泥頁巖有機質孔隙差異特征及影響因素分析
——以我國典型海相、陸相、過渡相儲層為例
2021-08-31 08:17:48谷淵濤李曉霞萬泉楊曙光
沉積學報 2021年4期
谷淵濤,李曉霞,萬泉,楊曙光
1.河南工程學院資源與環境學院,鄭州 451191
2.中國科學院地球化學研究所礦床地球化學國家重點實驗室,貴陽 550081
0 引言
近年來隨著非常規油氣資源工業化勘探開發的快速發展,頁巖油氣對于能源領域的支撐作用越發重要[1]。與常規油氣勘探中的生、儲、蓋、圈、運、保等核心要素不同,連續或準連續分布甜點區(段)是頁巖油氣地質學研究的核心,兩者均受到沉積環境的控制[1]。有機質作為頁巖油氣的主要載體,其沉積富集過程與泥頁巖沉積環境密切相關,其中發育的納米孔隙是有機質向烴類轉化的產物,對頁巖油氣資源富集有重要作用[2]。開展不同沉積相泥頁巖中有機質孔隙特征研究可為頁巖儲層甜點區(段)優選提供重要理論支撐,豐富非常規油氣沉積學的理論知識,以助力非常規油氣資源的勘探開發。
目前關于泥頁巖中有機質孔隙特征已有大量研究:1)演化過程極為復雜。部分學者認為有機質孔隙隨熱成熟度(Ro)的增加呈逐漸發育的趨勢[3-5]。這主要基于有機質的熱成熟過程伴隨著干酪根熱解、裂解以及液態烴裂解生氣的進行,從而使有機質孔隙不斷產生和演化[2,6]。也有觀點認為泥頁巖中有機質孔隙隨熱成熟度的增加呈現出階段性特征[7-11]。例如,Curtiset al.[9]通過對Woodford頁巖進行次生有機質孔隙的演化研究,發現有機質孔隙在熱成熟度為0.51%~6.36%的范圍內并無明顯規律,說明僅熱成熟度難以預測孔隙的發展趨勢。Chenet al.[10]通過對低熟泥頁巖的熱模擬實驗研究,發現在不同熱演化階段有機質孔隙發育特征有明顯區別:Ro為0.6%~2.0%時,有機質孔隙度呈先下降后上升的趨勢;Ro為2.0%~3.5%時,有機質孔隙持續發展;Ro>3.5%時,有機質孔隙發生破壞和轉化。2)影響因素的差異性。有機質在熱演化過程中向烴類轉化而形成納米孔隙,但演化過程中因各種因素差異性的影響,引起有機質孔隙的演化模式多種多樣。因此,熱成熟度并非影響有機質孔隙發育的唯一主導因素,其他因素在特定條件下對孔隙特征也有明顯的制約。例如,Millikenet al.[12]認為總有機碳(TOC)含量對于有機質孔隙的控制比熱成熟度更為明顯:當TOC小于5.5%時,TOC與孔隙度呈正相關;當TOC大于5.5%時,TOC的增加對于孔隙度幾乎沒有影響。此外,TOC含量對于微孔、介孔、大孔的影響也有明顯區別,多數研究表明微孔與TOC含量最為密切,其次為介孔[13-16]。有機質顆粒的大小也影響孔隙結構特征,顆粒越大越有利于孔隙的發育和生長,顆粒越小則抑制了孔隙的形成和演化[12]。有機質類型對于孔隙結構的影響也十分明顯:I型和II型干酪根發育有機質孔隙的能力遠遠高于III型干酪根,這取決于各類型有機質的生烴潛力[6,17-18]。類似地,有機質顯微組分因生烴潛力不同其發育有機質孔隙的能力也有差異,如腐泥組、鏡質組往往比惰質組更發育孔隙[6,19-20]。除上述有機地球化學特征外,無機礦物、構造作用等也嚴重影響著有機質孔隙的演化進程。例如,黏土礦物與有機質的相互作用對生烴有一定的催化或抑制作用,從而促進或抑制孔隙的形成及演化[21-22];構造條件通過影響排烴過程而引起有機質孔隙發育特征的差異性[23-25]。
