荒漠綠洲過渡帶不同年限雨養(yǎng)梭梭(Haloxylon ammodendron)對土壤水分變化的響應
2021-09-02 01:28:02王國華陳蘊琳緱倩倩
生態(tài)學報 2021年14期
王國華,陳蘊琳,緱倩倩
1 山西師范大學,地理科學學院,臨汾 041004 2 中國科學院西北生態(tài)環(huán)境資源研究院,中國生態(tài)系統(tǒng)研究網(wǎng)絡臨澤內陸河流域研究站,蘭州 730000 3 中國科學院沙漠與沙漠化重點實驗室 中國科學院西北生態(tài)環(huán)境資源研究院,蘭州 730000
干旱地區(qū)約占全球整個陸地面積的30%,涉及到全球近20多個國家,是陸地生態(tài)系統(tǒng)重要的組成部分[1]。我國干旱地區(qū)主要包括分布在35°N以北、106°E以西的干旱荒漠地區(qū),也包括年降水量在300—400mm左右的農牧交錯地區(qū),這些地區(qū)一個顯著的地理特征是形狀和大小各異的天然或人工綠洲沿河流發(fā)育,在廣袤的荒漠中呈斑塊狀或帶狀分布[2]。近50年來,隨著我國西北地區(qū)人口快速增加,綠洲邊緣天然灌木林地和草地被大面積開墾,人工綠洲過度擴張,從而導致荒漠與綠洲之間過渡帶嚴重萎縮,綠洲邊緣人工固沙植被種植面積不斷向荒漠擴展[3]。然而,由于遠離河流,種植密度過大,加之干旱荒漠地區(qū)降水稀少且時空分布極其不均,干旱造成的土壤水分虧缺導致綠洲邊緣人工林在種植后期普遍出現(xiàn)退化現(xiàn)象[4]。
在目前全球氣候變化的背景下,北半球地區(qū)可能會面臨更多、更嚴重的極端強降水事件[5- 6],年內年際間的降水波動更加頻繁[7],區(qū)域降水不平衡和水資源短缺的局面也將更為嚴峻[8- 9],尤其是對于降水稀少、植被稀疏、生態(tài)環(huán)境最為脆弱的干旱荒漠地區(qū)[10]。盡管荒漠植物在長期進化過程中已形成一系列生態(tài)適應策略[11],但劇烈的環(huán)境變化仍有可能導致植物在生理或個體形態(tài)特征極度不適應,影響植物的生長,造成種群適合度下降,增加植物物種區(qū)域性滅絕的風險,尤其是對于干旱荒漠地區(qū)大面積種植的雨養(yǎng)固沙植物類群而言這種風險更大。
梭梭(Haloxylonammodendron)作為藜科(Chenopodiaceae)、梭梭屬(HaloxylonBge)多年生小喬木,超旱生、耐鹽、抗風蝕,在中亞地區(qū)和我國西北干旱荒漠、半荒漠地區(qū)自然分布極廣[12]。梭梭,作為優(yōu)秀的防風固沙先鋒植物種,在我國西北荒漠地區(qū)種植面積十分龐大,僅僅在阿拉善地區(qū),“螞蟻森林”就計劃種植一億棵梭梭。然而近幾十年來我國西北干旱荒漠地區(qū),例如新疆區(qū)甘家湖天然梭梭林國家自然保護區(qū)[13],古爾班通古特沙漠[14]、張掖[15]和民勤[16]綠洲邊緣等,天然梭梭和人工種植梭梭都普遍出現(xiàn)大面積衰退和死亡的現(xiàn)象[17],生理和生長形態(tài)變化及其對梭梭生長適合度敏感性影響一直被視為是其脆弱性評估的重要指標,一直是梭梭研究關注的焦點問題之一。
