有耐粗飼、適合放牧、肥育性能好等特點(diǎn),很好地適應(yīng)了當(dāng)?shù)靥厥獾淖匀画h(huán)境[1-2]。近年來,隨著圈養(yǎng)規(guī)模的增加以及人們對肉品質(zhì)要求的提高,增加產(chǎn)肉量,提高瘦肉率成為哈薩克羊的育種目標(biāo)。肌肉生長抑制素(myostatin, MSTN)又稱生長分化因子-8(growth differentiation factor-8, GDF-8),是轉(zhuǎn)化生長因子-β(transforming growth factor-β, TGF-β)超家族的一員,參與抑制肌肉分化和生長[3-4]。MSTN基因突變導(dǎo)致其蛋白功能喪失,從而增加肌肉細(xì)胞的數(shù)量以及增大肌肉纖維的直徑,使肌肉過度發(fā)育[5]。在家畜中,MSTN基因存在著一些有益的自然突變,使其表現(xiàn)出雙肌性狀,如雙肌肉牛品種比利時(shí)藍(lán)牛的MSTN基因第三外顯子有11個堿基(937~947 bp)缺失,使MSTN蛋白翻譯提前終止;皮埃蒙特牛第3外顯子發(fā)生g.938G>A替換,這種突變破壞了TGF-β超家族的典型半胱氨酸結(jié)構(gòu),并導(dǎo)致MSTN蛋白失活[6];雙肌肉羊品種特塞爾綿羊(Texel)MSTN基因3′UTR區(qū)存在g.6723G>A突變,為骨骼肌中高度表達(dá)的microRNAs(miR1和 miR206)創(chuàng)建了靶點(diǎn),引起MSTN翻譯抑制[7]。人為敲除MSTN基因會導(dǎo)致綿羊[8-9]、牛[10]和山羊[11]等動物的肌肉增加、體脂率降低。因此,運(yùn)用基因編輯技術(shù)敲除MSTN基因培育雙肌品種成為畜牧業(yè)育種的一個新目標(biāo)。
目前,應(yīng)用最普遍的基因編輯技術(shù)是CRISPR/Cas9(clustered regularly interspersed short palindromic repeats(CRISPR)-associated protein 9)技術(shù),在引導(dǎo)RNA(small guide RNA, sgRNA)的作用下,Cas9酶可以在靶位點(diǎn)造成DNA雙鏈斷裂(double-strand break, DSB),然后利用細(xì)胞自身的2種修復(fù)機(jī)制——非同源末端連接(nonhomologous end joining, NHEJ)和同源性定向修復(fù)(homology directed repair, HDR)機(jī)制實(shí)現(xiàn)基因編輯。其中,NHEJ修復(fù)機(jī)制會引入堿基對插入和缺失(indels)[12];而HDR修復(fù)機(jī)制結(jié)合外源模板可以實(shí)現(xiàn)基因組精確編輯,但HDR機(jī)制較為復(fù)雜,且對細(xì)胞周期(在G2/M中更具活性)有較強(qiáng)的依賴性,其效率通常很低[13]。因此,在特定位點(diǎn)引入精確的遺傳變異仍然具有挑戰(zhàn)性。