上述研究深入闡明了泥頁巖中有機質孔隙的形成演化機理,并揭示了各因素在孔隙演化過程中的影響機制。然而,關于不同沉積相泥頁巖有機質孔隙發育特征的影響因素仍無定論。本文在梳理前人關于有機質孔隙研究結果的基礎上,以我國不同沉積相泥頁巖儲層(鄂爾多斯盆地延長組(T3y)陸相泥頁巖、四川盆地東南緣龍馬溪組(S1l)和牛蹄塘組(?1n)海相泥頁巖、南華北盆地山西組(P1s)過渡相泥頁巖)為研究對象,系統表征了有機質孔隙發育特征,分析了各組儲層影響有機質孔隙發育特征的主要因素,研究結果對頁巖油氣生儲機制和勘探開發具有重要的理論和實踐意義。
1 地質概況
鄂爾多斯盆地是屬于構造穩定的大型沉積盆地,具有巨大的油氣資源儲量[26-27]。盆地包含六個構造單元:北部的伊盟隆起、南部的渭北隆起、西部的天環凹陷及西緣沖斷帶、東部的晉西撓褶帶以及中部的伊陜斜坡[27-28]。盆地中延長組被證實是最具潛力的烴源巖,并被劃分為10段。其中,在深湖—半深湖沉積環境下形成的長7段的巖性包括油頁巖、黑色頁巖和碳質頁巖,長7段因含有豐富的有機質而成為最重要的烴源巖[29]。本研究所采集的延長組泥頁巖樣品取自伊陜斜坡東部(圖1A),且全部來自長7段泥頁巖。四川盆地是我國最大的含油氣盆地,也是目前頁巖氣開采最成功的區域[30]。在四川盆地內部及周緣,志留系和寒武系富有機質頁巖分布范圍廣、厚度大,并且經歷了多旋回構造體系下的深埋藏和強改造過程,是頁巖氣勘探開發的重要目標層位[31-32]。其中龍馬溪組和牛蹄塘組泥頁巖是最具潛力的頁巖氣儲層。本研究所選取的龍馬溪組和牛蹄塘組樣品均采自四川盆地東南緣的貴州省境內(圖1B)。上古生界地層在華北地區分布十分廣泛,南華北盆地的頁巖氣調查表明該區域海陸過渡相泥頁巖具有一定的生儲烴能力,是潛在的頁巖氣儲層[33]。該盆地最具代表性的烴源巖是二疊系山西組和太原組[34]。本研究所選取的山西組樣品采自太康隆起西部(圖1C),緊鄰中牟凹陷和中條—豫西隆起。
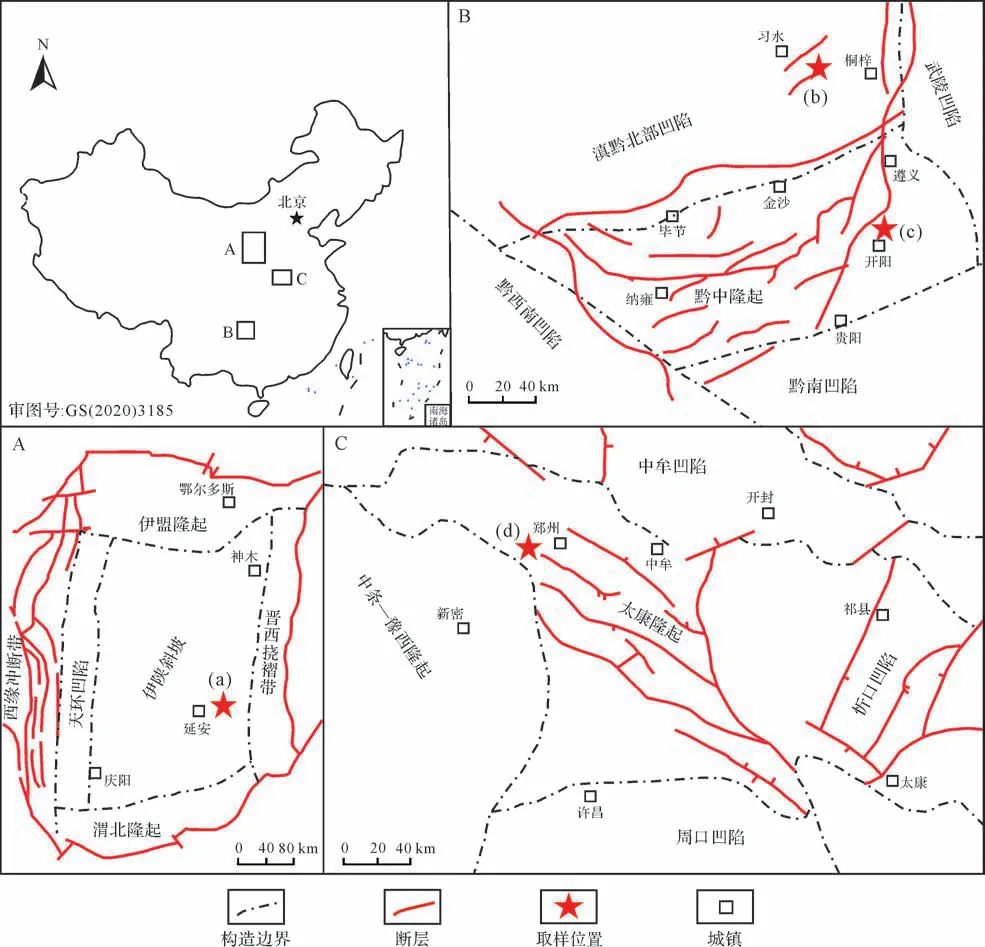
圖1 研究區地質概況及取樣位置Fig.1 Regional geology of the study area and sampling locations
沉積環境決定了泥頁巖的有機質豐度、類型及顯微組成[6],四組泥頁巖樣品因沉積環境差異形成了不同類型的有機質,進而影響泥頁巖的生烴潛能。鄂爾多斯盆地延長組長7段在深湖—半深湖沉積環境下形成II型為主的干酪根,少量干酪根為III型;顯微組分中腐泥組最發育,鏡質組次之,惰質組最不發育[35]。四川盆地龍馬溪組沉積于深水—淺水陸棚環境,有機質母源輸入以各種浮游藻類為主,有機質類型以Ⅰ型為主,顯微組分主要是鏡質組(瀝青),伴有少量腐泥組[6,20,36]。研究區內牛蹄塘組沉積環境與龍馬溪組類似,為深水陸棚向淺水陸棚過渡的沉積相,主要有機質類型為Ⅰ型,顯微組分以腐泥組和瀝青為主[37]。南華北盆地山西組主要形成于三角洲體系的海陸過渡相環境,有機質類型主要為III型,少量為Ⅱ2型,顯微組分以鏡質組、惰質組為主[33,38-39]。
2 樣品及研究方法
2.1 樣品
本研究一共采集了四套地層共53個泥頁巖樣品。其中,13個延長組長7段樣品采集于鄂爾多斯盆地941#采油井(取樣點a),取樣深度范圍694.0~718.0 m(表1)。龍馬溪組樣品取自貴州習水縣騎龍村剖面(取樣點b),為避免風化作用的影響,我們利用紹爾便攜式取樣鉆機采集淺層鉆孔樣品。該剖面將五峰—龍馬溪組分為三段,其中第一段為典型的富有機質泥頁巖,第一段又分為9小層,除第1小層為五峰組外,其余8層均為龍馬溪組。我們依據這8小層的垂直厚度分別取樣,共獲取14個淺層鉆孔巖心樣品(表1)。牛蹄塘組14個泥頁巖樣品取自貴州開陽ZK105鉆孔(取樣點c),取樣深度范圍為656.8~717.8 m(表1)。山西組12個樣品取自河南鄭州ZK02109鉆孔(取樣點d),取樣深度范圍為139.0~222.7 m(表1)。