近年來,學者們主要對梭梭抗旱性[18]、鹽旱脅迫[19]、種子萌發(fā)特性[20]、梭梭幼苗存活[21]、梭梭生理特征與環(huán)境的關系[22]、以及梭梭的光合水分關系[23]等方面開展了大量研究,為揭示梭梭生存和退化機制以及群落穩(wěn)定性提供了大量依據(jù)。例如,李彥、徐皓等[24- 26]研究發(fā)現(xiàn),梭梭主要利用淺層土壤水分維持生長和存活;許強[27]通過研究梭梭不同生長階段的分支特征,認為梭梭主要通過形態(tài)建成改變來適應環(huán)境脅迫;田媛[28]研究梭梭種子萌發(fā)到梭梭定居的過程,表明梭梭個體形態(tài)調整對其幼苗生長存活至關重要;趙文智[29]等從不同年限的梭梭種群尺度說明了梭梭種群的適應性演變特征;周海[30]等發(fā)現(xiàn)梭梭根系空間分布具有顯著的二態(tài)性特征:梭梭具有廣泛分布的表層和淺層根系,能夠大量吸收由降水和凝結水補給的淺層土壤水分;另一方面,梭梭具有發(fā)達的主根系,在地下水較淺的生境下可以利用地下水來滿足植物的水分生理需求;王亞婷[31]等研究表明,梭梭對5mm小降水沒有明顯的生理響應;而吳玉[32]等對1mm單次降水對梭梭影響的研究發(fā)現(xiàn),梭梭可以間接利用小降水。以上研究認為梭梭通過較強的氣孔控制機制和個體形態(tài)調節(jié)來適應干旱、鹽分等脅迫環(huán)境。但是,關于梭梭的生理和個體適應機制也存在不同的觀點和認識,這可能與梭梭的林齡、具體生境差異有關。同時,人工固沙植被對干旱的適應過程是一個連續(xù)長期的過程,然而迄今為止,尚未有研究系統(tǒng)論述梭梭退化死亡原因及其與環(huán)境脅迫因素的關系,也并未充分考慮不同生境下或不同生長階段植物生理特性與適應對策之間、生理特征與個體形態(tài)之間的復雜聯(lián)系。
河西走廊作為我國西北主要的糧食生產基地,地處我國內陸干旱荒漠地區(qū),氣候干燥,風沙活動強烈,生態(tài)環(huán)境脆弱,是我國風沙活動危害最為嚴重的地區(qū)之一,也是西北地區(qū)建立防風固沙生態(tài)屏障的重點區(qū)域[33]。多年來為了有效遏制風沙危害,該地區(qū)開展了一系列以人工植被建設為主要生態(tài)修復措施的生態(tài)建設工程,有效促進了局地生境恢復[34]。梭梭作為當?shù)仃P鍵的人工固沙植被建群種,種植面積最大,分布范圍最廣,是維持和保護河西走廊荒漠綠洲穩(wěn)定的關鍵植被。近50a來,隨著綠洲面積不斷擴張,荒漠綠洲過渡帶的人工梭梭生境破碎化嚴重,種植梭梭大量出現(xiàn)退化、死亡以及種群無法天然更新等問題,這直接影響了綠洲邊緣風沙區(qū)生態(tài)恢復以及人工固沙林防風固沙效益的可持續(xù)性[35]。但目前對于人工梭梭林長期觀測數(shù)據(jù)較少,對不同種植年限梭梭生理和生長特征及其與主導制約因素(土壤水分)的關系還并不清楚,這不僅影響著利用特有植物建立固沙植被實踐的開展,也是荒漠生態(tài)系統(tǒng)生態(tài)水文研究的知識缺陷。