堿基編輯器(base editors, BEs)是近年來發(fā)展迅速的基因編輯工具之一,其中的胞嘧啶堿基編輯器(cytidine base editor, CBE)主要由胞苷脫氨酶(cytidine deaminase)、尿嘧啶DNA糖基化酶抑制劑(uracil DNA glycosylase inhibitor, UGI)、Cas9蛋白和sgRNA組成,可以在無需雙鏈斷裂的情況下靶向誘導(dǎo)胞嘧啶(C)向胸腺嘧啶(T)或鳥嘌呤(G)向腺嘌呤(A)的精準(zhǔn)編輯[14]。腺嘌呤堿基編輯器(adenine base editor, ABE)是在CBE的基礎(chǔ)上開發(fā)的,其結(jié)構(gòu)和堿基編輯機(jī)制均類似于CBE,只是腺苷脫氨酶(adenine deaminase)取代了胞嘧啶脫氨酶,可以靶向誘導(dǎo)胞嘧啶A向G或T向C的轉(zhuǎn)變[15]。CBE可以通過將4個密碼子(CAA、CAG、CGA或TGG)精確地轉(zhuǎn)換成終止密碼子阻止基因翻譯,這為基因敲除提供了一種新方法[16],并且不會產(chǎn)生因雙鏈DNA斷裂導(dǎo)致的過度DNA損傷和細(xì)胞死亡[17]。另外,與CRISPR/Cas9依賴HDR實(shí)現(xiàn)精確編輯基因組序列相比,BEs不需要外源供體模板并且不依賴于細(xì)胞周期,因此,可以廣泛用于不同的細(xì)胞類型。近年來,BEs被不斷優(yōu)化改進(jìn),并且已經(jīng)產(chǎn)生了具有更高效率、更窄編輯框和更寬泛PAM識別能力的眾多變體,使其應(yīng)用范圍更廣[18-19]。迄今為止,BEs已在多種動物(如小鼠、兔子、豬、綿羊和猴子等)中獲得成功[20-24]。
經(jīng)檢測,哈薩克羊自然情況下不存在MSTN有益突變,因此,可以利用單堿基編輯技術(shù)編輯該品種MSTN基因,定向引入突變,以期提高產(chǎn)肉量。本試驗(yàn)利用CBE系統(tǒng)在MSTN基因編碼區(qū)提前引入終止密碼子,建立敲除MSTN基因的單堿基編輯體系,為培育MSTN基因編輯哈薩克羊奠定基礎(chǔ),為生產(chǎn)產(chǎn)肉率高、體脂率低的哈薩克羊新品種提供新材料。
1 材料與方法
1.1 材料與試劑
pCMV-AncBE4 max-P2A-GFP質(zhì)粒購自上海吉熒生物技術(shù)有限公司;pGL3-U6-sgRNA-PGK-puromycin為省部共建綿羊遺傳改良與健康養(yǎng)殖國家重點(diǎn)實(shí)驗(yàn)室提供。