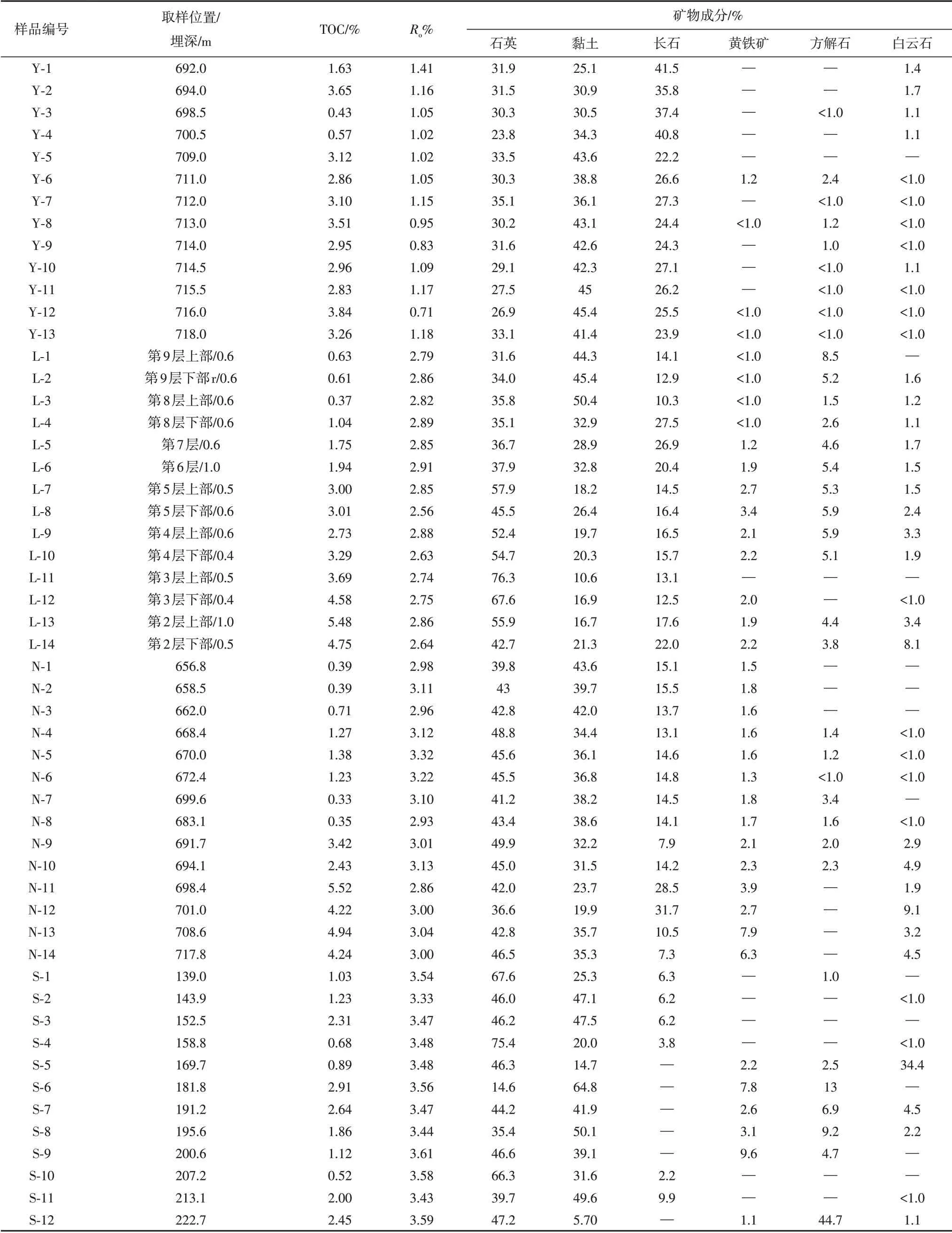
表1 泥頁巖樣品的物質組成特征Table 1 The material composition of the shale samples
2.2 總有機碳(TOC)
總有機碳(TOC)含量是根據GB/T 19145—2003《沉積巖中總有機碳的測定》進行分析測試的。稱取0.5 g左右樣品于燒杯中,緩慢加入過量的鹽酸溶液(HCl∶H2O=1:7(體積比)),以去除樣品中的無機碳;溫度控制在60℃~80℃,溶樣2 h以上至反應完全為止,要盡量保證樣品在反應過程中沒有濺出;反應完全后用去離子水洗滌,并在10 000 r/min的轉速下離心沉淀,如此重復三次以去除氯離子;洗滌干凈后的樣品在60℃下烘干備用;使用河南工程學院分析測試中心的vario MACRO cube有機元素分析儀完成樣品TOC含量測試。
2.3 熱成熟度
根據前人關于熱成熟度的研究,激光拉曼光譜是一種計算反射率的非常合適的方法[40-42]。我們采用河南工程學院分析測試中心的Renishaw Invia Reflex激光拉曼光譜儀來測試樣品的成熟度。計算公式為[43]:R=0.0537d(G-D)-11.21,其中G、D、d分別指的是石墨碳、無序碳的峰位置以及G峰和D峰的間隔。由于在下古生界海相頁巖中缺乏鏡質體,因此利用瀝青反射率來代表樣品的熱成熟度。鏡質體反射率(Ro)和瀝青反射率(Rb)的關系已獲得較為成熟的研究,根據公式Ro=(Rb+0.244 3)/1.049 5進行熱成熟度的換算[44]。
2.4 礦物成分分析
樣品的礦物成分采用河南工程學院分析測試中心Bruker D8 ADVANCE X射線衍射儀進行測試分析。測試之前需將樣品破碎并研磨至200目以下。掃描范圍、步長分別為5°~80°、0.013°。參照石油天然氣行業標準(SY/T)5163—2010進行半定量計算。
2.5 低壓N2吸附
低壓N2吸附實驗采用河南工程學院分析測試中心的Quantachrome autosorb iQ全自動氣體吸附分析儀完成。在吸附測試之前,將樣品在150℃條件下脫氣4 h以去除易揮發物質。樣品的比表面積采用多點BET(Brunauer-Emmett-Teller)方法計算。
2.6 氬離子拋光—掃描電鏡
為了獲得平整表面,在掃描電鏡觀察之前采用Leica EM TIC 3X氬離子拋光儀對樣品進行拋光。同時,對樣品表面進行了噴金處理,使其具有導電性,然后采用中國科學院地球化學研究所月球與行星研究中心的FEI Scios FE-SEM進行表面形貌及成分分析,加速電壓為20 kV。
3 結果與討論
3.1 有機地球化學特征
四組泥頁巖樣品的有機地球化學特征見表1。其中,延長組樣品TOC分布在0.4%~3.9%范圍內,平均含量2.8%;該組樣品Ro范圍為0.7%~1.4%,平均1.1%,表明該組樣品多為富有機質低熟泥頁巖。龍馬溪組泥頁巖TOC分布范圍為0.3%~5.5%,平均含量2.6%;Ro分布在2.5%~2.9%之間,平均2.8%。牛蹄塘組泥頁巖TOC范圍為0.3%~5.5%,平均含量2.2%;Ro范圍為2.8%~3.3%,平均3.1%。龍馬溪組和牛蹄塘組泥頁巖由底至頂均表現出TOC逐漸降低的趨勢,是由沉積環境的變化所引起,即從深水環境向淺水環境過渡[36,45]。山西組泥頁巖TOC范圍為0.5%~2.9%,平均含量1.6%;Ro范圍為3.3%~3.6%,平均3.5%。該組樣品垂向上TOC分布不均,這與海陸過渡相沉積環境有關。雖然該地層晚于龍馬溪組和牛蹄塘組,但區域熱事件造成了更高的熱成熟度(>3.0%)[34,38]。
3.2 礦物組成
