3個(gè)欒樹(shù)品種抗寒性比較及生理指標(biāo)變化
2021-08-31 05:54:56劉金秋曹幫華任永俊王世棟段正洪王桂苓孫豐勝
山東林業(yè)科技 2021年4期
劉金秋,曹幫華,任永俊,王世棟,段正洪,高 莉,王桂苓,孫豐勝
(1.山東省地質(zhì)科學(xué)研究院,山東濟(jì)南250013;2.山東農(nóng)業(yè)大學(xué) 黃河下游森林培育國(guó)家林業(yè)和草原局重點(diǎn)實(shí)驗(yàn)室,山東泰安271001;3.淄博市沂源縣歷山街道辦事處,山東淄博256199;4.五蓮縣洪凝街道辦事處,山東日照262300;5.滕州市國(guó)有西崗苗圃,山東滕州277519;6.山東省國(guó)土空間規(guī)劃院,山東濟(jì)南250014;7.東平縣林種苗圃管理服務(wù)中心,山東東平271509)
欒樹(shù)又名燈籠樹(shù),原產(chǎn)于我國(guó)中、北部地區(qū),主要在江蘇、浙江,安徽、江西、河南大量種植,近年來(lái)在山東省城市綠化中廣泛應(yīng)用。欒樹(shù)為無(wú)患子科欒樹(shù)屬落葉喬木或灌木,為深根性植物,生長(zhǎng)速度中等,樹(shù)形優(yōu)美,枝葉茂盛,是庭園遮蔭樹(shù)的良好樹(shù)種; 春季葉子幼嫩多呈紅色,夏季開(kāi)滿黃花,到秋天葉色開(kāi)始變黃,長(zhǎng)出似燈籠的紫紅色果實(shí),具有極高的觀賞價(jià)值;常作為優(yōu)良行道樹(shù)及園林景觀品種[1]。我國(guó)冬季低溫區(qū)域廣泛且差異較大,北欒在北方具有廣泛適應(yīng)性,南欒在北方由于低溫原因,限制了其推廣應(yīng)用,開(kāi)展南欒抗寒良種選育及抗寒性研究對(duì)樹(shù)木引種栽培及選育具有重要意義。
目前欒樹(shù)抗逆性研究方面涉及抗旱性、抗寒性、抗滯塵、抗SO2等[1-5],樹(shù)種間比較較多,欒樹(shù)品種間比較較少。王珊珊[1]、馮獻(xiàn)賓[2]、王巖[6]等對(duì)欒樹(shù)與其他樹(shù)種抗寒性進(jìn)行了比較研究,但關(guān)于欒樹(shù)品種水平上的抗寒性研究很少。
低溫是限制植物分布和園林應(yīng)用的重要環(huán)境因子。采用人工冷凍處理測(cè)定植物抗寒生理指標(biāo)的變化來(lái)揭示和研究植物的抗寒性是目前較為常用的研究方法。本文以引種馴化選育的南欒優(yōu)良無(wú)性系 “滕選”4 號(hào)、5 號(hào)為研究對(duì)象,采取低溫脅迫方法,研究了2 個(gè)無(wú)性系抗寒性及其低溫脅迫下相關(guān)生理生化指標(biāo)的變化并與北欒進(jìn)行了比較,以期為欒樹(shù)良種選育及推廣應(yīng)用提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
2020年12月下旬采集位于山東滕州市西崗國(guó)有苗圃欒樹(shù)“滕選”4 號(hào)、5 號(hào)及北欒長(zhǎng)勢(shì)基本一致、無(wú)病蟲(chóng)害、木質(zhì)化良好的1年生枝條,在0℃至4℃儲(chǔ)存待測(cè)。