100 bp DNA Ladder和高保真酶(Pyrobest DNA Polymerase)均購自寶生物工程(大連)有限公司;制性內(nèi)切酶BsaI和T4 DNA Ligase購自美國NEB公司;CruiserTM酶檢測試劑盒(Cat.No.GP0105)購自江蘇吉銳公司;DMEM BASIC培養(yǎng)基購自賽默飛世兒科技(中國)有限公司;胎牛血清購自Biologcal Industries公司;細(xì)胞裂解液(QuickExtractTMDNA Extraction Solutio, Cat.No.QE09050)購自美國Epicentre公司;T載體購自美國Promega公司;嘌呤霉素購自美國Sigma公司;無內(nèi)毒素質(zhì)粒小抽提取試劑盒II型(D6950-01)及Gel & PCR Clean Up Kit購自美國OMEGA公司。葡萄糖、MgCl2·6H2O、NaHCO3、KH2PO4均購自日本和光株式會社。
1.2 試驗(yàn)方法
1.2.1 哈薩克羊胎兒成纖維細(xì)胞的原代培養(yǎng) 通過手術(shù)法取出妊娠30 d的哈薩克羊胎兒,放置于含有200 IU·mL-1青霉素與鏈霉素的生理鹽水中帶回實(shí)驗(yàn)室,取出胎兒用1%PBS清洗3次,然后用75%的無水乙醇消毒30 s,再用1%PBS清洗3次,放入培養(yǎng)皿中,剝?nèi)√荷眢w的皮膚剪碎,均勻鋪在10 cm培養(yǎng)皿中,在培養(yǎng)箱放置3 h后加入8 mL完全培養(yǎng)基(DMEM+15% FBS+1%雙抗),放入5% CO2,37 ℃的培養(yǎng)箱中培養(yǎng),每2 d換一次液,細(xì)胞暈形成后除去組織塊(記為d1代)。
1.2.2 sgRNA及檢測引物設(shè)計(jì) 針對綿羊MSTN基因外顯子序列(序列號:DQ530260.1)設(shè)計(jì)sgRNA(圖1)。pCMV-AncBE4 max-P2A-GFP質(zhì)粒識別間隔相鄰基序(proto-spacer adjacent motifs, PAM)為NGG,sgRNA長度設(shè)為20 bp,為便于連接到載體中,在正義鏈5′端添加ACCG,反義鏈5′端添加AAAC,共設(shè)計(jì)2條sgRNAs(表1)。用Primer Premier 5.0設(shè)計(jì)靶位點(diǎn)檢測引物,STOPsg-1位點(diǎn)檢測引物為Exon-2F、Exon-2R;STOPsg-2位點(diǎn)檢測引物為Exon-3F、Exon-3R(表1)。
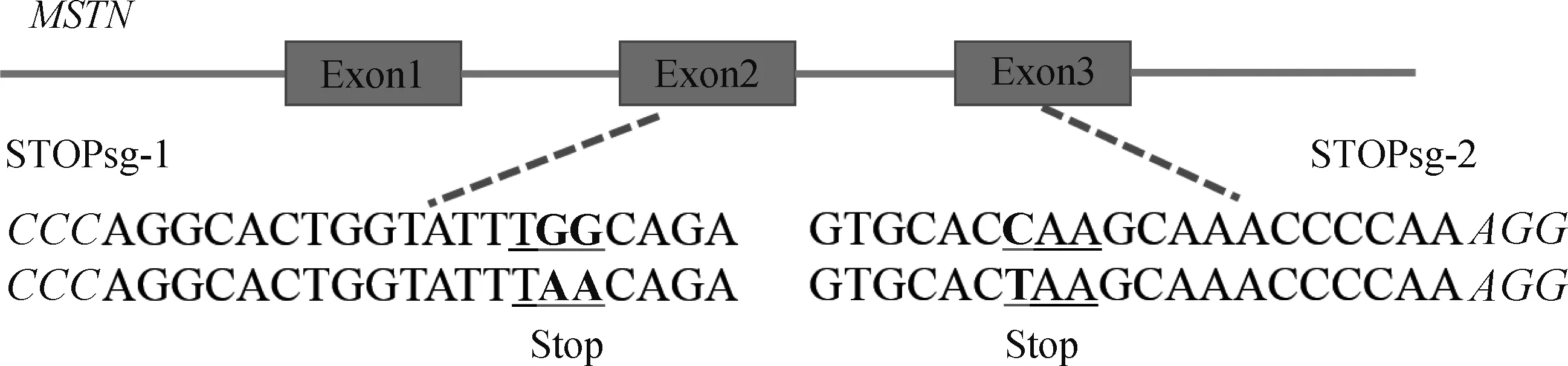
PAM區(qū)域標(biāo)為斜體,靶位點(diǎn)標(biāo)為加粗黑體,突變氨基酸帶下劃線Protospacer adjacent motif (PAM) region in italic, target sites in bold, and mutant amino acids in underscore圖1 哈薩克羊MSTN基因靶位點(diǎn)的示意圖Fig.1 Schematic diagram of the target site of MSTN gene in the Kazakh sheep