根據表1的礦物成分,以石英和長石、碳酸鹽、黏土為三端元對比了這四組樣品的礦物組成特征。如圖2所示,延長組、龍馬溪組和牛蹄塘組樣品的主要礦物成分均為石英、黏土和長石,并含有少量的黃鐵礦和碳酸鹽礦物。相比于牛蹄塘組和延長組樣品,龍馬溪組泥頁巖含有更高的石英和長石含量,代表了更高的脆性,可壓裂性較好。另外,山西組泥頁巖礦物組成差異性較大,這與沉積環境變化頻繁關系密切,體現了海陸過渡相環境對礦物組成的重要影響。

圖2 礦物組成三端元圖Fig.2 Ternary diagram of the mineralogical constituents
3.3 孔隙參數特征
根據低壓N2吸附,分別計算了各個樣品的BET比表面積、孔體積、平均孔徑,如表2所示。其中,延長組樣品的比表面積范圍為8.4~17.2 m2/g,平均11.5 m2/g;孔體積范圍為0.050~0.088 cm3/g,平 均0.069 cm3/g;平均孔徑范圍為16.6~30.8 nm,平均24.4 nm。龍馬溪組泥頁巖的比表面積范圍為21.6~36.5 m2/g,平均26.8 m2/g;孔體積范圍為0.088~0.111 cm3/g,平均0.095 cm3/g;平均孔徑范圍為10.1~18.4 nm,平均14.4 nm。牛蹄塘組泥頁巖的比表面積范圍為15.5~32.0 m2/g,平均22.6 m2/g;孔體積范圍為0.062~0.107 cm3/g,平均0.087 cm3/g;平均孔徑范圍為9.3~22.3 nm,平均16.4 nm。山西組樣品的比表面積范圍為8.0~15.5 m2/g,平均11.8 m2/g;孔體積范圍為0.050~0.079 cm3/g,平均0.065 cm3/g;平均孔徑范圍為19.3~27.5 nm,平均22.3 nm。

表2 泥頁巖樣品的孔隙參數特征Table 2 Pore parameter characteristics of the shale samples
3.4 有機質孔隙發育特征差異
圖3~6顯示不同熱成熟度泥頁巖中的孔隙發育特征差異明顯。延長組樣品基本不發育有機質孔隙,但發育有較多的礦物粒間孔及微裂隙;有機質周圍發育有較多微裂隙,對于烴類運移有重要作用(圖3)。龍馬溪組泥頁巖有機質孔隙極為發育,多為大孔和介孔,且形態多樣,主要以圓形、橢圓形、不規則的狹縫型為主(圖4)。牛蹄塘組泥頁巖中多數有機質顆粒無納米孔隙,且有機質與礦物顆粒之間往往發育因成巖演化所形成的微裂隙[46-47];也有部分有機質發育介孔,多為圓形、橢圓形(圖5)。這種有機質孔隙發育的非均質性與孔隙的排烴作用是否完全密切相關[12]。值得注意的是,龍馬溪組和牛蹄塘組泥頁巖中有機質—黏土復合體中發育有較多的納米孔隙(圖4d,e、圖5d,f),且形態特征與有機質顆粒中的孔隙有一定差異,源于黏土層對孔隙的影響。山西組樣品與延長組類似,有機質孔隙基本不發育(圖6b,c),這與其III型干酪根生烴潛量較低(不易發育納米孔隙)有密切關系[38];但明顯可見部分有機質的塌陷(圖6d~f),可使數個甚至數十個小孔轉化為大孔,因此產生了較多數百納米甚至微米級大孔,說明這部分有機質在演化過程中產生過較多的納米級孔隙,之后由于過高的熱演化程度導致有機質結構發生改變造成孔隙坍塌。
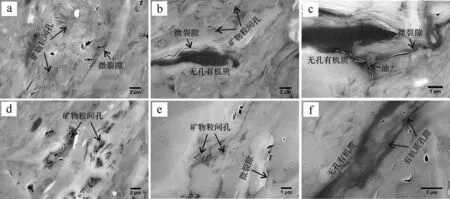
圖3 延長組(T3y)泥頁巖樣品的微結構特征Fig.3 Microstructure characteristics of the T3y shale samples

圖4 龍馬溪組(S1l)泥頁巖樣品的微結構特征Fig.4 Microstructure characteristics of the S1l shale samples