1.2 研究方法
1.2.1 試驗(yàn)設(shè)計(jì)
低溫處理方法參照曹曉敏[9]等的方法,將采集的供試枝條用去離子水沖洗干凈,用吸水紙吸干,剪成15~20 cm 的枝段,用石蠟封閉剪口,按照品種貼標(biāo)簽,裝入自封袋,放入可控超低溫冰箱。依次設(shè)-5℃、-10℃、-15℃、-20℃、-25℃、-30℃共6 個(gè)處理溫度,以-5℃為對(duì)照。待冰箱降至設(shè)定溫度后保持12 h,取出第一批試材,并繼續(xù)將冰箱降至下一設(shè)定溫度后保持12 h,取出下一批試材,依次進(jìn)行。將取出的試材在4℃環(huán)境下解凍12 h,測(cè)定電導(dǎo)率。在-5 ~-25℃之間測(cè)定超氧化物歧化酶 (SOD)、脯氨酸、可溶性糖、可溶性蛋白、丙二醛(MDA)的變化。每個(gè)指標(biāo)重復(fù)測(cè)定3 次。
1.2.2 指標(biāo)及測(cè)定方法
(1)相對(duì)電導(dǎo)率測(cè)定[3]
將處理后的枝條剪成0.1~0.2 cm 的小段(去除芽)。稱取0.5 g 放入試管中,加入40ml 去離子水在室溫下浸泡12 h 后待測(cè)。測(cè)定浸泡液的電導(dǎo)率S1,再將盛有浸泡液的試管置于沸水浴30 min,冷卻至室溫后測(cè)定浸泡液電導(dǎo)率S2,相對(duì)電導(dǎo)率=(初電導(dǎo)值/終電導(dǎo)值)×100%。
(2)半致死溫度計(jì)算
通過(guò)Logitic 回歸方程y =k / (1+ae-bx),計(jì)算半致死溫度。其中:y 為相對(duì)電導(dǎo)率,x 為處理溫度、k 為細(xì)胞傷害率的飽和容量,a,b 為曲線方程的參數(shù)[11]。根據(jù)王國(guó)霞[12]等的方法,將k 設(shè)為100%,計(jì)算得出a、b 和決定系數(shù)R2的值,處理溫度與相對(duì)電導(dǎo)率曲線的拐點(diǎn)溫度,即為半致死溫度LT50=lna / b。
(3)生理生化指標(biāo)測(cè)定
采用氮藍(lán)四唑(NBT)法測(cè)定超氧化物歧化酶(SOD)、酸性茚三酮法測(cè)定脯氨酸、蒽酮比色法測(cè)定可溶性糖、考馬斯亮藍(lán)法測(cè)定可溶性蛋白、硫代巴比妥酸顯色法測(cè)定丙二醛( MDA)[13]。
1.2.3 數(shù)據(jù)的處理與分析
利用Excel 軟件進(jìn)行數(shù)據(jù)歸整理、作圖,SPSS 20.0 軟件對(duì)試驗(yàn)數(shù)據(jù)采用單因素方差分析,鄧肯(Duncant)方法進(jìn)行多重比較。
2 結(jié)果與分析
2.1 低溫脅迫下相對(duì)電導(dǎo)率的變化及半致死溫度
低溫脅迫下相對(duì)電導(dǎo)率的變化見(jiàn)圖1,低溫脅迫下,3 個(gè)欒樹(shù)品種的相對(duì)電導(dǎo)率都呈不斷上升趨勢(shì)總體呈S 型曲線。。各品種電導(dǎo)率變化顯著,其中北欒增加幅度最小為88.69%(P<0.05),4 號(hào)增加幅度最大為194.08%(P<0.05),5 號(hào)增加幅度居中為89.80%(P<0.05)。