1.2.3 MSTN-sgRNA表達(dá)載體構(gòu)建 合成的sgRNA引物分別稀釋成100 μm·L-1后形成雙鏈,反應(yīng)體系如下:正義鏈和反義鏈各1 μL,10×T4 Ligation Buffer 1 μL,T4 PNK 0.5 μL,ddH2O 6.5 μL。反應(yīng)條件:95 ℃ 5 min,95~85 ℃(-2 ℃·s-1),85~25 ℃ (-0.1 ℃·s-1),4 ℃保存。pGL3-U6-sgRNA-PGK-puromycin質(zhì)粒經(jīng)內(nèi)切酶BsaI線性化后膠回收純化。然后將退火后的sgRNA連接至pGL3-U6-sgRNA-PGK-puromycin質(zhì)粒,反應(yīng)體系:雙鏈sgRNA 2 μL,線性化pGL3-U6-sgRNA-PGK-puromycin質(zhì)粒50 ng,10×T4連接酶buffer 1 μL,T4連接酶1 μL,補(bǔ)加ddH2O至10 μL,置于恒溫干浴鍋中16 ℃過夜連接。連接好的MSTN-sgRNA表達(dá)載體轉(zhuǎn)化后用PCR進(jìn)行鑒定,上游引物用A-F(表1),下游引物用sgRNA互補(bǔ)鏈,選取3個 PCR鑒定陽性的質(zhì)粒送測序進(jìn)一步鑒定。
1.2.4 電轉(zhuǎn)液的配制 Solution 1: MgCl2·6H2O 1.2 g,ATP-Na22 g,溶于10 mL ddH2O中,0.22 μm濾器過濾除菌,分裝后-20 ℃保存。 Solution 2:葡萄糖0.02 g,NaHCO30.06 g,KH2PO40.6 g,溶于50 mL ddH2O中,用NaOH將pH調(diào)到7.4,0.22 μm濾器過濾,分裝后4 ℃保存。使用時(shí)按Solution 1∶Solution 2為1∶50的體積比配制成電轉(zhuǎn)液。
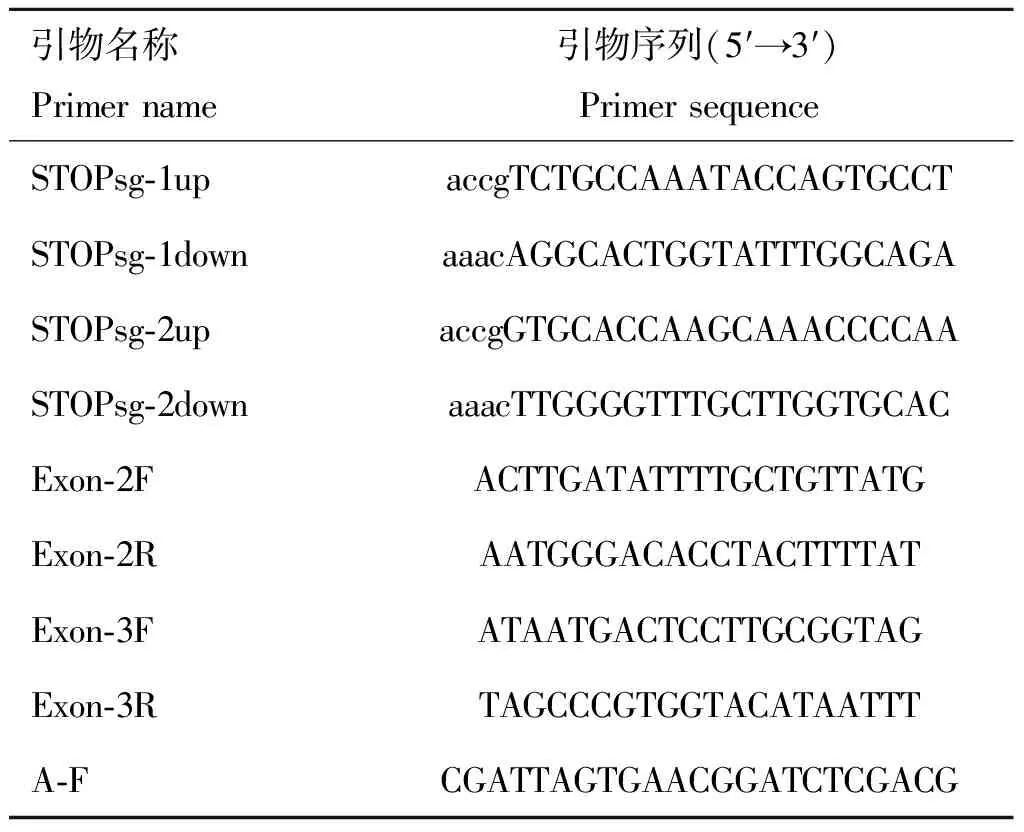
表1 本研究所用引物
1.2.5 電轉(zhuǎn)染哈薩克羊胎兒成纖維細(xì)胞 哈薩克羊胎兒成纖維細(xì)胞傳代后培養(yǎng)于6 cm培養(yǎng)皿中,當(dāng)細(xì)胞融合度為80%~90%時(shí)消化細(xì)胞,離心5 min 后用PBS清洗細(xì)胞,再離心5 min,盡量吸干凈液體后用100 μL電轉(zhuǎn)液懸浮細(xì)胞,每組細(xì)胞的細(xì)胞量約為1×106個,將 6 μg pCMV-AncBE4 max-P2A-GFP質(zhì)粒和2 μg 連接好的pGL3-U6-sgRNA-PGK-puromycin質(zhì)粒共轉(zhuǎn)染哈薩克羊胎兒成纖維細(xì)胞,電轉(zhuǎn)完成后將電轉(zhuǎn)杯放入培養(yǎng)箱中靜置10 min, 接種至6 cm培養(yǎng)皿,37 ℃、5% CO2飽和濕度條件下培養(yǎng)6 h后更換新鮮完全培養(yǎng)基。繼續(xù)培養(yǎng)24 h后觀察熒光表達(dá)效果,運(yùn)用Image J計(jì)數(shù)視野范圍內(nèi)的細(xì)胞數(shù),計(jì)算轉(zhuǎn)染效率,同時(shí)加入嘌呤霉素進(jìn)行藥篩。