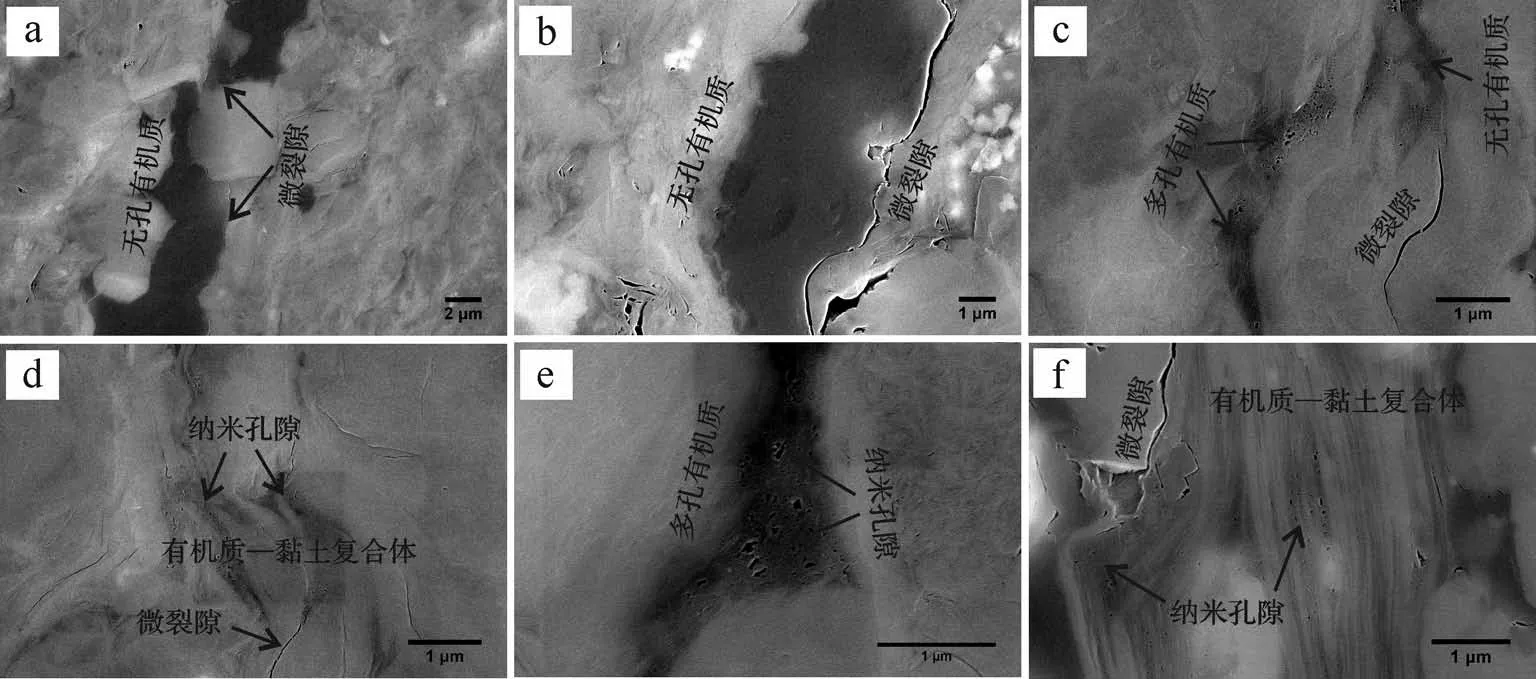
圖5 牛蹄塘組(?1n)泥頁巖樣品的微結構特征Fig.5 Microstructure characteristics of the?1n shale samples
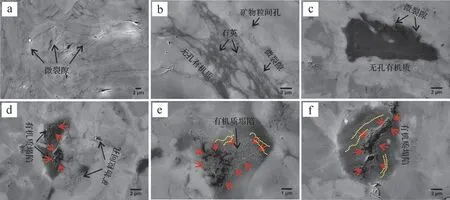
圖6 山西組(P1s)泥頁巖樣品的微結構特征Fig.6 Microstructure characteristics of the P1s shale samples
根據TOC與孔隙參數的相關性分析(圖7),可見有機質對于孔隙參數有重要影響,熱演化程度不同兩者之間的相關性也會發生明顯改變。隨著熱成熟度的增加,比表面積與TOC先呈負相關,后呈明顯的正相關,最后趨于無明顯相關性;同時,平均孔徑與TOC表現出正好相反的相關性變化規律,這表明有機質發生了從無孔到多孔再到消失的轉化;而孔體積總體上與TOC的相關性較弱,這主要是因為孔體積是由與礦物相關的大孔所主導。另外,龍馬溪組和牛蹄塘組樣品的TOC與孔隙參數具有相似的相關性,只是牛蹄塘組的相關系數更大,并且牛蹄塘組樣品的孔體積與TOC呈弱負相關,根據SEM觀察分析有機質孔隙的收縮導致孔徑減小,使得TOC與比表面積的正相關、與平均孔徑的負相關都更為顯著,而孔體積則隨TOC增加而減小。TOC與孔隙參數的相關性變化趨勢表明有機質在向烴類轉化過程中所產生的孔隙演化過程十分復雜。結合SEM觀察結果及前人的熱模擬研究,盡管熱演化是泥頁巖中有機質孔隙形成、發展及轉化或者消失的驅動力[10],但并非唯一影響因素。前人研究也表明有機質的熱成熟過程既有新孔隙的產生,也伴隨著孔隙被石油、瀝青等填充以及孔隙之間的轉化[48]。因此,為了探討除熱成熟度以外其他影響有機質孔隙發育特征的因素,我們依據對上述四組樣品(延長組、龍馬溪組、牛蹄塘組、山西組)大量的FE-SEM觀察,通過定量統計分析來深入探討有機質孔隙發育特征的主要影響因素。
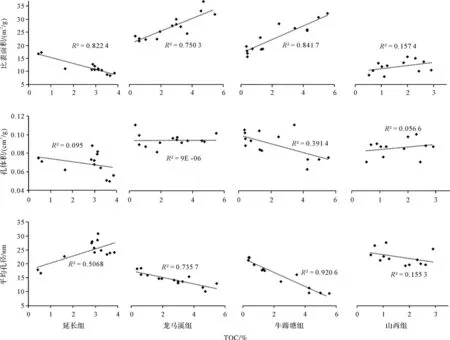
圖7 TOC與孔隙參數的相關性分析Fig.7 Correlation analysis between total organic carbon(TOC)and pore parameters
4 有機質孔隙特征的影響因素分析
四組泥頁巖的熱成熟度逐步遞增表示其處于不同的熱演化階段,但由于沉積環境、構造背景及區域性特征差異,各組泥頁巖儲層在保存條件、礦物組成、有機質豐度、類型及顯微組分等方面也存在明顯不同,進而影響著泥頁巖中有機質的生烴潛力和儲集能力。因此,各組泥頁巖中有機質孔隙發育特征必然受多重因素共同控制。通過對四組泥頁巖樣品的大量觀察分析,本文分別論述了各演化階段泥頁巖中有機質孔隙發育的主要影響因素。
4.1 延長組
為更好地對比有機質孔隙發育的差異性,我們利用Image J軟件根據灰度差異對發育納米孔隙的有機質進行了孔隙的定量統計分析。依據延長組泥頁巖FE-SEM圖像發現大多數有機質無孔隙發育(圖3),少量有機質發育有大孔、介孔,甚至微孔(圖8)(FE-SEM的分辨率不足以識別)。針對同一區域的有機質孔隙,定量統計發現孔隙發育特征有明顯差異:如圖8A所示,區域a中納米孔隙的孔徑(78 nm)是區域b(47 nm)的近兩倍,而另一個樣品中的有機質孔隙孔徑只有5 nm(表3)。由此可見,對于延長組樣品,既存在不發育孔隙的有機質,也存在孔徑差異較大的有機質孔隙。
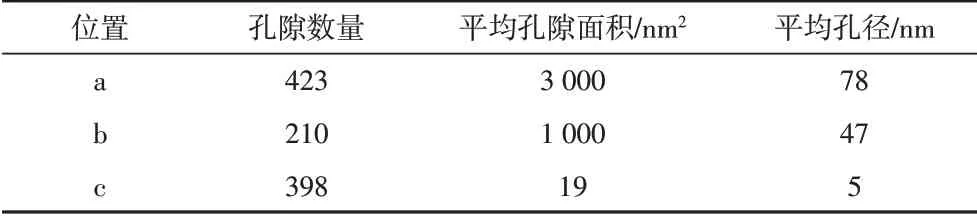