本文擬通過在5、10、20、30和40a固沙梭梭林地調查取樣,還原一個長時間序列的梭梭人工林的土壤和植被變化過程,分析不同種植年限梭梭不同土壤干旱條件下梭梭的生理和形態(tài)特征,并闡明梭梭生理和個體形態(tài)對不同水平土壤干旱的適應機制,研究結果有助于更加準確地預測土壤干旱對荒漠綠洲過渡帶人工固沙植被的潛在影響。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于甘肅省河西走廊臨澤縣北部荒漠綠洲邊緣,地處巴丹吉林沙漠邊緣和臨汾綠洲的交匯處(39°09′—39°21′N,100°02′—100°21′E,海拔1350—1367 m),是風沙入侵臨澤綠洲的主要通道。氣候屬于典型的溫帶大陸性干旱荒漠氣候,年溫差大,年均溫為7.6℃;年均降水量為116.8 mm,降水集中于7—9月份;日照充足,晝夜溫差大;春季風沙活動強烈,主要集中于3—5月,以西北風為主,最大風速可達21 m/s;土壤類型主要為非地帶性的風沙土、鹽土。綠洲邊緣沙丘上主要分布有沙拐棗 (Calligonummongolicum)、紅砂(Reaumuriasoongorica)、泡泡刺 (Nitrariasphaerocarpa)等天然植被,梭梭是人工固沙植被的建群種和優(yōu)勢種,梭梭林下常常伴生有一年生草本植物例如霧冰藜 (Bassiadasyphylla)、沙米(Agriophyllumsquarrosum)、刺沙蓬(Salsolaruthenica)等。從20世紀70年代開始,綠洲邊緣開始大面積種植梭梭防治沙漠化,并在2000年以后,又在荒漠進行了多期的固沙造林建設,逐漸形成了從綠洲到荒漠大約5km×10km的人工固沙植被帶,同時也為本研究提供一個長時間序列梭梭防護林體系。
1.2 調查取樣
植被調查于2019年7—9月進行,在種植5、10、20、 30和40a的典型梭梭林內選擇3個典型梭梭種植樣點作為植物和土壤取樣點,同時,在未栽種梭梭的流動沙地作為對照樣點(0a)。由于梭梭都栽種在流動沙地,土壤質地、養(yǎng)分和結構基本一致。取樣方法采用巢式取樣法,在不同種植年限梭梭林每個樣點設置3個25×25 m2的樣方,樣方位置均為平坦沙地,且每個灌木樣方之間的距離大于20m。在每個樣方內,各選取5株長勢良好,高度冠幅接近于樣方內平均高度冠幅的林內植株,同時避免邊緣效應,摘取每株梭梭的新葉(即綠葉)與老葉(即黃葉),保證葉片足量,且上中下部位的葉子均勻采集(從20a開始新葉即綠葉主要集中在梭梭上部,老葉即黃葉集中在梭梭的下部,圖1);不同種植年限梭梭的根系調查主要通過挖土法完成,在5株梭梭取樣植株根莖附近分層提取,每層深度為10 cm,提取兩次取到20cm。取樣后篩出健康的毛細根,用游標卡尺測量根系直徑,將小于等于2mm的根系認定為吸收根系。將新鮮葉片(新葉和老葉)和毛細根均勻保存在鋪有干冰的泡沫保溫箱內,以備實驗室測定葉片和根系各項生理指標。記錄取樣梭梭植株個體的株高、冠幅、莖干枝條生物量、葉片生物量和枯枝比例。生物量采用烘干法測定,即將取回的植物樣品放入定溫80℃的烘干箱內烘干至恒重。