1.2.6 CruiserTM酶酶切檢測及測序分析 藥篩72 h后運(yùn)用數(shù)控顯微操作系統(tǒng)(Eppendorf Transfer-Man)收集200個左右?guī)ЬG色熒光的陽性細(xì)胞,將收集到的細(xì)胞用20 μL細(xì)胞裂解液裂解后直接作為PCR模板,使用檢測引物(表1)擴(kuò)增目標(biāo)DNA序列,PCR產(chǎn)物純化后進(jìn)行CruiserTM酶檢測。反應(yīng)體系如下:PCR純化產(chǎn)物取500 ng,補(bǔ)加ddH2O至7 μL,98 ℃變性3 min后關(guān)掉干浴鍋,待溫度下降到40 ℃后取出加2 μL Curiser Buffer和1 μL CruiserTM酶,45 ℃孵育20 min后加入2 μL Stop Buffer,用2%的瓊脂糖凝膠電泳分離檢測,并將PCR純化產(chǎn)物送測序檢測。同時(shí)將PCR純化產(chǎn)物TA克隆后進(jìn)一步測序鑒定,反應(yīng)體系如下:2× Rapid Ligation Buffer 5 μL,pGEM?-T Easy Vector 1 μL,PCR純化產(chǎn)物2 μL,T4 DNA Ligase 1 μL,ddH2O 1 μL。放置于干浴鍋中16 ℃恒溫過夜連接,連接完成后轉(zhuǎn)化送測序。
2 結(jié) 果
2.1 哈薩克羊胎兒成纖維細(xì)胞的培養(yǎng)
采用組織塊貼壁法建立哈薩克羊胎兒成纖維細(xì)胞系。剪碎的胎兒皮膚組織在培養(yǎng)2 d后即可以明顯的觀察到細(xì)胞生長,培養(yǎng)4 d后在顯微鏡下可以觀察到細(xì)胞生長暈,細(xì)胞以梭形為主,形態(tài)不均一,生長力旺盛,無污染,此時(shí)取出組織塊(圖2A)。培養(yǎng)7 d后傳代,由于成纖維細(xì)胞貼壁較快,因此在傳代2 h后換液,以純化胎兒成纖維細(xì)胞。繼續(xù)培養(yǎng)3 d 后再次傳代,細(xì)胞形態(tài)均一,呈梭形,邊緣清晰,生長力旺盛(圖2B),證明成纖維細(xì)胞已純化,成功獲得哈薩克羊胎兒成纖維細(xì)胞。
2.2 構(gòu)建MSTN-sgRNA載體并轉(zhuǎn)染哈薩克羊胎兒成纖維細(xì)胞
測序結(jié)果顯示,MSTN-U6-STOPsg-1和MSTN-U6-STOPsg-2質(zhì)粒構(gòu)建成功(圖3)。
采用電轉(zhuǎn)染將pCMV-AncBE4max-P2A-GFP質(zhì)粒和MSTN-sgRNA質(zhì)粒共轉(zhuǎn)染哈薩克羊胎兒成纖維細(xì)胞,24 h后用熒光顯微鏡觀察綠色熒光表達(dá)情況(圖4)。使用Image J計(jì)算得出STOPsg-1和STOPsg-2的轉(zhuǎn)染效率分別為45.3%、44.7%。
2.3 MSTN基因靶位點(diǎn)突變檢測