表3 圖8中三個區域的孔隙統計分析Table 3 Pore statistical analysis for the three regions in Fig.8
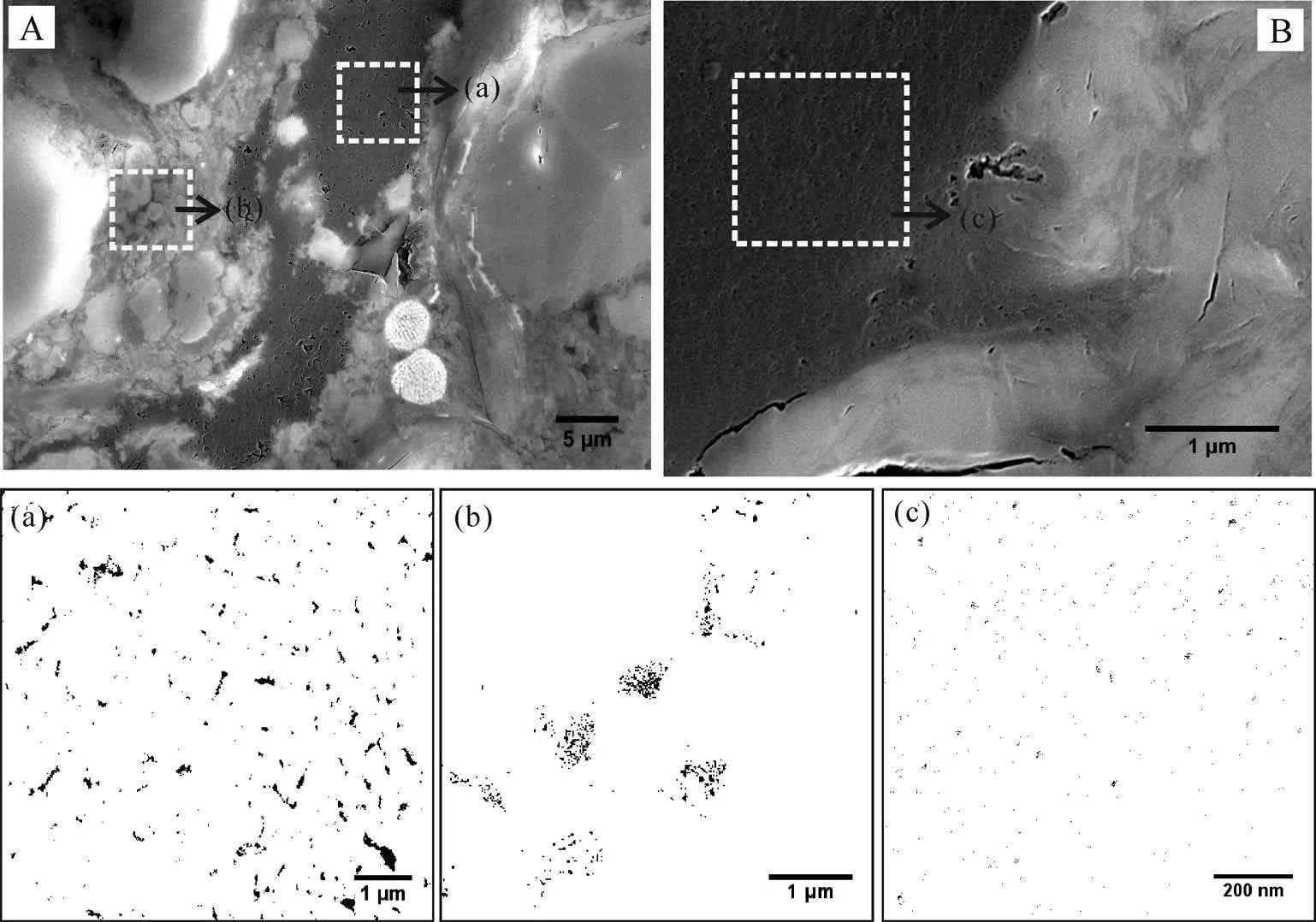
圖8 延長組泥頁巖中有機質孔隙發育特征對比分析Fig.8 Comparison of organic matter(OM)pore development in T3y shales
結合前人研究,不同類型有機質的生烴能力、生烴時限有明顯差異。根據干酪根顯微組分比例,一般將有機質分為I型、II1型、II2型、III型。I型和II型干酪根發育有機質孔隙的潛力遠高于III型。這是由于I型和II型干酪根具有比III型更好的生烴潛力[6,17-18]。延長組熱演化程度相對較低,此時干酪根類型及顯微組分對于孔隙產生的時間有重要影響。易于生烴的組分往往更早地產生孔隙,而生烴較晚或生烴能力較弱的組分則難以形成有機質孔隙。例如腐泥組和鏡質組往往比惰質組具有更好的生烴潛能,因此也更易產生孔隙[6,20,38]。延長組泥頁巖中有機質孔隙發育特征的非均質性說明孔隙產生的時間差異明顯,表明其正處于有機質孔隙的形成階段,有機質類型及顯微組分對于有機質孔隙的發育有主導作用。
4.2 龍馬溪組
泥頁巖熱演化達到一定程度時,生烴潛能較高的有機質(如I型)大量生烴會形成豐富的納米孔隙,并且隨著生烴過程的進行,孔隙結構也會隨之發生改變。龍馬溪組泥頁巖I型有機質占主導地位,普遍發育納米孔隙。通過對有機質孔隙進行統計分析發現,有機—無機相互作用嚴重影響著有機質孔隙的結構特征。根據無機礦物組分特征,可將有機—無機作用進一步劃分為有機質—脆性礦物相互作用和有機質—黏土礦物相互作用。由于有機質對應力的抵抗性較弱,有機質與脆性礦物的接觸往往造成有機質發生一定程度的變形[49]。如圖9所示,當有機質處于擠壓環境時,孔隙發育較少且孔徑很小,且沿礦物顆粒邊緣孔隙展現出定向分布的特征;當遠離應力來源的礦物顆粒時,壓應力減弱,孔隙逐漸發育,且孔徑變大。而當有機質顆粒處于拉張環境時,孔隙往往更為發育且孔徑更大。造成這種現象的原因主要有兩個:一是有機質生烴反應是一個體積增大的反應,根據化學平衡原理,壓力增大使生烴反應受到抑制,從而延遲有機質的成熟進程[50];二是在孔隙產生后,脆性礦物對有機質顆粒的擠壓造成垂直應力方向孔寬減小,易于形成定向排列,而拉張環境則更利于孔隙的生長[49]。
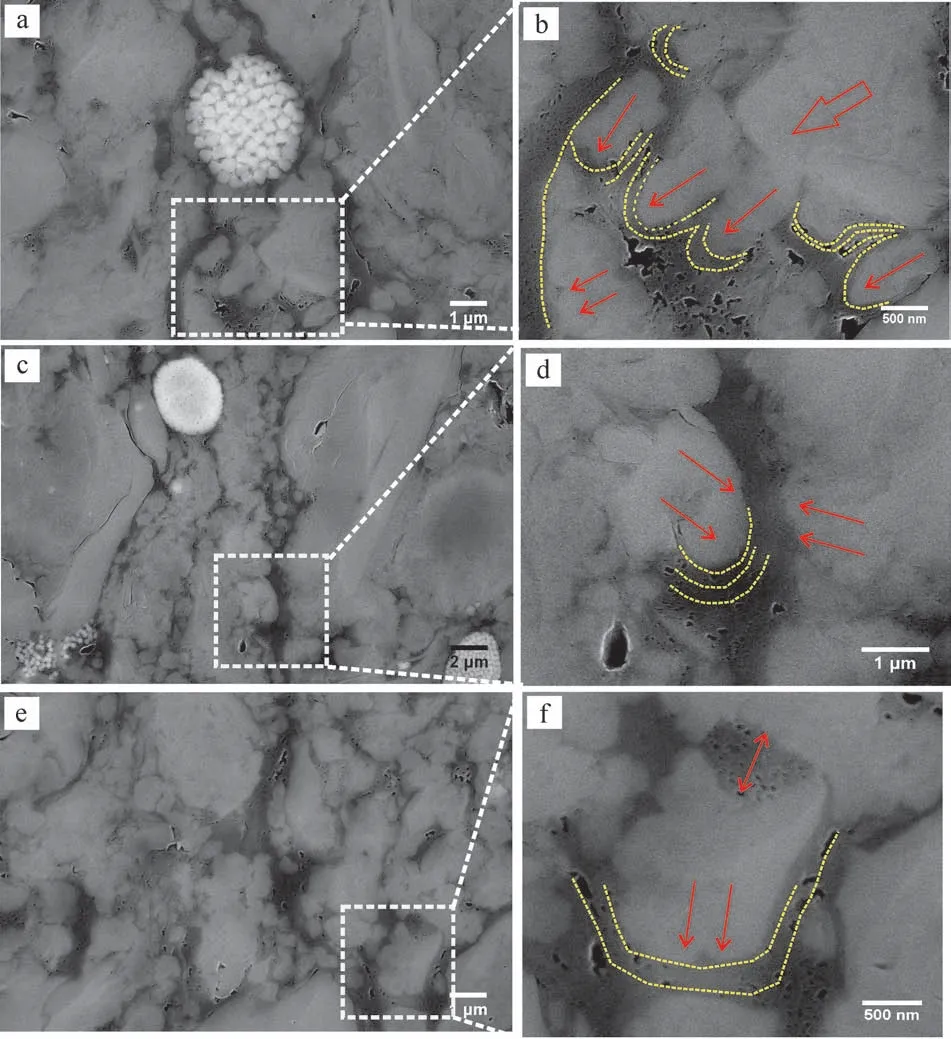
圖9 應力條件對有機質孔隙發育的影響Fig.9 Influence of stress conditions on OM pore development
泥頁巖中部分可溶有機質可與黏土礦物相互作用形成復合體,其對有機質的聚集、沉積、保存均有重要作用,是烴源巖中油氣生成的一種天然母質。有機質和黏土礦物的結合過程既有礦物的轉化(如蒙脫石的伊利石化),也伴隨著有機質的生烴,因此有機質—黏土復合體中的有機質孔隙具有比顆粒有機質孔隙更為復雜的演化過程。通過對比相同區域有機質—黏土復合體和顆粒有機質中的孔隙大小(圖10),發現顆粒有機質孔隙(29 nm和30 nm)比復合體中的孔隙(21 nm和23 nm)更大(表4)。分析認為該階段有機質—黏土復合體對于孔隙的發育起到一定的抑制作用,限制了孔隙的生長。這是因為在熱演化過程中,有機質—黏土復合體中的黏土層通過隔絕或吸收外部溫壓而抑制有機質的熱演化。泥頁巖中蒙脫石的伊利石化與干酪根生油的埋深和溫度范圍一致[51-52],且隨著熱演化程度的增加,混層礦物(伊蒙混層)中伊利石的含量逐漸增加[50,53-54],表明在這個過程中蒙脫石可吸收部分溫壓而發生礦物轉化,進而對內部有機質形成保護,不利于有機質的熱演化和孔隙生長。另外黏土層之間的狹小空間及微小的有機質顆粒也不利于納米孔隙的生長[12]。