圖1 低溫脅迫下不同欒樹(shù)相對(duì)電導(dǎo)率的變化/ %
根據(jù)3 個(gè)欒樹(shù)品種在低溫脅迫下相對(duì)電導(dǎo)率的變化,通過(guò)Logistic 擬合回歸方程y=k/(1+ae-bx),計(jì)算得出各品種的半致死溫度(LT50)見(jiàn)表1,欒樹(shù)各品種擬合相關(guān)系數(shù)的R2均在0.9 以上,擬合度較高。各品種的低溫半致死溫度表現(xiàn)出差異性,北欒低溫半致死溫度最低為-27.549℃,其次為 “滕選”4 號(hào)-20.895℃,最高為“滕選”5 號(hào)-18.723℃。3個(gè)欒樹(shù)品種抗寒性排序?yàn)楸睓瑁尽半x”4 號(hào)>“滕選”5 號(hào)。

表1 3 個(gè)欒樹(shù)品種的半致死溫度
2.2 低溫脅迫下抗氧化酶SOD 活性的變化
隨著脅迫溫度的降低,欒樹(shù)各品種的SOD 含量如圖2所示,都呈先降低再升高再降低趨勢(shì)。其中,“滕選”4 號(hào)、5 號(hào)在-5~-10℃不斷降低,-10~-20℃后上升,北欒在-5~-15℃不斷降低,-15~-20℃后上升,所有欒樹(shù)SOD 含量在-20℃快速上升后降低。“滕選”4 號(hào)、5 號(hào)、北欒SOD 含量最高比最低分別增加了108.26%(P<0.01)差異極顯著、81.56%(P<0.01)差異極顯著、31.22%(P<0.05)差異顯著。

圖2 低溫脅迫下不同欒樹(shù)品種SOD 活性的變化/(U·g-1)
2.3 低溫脅迫下滲透調(diào)節(jié)物質(zhì)脯氨酸、可溶性糖、可溶性蛋白含量的變化
隨著脅迫溫度的降低,欒樹(shù)各品種的脯氨酸含量如圖3所示,“滕選”4 號(hào)、5 號(hào)呈先降低后升高趨勢(shì),北欒則不斷升高。“滕選”4 號(hào)、5 號(hào)在-10℃前有所下降,-15℃后都持續(xù)升高,-15~-25℃之間增加緩慢,而北欒在-5~-10℃增加緩慢,-10~-15℃之間增加迅速,-15~-25℃之間增加緩慢,可能與-15℃之前北欒所受損傷低于“滕選”4 號(hào)、5 號(hào),脯氨酸含量偏低,隨著傷害加重,脯氨酸含量急劇增高。“滕選”4號(hào)、5 號(hào)分別比對(duì)照增加了136.56%(P <0.05)、129.01%(P<0.05),差異顯著,北欒比對(duì)照增加了320.16%(P<0.01),差異極顯著。

圖3 低溫脅迫下不同欒樹(shù)品種脯氨酸含量的變化/(mg·g-1)
低溫脅迫下,欒樹(shù)各品種的可溶性糖含量變化如圖4所示,隨著低溫脅迫的加劇,“滕選”4 號(hào)、5號(hào)可溶性糖含量均呈先上升后下降趨勢(shì),在-20℃含量高于其他脅迫溫度。北欒可溶性糖含量呈不斷升高趨勢(shì)。其中,在-5~-20℃低溫處理階段內(nèi),所有品種可溶性糖含量都呈上升趨勢(shì),在-20~-25℃“滕選”4 號(hào)、5 號(hào)緩慢下降而北欒迅速增加,“滕選”4號(hào)、5 號(hào)、北欒相比較對(duì)照分別增加了211.18%(P<0.01)、99.37%(P<0.01)、112.21%(P<0.01),均差異極顯著。