CruiserTM酶能對不完全配對的雙鏈DNA切割造成雙鏈DNA斷裂,其識別范圍最小為1個堿基,最大可達(dá)上百個堿基的不配對序列。靶位點(diǎn)的PCR純化產(chǎn)物經(jīng)CruiserTM酶酶切,如果靶位點(diǎn)有突變,則STOPsg-1酶切后出現(xiàn)402和306 bp的片段,STOPsg-2會出現(xiàn)305和231 bp的片段。酶切結(jié)果顯示,STOPsg-1和STOPsg-2的PCR產(chǎn)物均被切開(圖5),說明STOPsg-1和STOPsg-2靶位點(diǎn)可能存在突變,需進(jìn)一步測序驗(yàn)證。
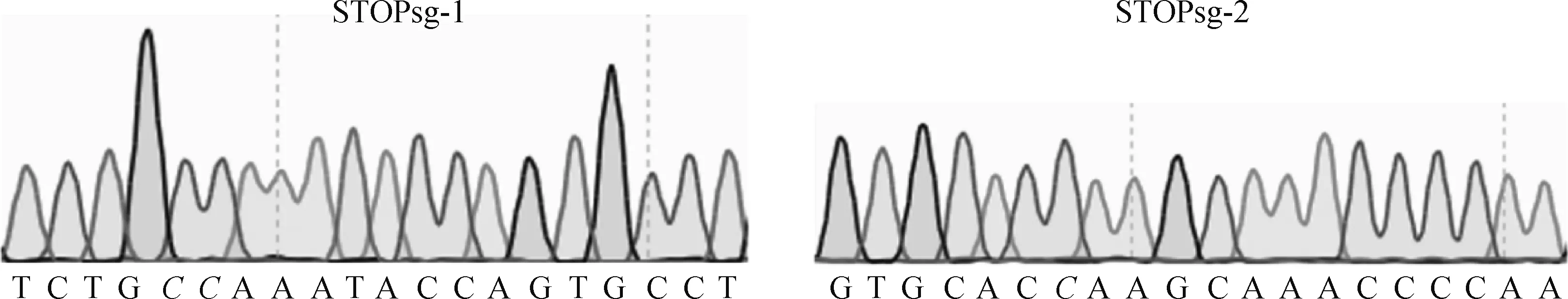
靶位點(diǎn)標(biāo)為斜體The sgRNA target site is labeled in italic圖3 sgRNA質(zhì)粒測序分析Fig.3 Sequencing analysis of sgRNA plasmid

圖4 電轉(zhuǎn)染24 h的熒光表達(dá)情況Fig.4 Green fluorescence expression at 24 h after electrotransfection
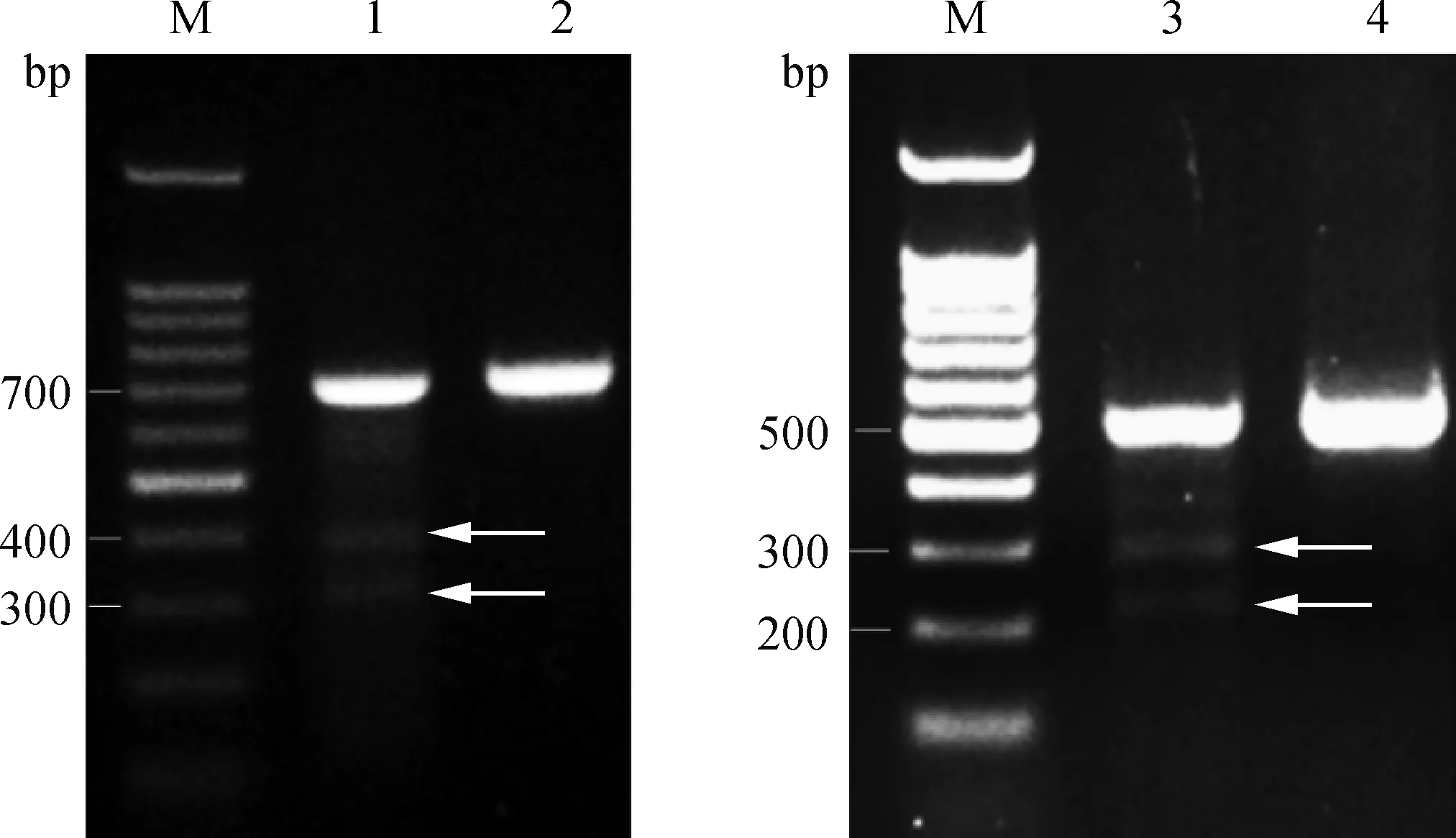
M. DNA相對分子質(zhì)量標(biāo)準(zhǔn); 1.STOPsg-1; 2. STOPsg-1陰性對照; 3.STOPsg-2; 4.STOPsg-2陰性對照; 箭頭表示CruiserTM鑒定的條帶M. 100 bp DNA ladder; 1.STOPsg-1; 2. STOPsg-1 negative control; 3.STOPsg-2; 4.STOPsg-2 negative control; The arrows indicate the bands identified by CruiserTM圖5 CruiserTM酶鑒定結(jié)果Fig.5 Mutation detection by CruiserTM
2.4 MSTN基因靶位點(diǎn)測序結(jié)果分析
AncBE4 max可以使活性窗口內(nèi)的C轉(zhuǎn)換為T,活性窗口為sgRNA PAM位點(diǎn)遠(yuǎn)端數(shù)4~8位。STOPsg-1和STOPsg-2的PCR純化產(chǎn)物送測序檢測,結(jié)果顯示,STOPsg-1和 STOPsg-2靶位點(diǎn)均有重疊峰(圖6),表明STOPsg-1和STOPsg-2靶位點(diǎn)存在突變。然后對STOPsg-1和STOPsg-2的PCR純化產(chǎn)物TA克隆后進(jìn)一步測序驗(yàn)證,結(jié)果顯示,STOPsg-1和STOPsg-2靶位點(diǎn)均發(fā)生C>T或G>A的突變,證明成功提前引入終止密碼子,效率分別為26.7%(4/15)(圖7A)和6.7%(1/15)(圖7B)。同時(shí),在靶位點(diǎn)附近觀察到非目的突變(表2),未觀察到C到A/G的非預(yù)期的突變和隨機(jī)的indels。