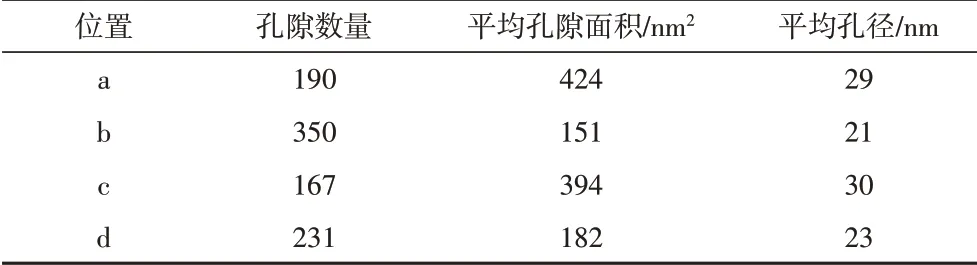
表4 圖10中四個區域的孔隙統計分析Table 4 Pore statistical analysis for the four regions in Fig.10
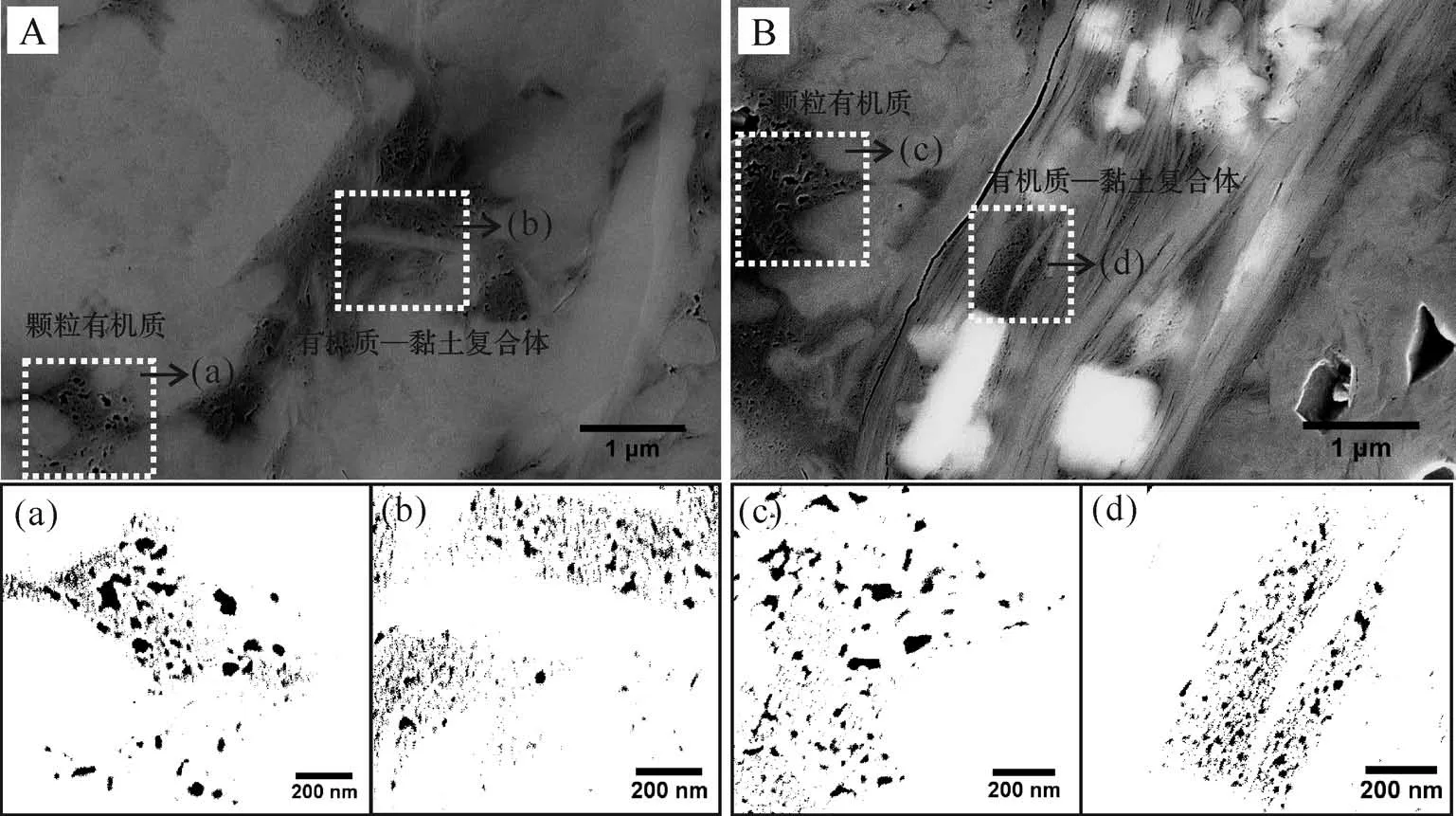
圖10 龍馬溪組泥頁巖顆粒有機質和有機質—黏土復合體中納米孔隙對比分析Fig.10 Comparison of nanopores in discrete OM and organic?clay composites for S1l shales
綜合上述分析,龍馬溪組泥頁巖處于有機質孔隙發育的高峰階段,有機—無機相互作用制約著有機質孔隙的形貌和結構特征,主要體現在兩個方面:脆性礦物對有機質顆粒所形成的應力作用和有機質—黏土復合體對孔隙生長的抑制作用。
4.3 牛蹄塘組
泥頁巖達到生烴高峰后,排烴作用開始占據主導地位,有機質孔隙中烴類的逸散加劇了這一進程[12]。相比于龍馬溪組,牛蹄塘組泥頁巖排烴作用更為完全,有機質孔隙因地層應力較大更易被壓實甚至消失[46-47]。微裂隙(成巖裂隙或構造成因裂隙)作為排烴作用的主要通道,能夠有效增加有機質孔隙的連通性,從而促進烴類逸散[55]。如圖11所示,相同區域的有機質顆粒孔隙發育差異明顯,A圖因周圍微裂隙的發育造成有機質孔隙中烴類散失,使得孔隙內部壓力降低,在地層壓力作用下而被壓實;D圖則因無微裂隙發育,孔隙仍然保持著內部壓力而免于被壓實。
此外,需要注意的是,牛蹄塘組泥頁巖中的有機質—黏土復合體也發育有大量的納米孔隙,并且復合體周圍是否存在微裂隙對孔隙發育幾乎沒有影響。通過對比統計分析有機質—黏土復合體和顆粒有機質中的孔隙大小(圖11C~F),發現兩種形態的有機質孔隙孔徑較為一致(表5)。結合龍馬溪組泥頁巖中兩種形態有機質中的孔隙分析,表明一方面牛蹄塘組中的顆粒有機質孔隙可能因壓實作用而使孔徑減小甚至消失,另一方面有機質—黏土復合體對孔隙有保護作用,分析認為在孔隙的發育過程中因黏土層的隔絕作用而降低了孔隙連通性,使有機質孔隙不會因為失去內部壓力而被壓實。另外,有機質—黏土復合體的結構特征也對孔隙有一定的保護作用,主要體現在:1)黏土層之間的表面張力。由于黏土礦物層之間相互連接,兩個黏土層之間一般為幾個納米到上百納米,使得層間的表面張力能夠對有機質形成強有力的吸附,從而使有機質不會因熱演化而收縮[56-57]。2)有機質—黏土復合體的結構穩定性。對于應力作用,復合體具有比顆粒有機質更強的抵抗能力[58],這進一步保護了復合體中的孔隙不被壓實。
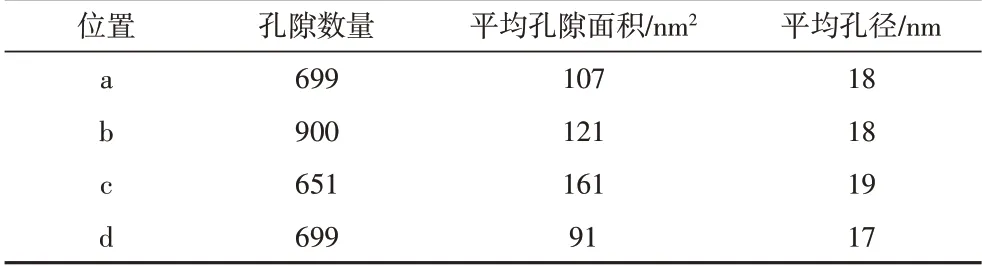
表5 圖11中四個區域的孔隙統計分析Table 5 Pore statistical analysis for the four regions in Fig.11
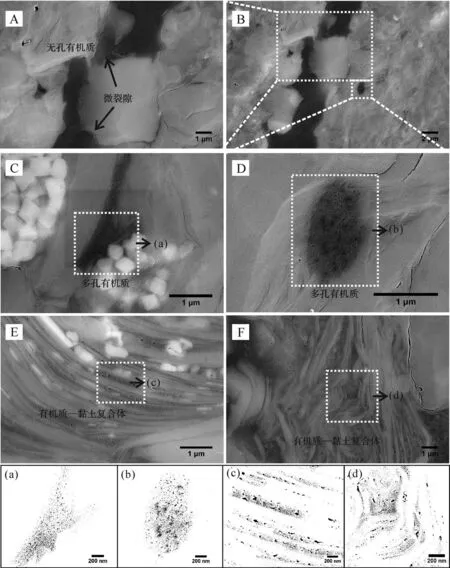
圖11 牛蹄塘組泥頁巖顆粒有機質中孔隙發育的差異性及其與有機質—黏土復合體中納米孔隙的對比分析Fig.11 The difference of pore development in discrete OM and the comparison with pores in organic?clay composites for?1n shales