圖4 低溫脅迫下不同欒樹(shù)品種可溶性糖含量的變化/(mg·g-1)
低溫脅迫下,欒樹(shù)各品種的可溶性蛋白含量變化如圖5所示,隨著低溫脅迫的加劇,“滕選”5 號(hào)、北欒可溶性蛋白含量均呈現(xiàn)先上升后下降趨勢(shì),在-20℃含量高于其他脅迫溫度。“滕選”4 號(hào)可溶性糖含量呈不斷升高趨勢(shì)但在最低溫-25℃時(shí)含量仍未高于“滕選”5 號(hào)及北欒。在-5~-15℃低溫處理階段內(nèi),所有品種可溶性蛋白含量緩慢增加,-15~-20℃迅速增加,之后“滕選”5 號(hào)、北欒至-20℃快速升高后下降明顯。“滕選”4 號(hào)呈持續(xù)增加趨勢(shì)。增加幅度上,“滕選”4 號(hào)、5 號(hào)、北欒分別比最低對(duì)照增加了1252.39%(P<0.01)、1467.90%(P<0.01)、1086.51%(P<0.01),均差異極顯著。
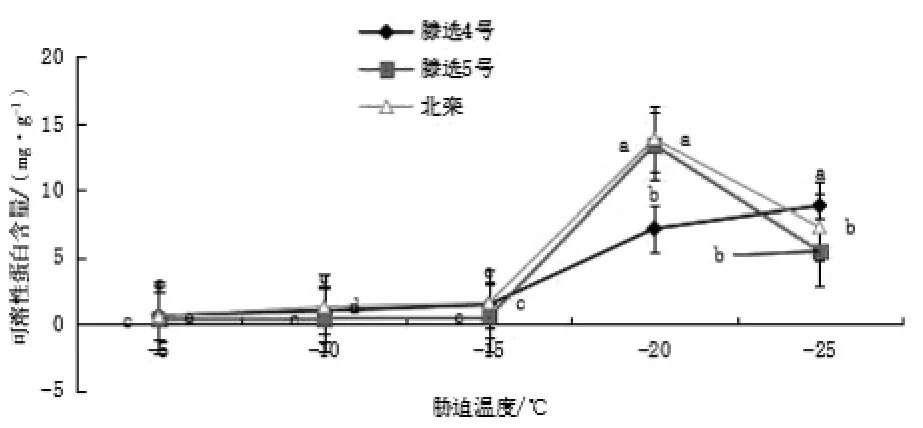
圖5 低溫脅迫下不同欒樹(shù)品種可溶性蛋白含量的變化/(mg·g-1)
2.4 低溫脅迫下MDA 含量的變化
低溫脅迫下,欒樹(shù)各品種的MDA 含量變化如圖6所示,隨著低溫脅迫的加劇,各欒樹(shù)品種MDA含量均呈現(xiàn)先上升后下降趨勢(shì)。其中,“滕選”4 號(hào)、5 號(hào)在-5~-15℃低溫處理階段內(nèi),含量升高,在-15℃之后下降。而北欒在-5~-20℃含量升高,自-20℃之后開(kāi)始下降。各欒樹(shù)MDA 含量開(kāi)始下降溫度都早于各品種的低溫半致死溫度。北欒MDA 含量開(kāi)始下降溫度晚于其他兩品種。各品種MDA 含量最高值比對(duì)照分別增加了45.50%(P<0.01)差異極顯著、28.30%(P<0.05)差異顯著、51.47%(P<0.01)差異極顯著。

圖6 低溫脅迫下不同欒樹(shù)品種MDA 含量的變化/(μmol·g-1)
3 討論與結(jié)論
低溫對(duì)膜的傷害主要表現(xiàn)在膜透性的改變,從而使細(xì)胞內(nèi)物質(zhì)大量向外滲透[14]。柳新紅等研究表明電導(dǎo)率的變化體現(xiàn)出質(zhì)膜受傷害程度以及所測(cè)材料的抗寒性大小,電解質(zhì)滲出率達(dá)50%得出的半致死溫度可以更加準(zhǔn)確反映植物的抗寒性[15]。相對(duì)電導(dǎo)率越小,抗寒性越強(qiáng),反之,抗寒性越弱[16]。隨著溫度降低,各品種相對(duì)電導(dǎo)率不斷升高,“滕選”4號(hào)、5 號(hào)在-20℃已經(jīng)達(dá)到51.91%、52.82%,北欒在-30℃達(dá)到50.25%,細(xì)胞膜受到嚴(yán)重?fù)p害,北欒到達(dá)50%電導(dǎo)率的脅迫溫度更低,表明北欒相對(duì)“滕選”4 號(hào)、5 號(hào)更耐寒。