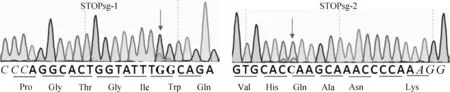
PAM區(qū)域標(biāo)為斜體,靶位點(diǎn)標(biāo)為加粗黑體Protospacer adjacent motif (PAM) region in italic, target sites in bold圖6 靶位點(diǎn)測序峰圖Fig.6 PCR product sequencing map of target sites
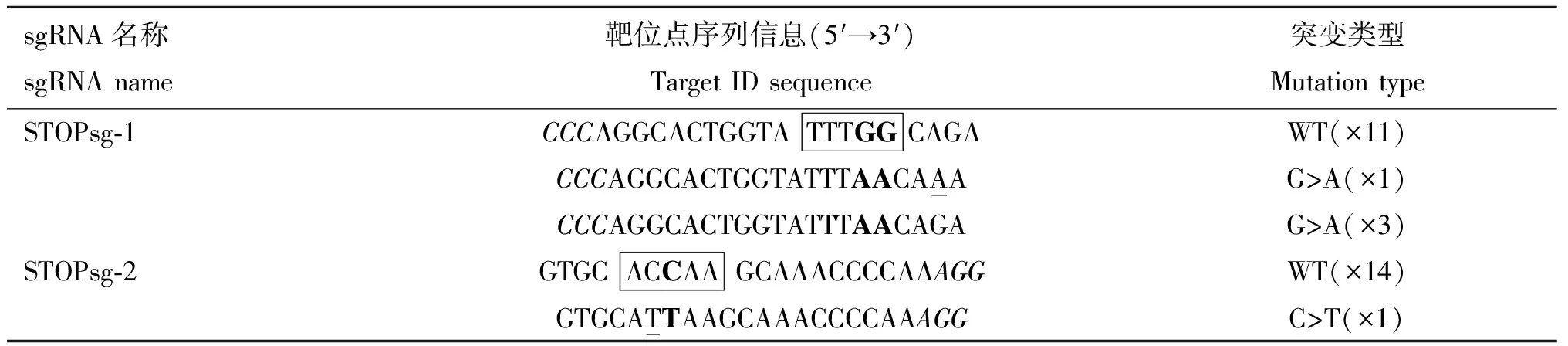
表2 MSTN基因TA克隆測序結(jié)果

A.STOPsg-1; B.STOPsg-2。PAM區(qū)域標(biāo)為斜體,靶位點(diǎn)標(biāo)為加粗黑體,下劃線表示靶點(diǎn)附近非目的突變A.STOPsg-1; B.STOPsg-2. Protospacer adjacent motif (PAM) region in italic, target sites in bold, non-target mutations near the target in underscore圖7 TA克隆Sanger測序峰圖Fig.7 Sanger sequencing peak map of the TA clone
3 討 論
目前,大多數(shù)基因敲除動物是通過CRISPR/Cas9技術(shù)實(shí)現(xiàn)的,這需要形成DSB以使基因產(chǎn)生indels,從而達(dá)到敲除基因的目的。但是,并非所有indels都會導(dǎo)致基因沉默,有些甚至可能導(dǎo)致目標(biāo)蛋白質(zhì)具有新功能[25]。并且當(dāng)CRISPR/Cas9靶向高拷貝數(shù)區(qū)域時(shí),可能會導(dǎo)致過度的DNA損傷并降低適應(yīng)性[26-27]。相比之下,BEs系統(tǒng)不需要引入DSB就能使基因沉默。本研究運(yùn)用CBE(AncBE4 max)系統(tǒng)編輯哈薩克羊胎兒成纖維細(xì)胞MSTN基因,經(jīng)Sanger 測序可知,成功篩選出在MSTN基因第二和第三外顯子能提前引入終止密碼子的2條sgRNAs,未發(fā)現(xiàn)indels。Liu等[28]運(yùn)用BE3在兔MSTN基因提前引入終止密碼子使其失活,獲得的7只兔子中6只是純合子突變,并且對潛在脫靶位點(diǎn)進(jìn)行檢測后未發(fā)現(xiàn)indels和脫靶突變。迄今為止,獲得特定堿基突變動物的主流方法是CRISPR/Cas9系統(tǒng)與供體DNA結(jié)合,通過HDR途徑實(shí)現(xiàn)的,但效果較差,并且會產(chǎn)生許多不需要的indel突變[29-30]。與之相比,BEs系統(tǒng)可以更簡單有效地編輯目標(biāo)堿基,不依賴細(xì)胞周期,不產(chǎn)生DSB或外源DNA模板導(dǎo)入,并且引入隨機(jī)indels很低。Komor等[14]的研究表明,BE3編輯人類細(xì)胞時(shí)突變效率為37%,但indels頻率很低,平均為1.1%,相反,供體DNA介導(dǎo)的HDR僅有0.5%的效率,但indels的百分比更高(4.3%)。