綜上所述,牛蹄塘組泥頁巖處于有機質孔隙的收縮階段,顆粒有機質周圍微裂隙的發育情況(或保存條件)決定著孔隙的發育。該階段有機質—黏土相互作用在一定程度上抑制了孔隙的收縮,也是孔隙發育的主要影響因素。
4.4 山西組
豫西地區上古生界二疊系烴源巖熱成熟度普遍過高,可能與區域熱事件有關[33,38]。該地區大規模的熱事件所形成的過高的地溫促使豫西地區烴源巖快速達到過高成熟階段,并生成終極產物CH4。山西組過高的熱演化程度雖能使該地區干酪根充分反應成烴,但由于有機質類型主要為III型,生烴潛能較差[38,59],產生有機質孔隙的能力十分有限。另外,該區域烴源巖生氣高峰的時代以中侏羅世為主,三疊紀和白堊紀次之,明顯早于南方龍馬溪組海相頁巖氣古近紀的生氣高峰[33]。生氣高峰時間越早,越不利于頁巖氣的保存。豫西地區野外露頭剖面和鉆井地層中發育了大量的裂隙和斷層,FE-SEM也觀察到微裂隙十分發育(圖6a~c)。這既有可能是生烴作用產生的不規則裂隙,也可能是后期構造作用形成的規則破裂,但無論成因如何都可促使烴類的逸散[33,38]。因此即使在熱成熟過程中產生了有機質孔隙,也會在后期因保存條件差使有機質孔隙被逐漸壓實而消失。
此外,山西組過高的熱演化程度也可能引起部分有機質發生變質或結構塌陷(圖6d~f)。前人關于高演化階段有機質的力學性能研究表明有機質的楊氏模量值與化學結構參數之間存在良好的正相關性,說明有機質表面微觀力學性能受控于其內部化學結構,有機質的微觀力學性能可影響其在生氣階段生成并保留納米孔隙的能力[60]。也就是說,隨著熱成熟度的持續增加,部分有機質達到變質期,其固體干酪根和焦瀝青的物理化學性質均趨近于石墨,導致其有機質內部不發育孔隙[8]。另一方面,有機質從粘彈態演變為玻璃態,產生的氣態烴無法保留形成孔隙,內部生成的納米孔隙不斷坍塌、合并[60],最終使得有機質孔隙轉化形成大孔甚至微米級孔隙或者消失。綜上,我們認為山西組泥頁巖處于有機質孔隙的轉化和消失階段,保存條件和有機質類型及結構是該組泥頁巖有機質孔隙發育情況的主要影響因素。
5 結論
(1)延長組泥頁巖基本不發育有機質孔隙,但發育有較多的礦物粒間孔及微裂隙;龍馬溪組泥頁巖有機質孔隙極為發育,多為大孔和介孔,且形態多樣;牛蹄塘組泥頁巖中多數有機質不發育納米孔隙,部分有機質發育介孔,這種有機質孔隙發育的非均質性可能與孔隙的排烴作用是否完全有關。山西組泥頁巖部分有機質不發育孔隙,部分有機質有明顯結構塌陷,可能發生了孔隙的轉化。
(2)隨著熱成熟度的增加,孔隙參數特征與TOC的相關性有明顯變化:比表面積與TOC先呈負相關,后呈明顯地正相關,最后趨于無明顯相關性;平均孔徑與TOC表現出正好相反的相關性變化規律;而孔體積總體上與TOC的相關性較弱,可能是因為孔體積是由與礦物相關的大孔所主導。這種相關性變化體現了不同演化階段有機質對于孔隙的貢獻有明顯差異。造成這種差異的因素除熱成熟度外,還有其他因素,如有機質類型及顯微組分、有機—無機相互作用及保存條件等。
(3)四組泥頁巖儲層分別處于有機質孔隙演化的四個階段:形成階段、高峰階段、收縮階段、轉化和消失階段,各階段孔隙發育特征的主導因素不同。延長組陸相泥頁巖處于有機質孔隙形成階段,有機質類型及顯微組分主導著有機質孔隙的發育;龍馬溪組海相泥頁巖處于有機質孔隙發育高峰階段,有機—無機相互作用制約著有機質孔隙的形貌和結構特征;牛蹄塘組海相泥頁巖處于有機質孔隙收縮階段,顆粒有機質周圍微裂隙和有機質—黏土復合體的發育情況決定著孔隙的發育特征;山西組海陸過渡相泥頁巖處于有機質孔隙的轉化和消失階段,有機質孔隙是否發育以及形態特征受制于保存條件和有機質類型及結構。
致謝 感謝編輯部和審稿專家在修改過程中給予的建議和幫助;感謝河南省國土資源科學研究院谷德敏等專家在采樣過程中的幫助。