通過(guò)Logistic 擬合回歸方程計(jì)算得出各品種的半致死溫度,3 種欒樹(shù)的低溫半致死溫度由低到高分別為北欒-27.549℃、“滕選”4號(hào)-20.895℃、“滕選”5 號(hào)-18.723℃。3 種欒樹(shù)抗寒性排序?yàn)楸睓瑁尽半x”4 號(hào)>“滕選”5 號(hào)。2 個(gè)新選南欒無(wú)性系抗寒性仍未達(dá)到北欒水平,但從抗寒性來(lái)看,完全可以在山東省范圍內(nèi)栽培應(yīng)用。
SOD 是植物細(xì)胞重要的保護(hù)酶,其活性高低與植物抗寒性強(qiáng)弱有密切關(guān)系[17]。本研究中,各品種SOD 活性均呈先下降再升高再下降趨勢(shì),在-5~-25℃低溫處理階段內(nèi)的峰值在出現(xiàn)在-20℃,但變化趨勢(shì)與王珊珊[1]不斷升高、馮獻(xiàn)賓[2]有所不同。
脯氨酸、可溶性糖、可溶性蛋白是植物體內(nèi)低溫脅迫下重要的滲透調(diào)節(jié)物質(zhì)[14]。低溫脅迫下大多數(shù)植物會(huì)通過(guò)積累滲透調(diào)節(jié)物質(zhì)增加細(xì)胞液的濃度,降低滲透勢(shì),借此提升抗寒能力[18]。本次研究表明,隨著低溫脅迫加劇,脯氨酸含量“滕選”4 號(hào)、5號(hào)先降低后升高,北欒持續(xù)升高,北欒上升幅度最大;可溶性糖含量“滕選”4 號(hào)、5 號(hào)先升高后降低,北欒持續(xù)升高;可溶性蛋白“滕選”5 號(hào)、北欒先升高后降低,“滕選4 號(hào)持續(xù)升高。各品種滲透調(diào)節(jié)物質(zhì)升高明顯,表明了其面對(duì)低溫脅迫的敏感性,低溫處理階段內(nèi)滲透調(diào)節(jié)物質(zhì)升高后降低說(shuō)明低溫脅迫對(duì)植物細(xì)胞造成了不可逆轉(zhuǎn)傷害,生理機(jī)能下降。滲透調(diào)節(jié)物質(zhì)的總體變化趨勢(shì)與之前的研究變化相似[1,19]。
植物在逆境下細(xì)胞產(chǎn)生大量的自由基加劇細(xì)胞膜過(guò)氧化,使細(xì)胞膜破壞加劇。MDA 是脂膜過(guò)氧化的最終產(chǎn)物,是反映了細(xì)胞膜傷害的重要指標(biāo)[20]。本試驗(yàn)MDA 含量呈先升高后降低趨勢(shì),表明低溫超出試材的耐受能力,植物細(xì)胞受到傷害嚴(yán)重難以抵御嚴(yán)寒使生理機(jī)能紊亂MDA 下降。北欒開(kāi)始下降溫度在-20℃低于“滕選”4 號(hào)、5 號(hào)-15℃,表明其脂膜過(guò)氧化導(dǎo)致生理機(jī)能下降的溫度低于 “滕選”4 號(hào)、5 號(hào)。MDA 含量都在低溫半致死溫度前開(kāi)始下降,此結(jié)論和呂小軍等[17]對(duì)花椒抗寒中MDA 的變化一致。
本文研究在室內(nèi)通過(guò)人工降溫實(shí)現(xiàn)的,各品種低溫半致死溫度作為參考,實(shí)際生產(chǎn)應(yīng)用中3 個(gè)樹(shù)種抗寒性可能與試驗(yàn)結(jié)果有一定誤差,可根據(jù)實(shí)際情況綜合判定。