本研究使用AncBE4 max系統(tǒng)在哈薩克羊MSTN基因中提前引入終止密碼子,以敲除MSTN基因,篩選出STOPsg-1和STOPsg-2兩條可以實(shí)現(xiàn)有效編輯的sgRNA,編輯效率分別為26.7%和6.7%,略低于其他文獻(xiàn)的報(bào)道[21,31]。這可能與sgRNA的設(shè)計(jì)有關(guān),本研究設(shè)計(jì)的兩個sgRNA靶核苷酸前面的核苷酸均為C,胞嘧啶核苷脫氨酶 APOBEC1偏好編輯靶核苷酸C的前面是T的序列,編輯效率遵從TC>CC?AC>GC的規(guī)律[14]。本研究還觀察到靶點(diǎn)附近出現(xiàn)非目的突變(表2),但未觀察到C到A/G的非預(yù)期突變和隨機(jī)的indel。出現(xiàn)C到A/G的非預(yù)期突變是因?yàn)镃脫氨基后的U會被細(xì)胞堿基切除修復(fù)(BER)機(jī)制視為錯誤或破壞,然后由無處不在的尿嘧啶DNA糖基化酶(UDG)去除[32],UDG去除U會導(dǎo)致無堿基位點(diǎn)的形成,并隨后通過易錯的聚合酶進(jìn)行修復(fù),從而導(dǎo)致隨機(jī)核苷酸摻入。另外,偶爾的鏈斷裂可能是indel產(chǎn)生的原因[33],迄今為止,很少有研究報(bào)道CBE形成大量插入缺失[34-35]。靶點(diǎn)附近出現(xiàn)非目的突變可能是由于APOBEC1的高活性引起多個Cs脫氨基[36]。盡管當(dāng)使用BEs來破壞啟動子、剪接位點(diǎn)或者過早引入的終止密碼子使基因沉默時(shí),靶點(diǎn)附近非目的突變可能并不是特別重要,但會對探索基因內(nèi)特定點(diǎn)突變產(chǎn)生影響。基于工程化人胞苷脫氨酶APOBEC3A(eA3A)所構(gòu)建的CBE具有編輯特定胞苷的特性,能顯著降低靶點(diǎn)附近的非目的突變[37]。AncBE4 max與工程化APOBEC3A結(jié)合是降低靶點(diǎn)附近非目的突變的有效措施。經(jīng)不斷改進(jìn),BEs在動植物品種改良、動物模型構(gòu)建和疾病治療等方面將展現(xiàn)出巨大的潛力。
4 結(jié) 論
本研究運(yùn)用CBE(AncBE4 max)系統(tǒng)成功編輯了哈薩克羊胎兒成纖維細(xì)胞MSTN基因,篩選出能有效在第二和第三外顯子提前引入終止密碼子的2條sgRNAs,STOPsg-1靶位點(diǎn)編輯效率為26.7%,STOPsg-2靶位點(diǎn)的編輯效率為6.7%。為后期生產(chǎn)產(chǎn)肉率高并具有生物安全性的哈薩克羊奠定基礎(chǔ)。