圖1 不同種植年限的梭梭生長狀態(tài) Fig.1 Growth status of Haloxylon ammodendron with different plantation ages 新葉即綠葉,主要集中在梭梭植株上部,老葉即黃葉,主要集中在梭梭植株下部;從30a開始,梭梭枯枝落葉明顯增加,40a梭梭生物量明顯減少
1.3 土壤水分測定與植物樣品測定
在5、10、20、 30和40a梭梭林內選擇3個典型梭梭定位樣點,在梭梭植株莖干10cm左右用土鉆取不同土壤深度20—40cm,100—120cm和180—200cm土壤,放入鋁盒,然后將鋁盒帶回實驗室立即稱重,在105℃恒溫箱內烘干至恒重,移到干燥器內冷卻至室溫,稱重,通過計算得到土壤水分含量,同時,在流動沙丘取樣作為對照(0a),并結合中國科學院臨澤內陸河流域研究站對20a梭梭林齡固定監(jiān)測樣點連續(xù)近20年土壤水分監(jiān)測,還原0—40a人工雨養(yǎng)梭梭林土壤水分狀況特征。
梭梭葉片(新葉和老葉)生理指標測定:選取植物上部和下部枝條上的葉片,立即稱量新鮮樣品0.1g,剪碎,用95%乙醇和80%丙酮(1:1)混合溶液室溫黑暗浸提24h直接測量,提取液分別在波長663nm和645nm下測定吸光度,通過Lambert-Bee定律計算出提取液中葉綠素a、b 含量的濃度(mg/g);葉片(新葉和老葉)滲透調節(jié)物質指標用蒽酮比色法測定可溶性糖含量;用考馬斯亮藍G- 250法測定可溶性蛋白含量;用茚三酮顯色法測定游離脯氨酸含量;采用硫代巴比妥酸法測定丙二醛(MDA)含量;根系生理指標測定:用TTC法測定0—10cm和10—20cm毛細根的根系活力。
1.4 數(shù)據(jù)處理與分析
實驗數(shù)據(jù)分析采SPSS21軟件進行數(shù)據(jù)整理和分析,通過在One-Way ANOVA在95%的置信水平上,采用Duncan法進行顯著檢驗。文中所有圖均利用Origin8軟件完成。
2 結果
2.1 不同年限梭梭林土壤水分變化
隨著梭梭種植年限的增加,沙土淺層20—40cm土壤水分呈波動狀態(tài),土壤水分含量保持在2%—3%左右;而在100—120cm和180—200cm,土壤水分在前期(0—20a)基本保持在3%—4%左右,而從30—40a開始土壤水分下降到只有1%—2%左右,并且保持低含水量穩(wěn)定狀態(tài)。
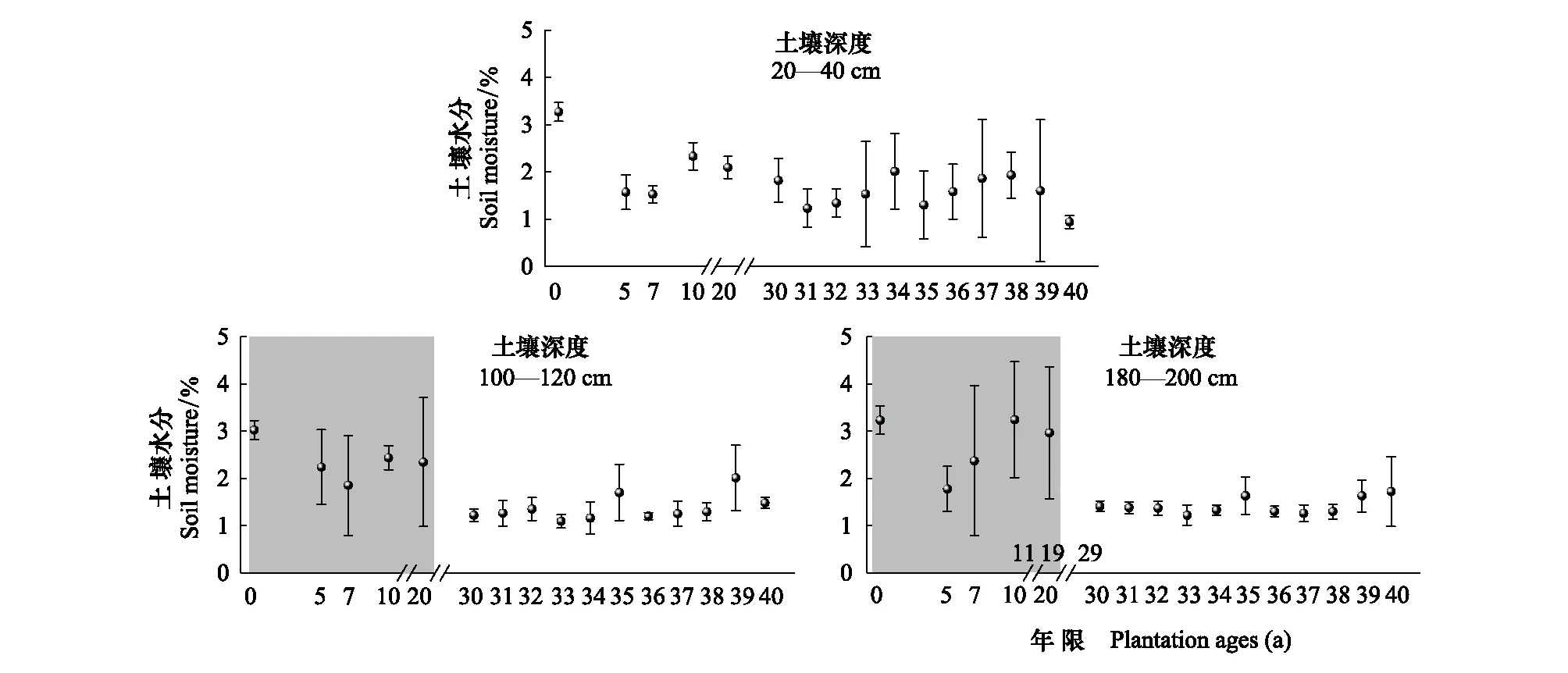
圖2 不同種植年限梭梭林不同土壤深度土壤水分變化Fig.2 Soil moisture changes in different soil depths of Haloxylon ammodendron with different plantation ages
2.2 不同年限梭梭葉片和根系生理特征
2.2.1葉片滲透調節(jié)物質含量變化
不同年限梭梭葉片(新葉和老葉)的滲透調節(jié)物質可溶性糖和可溶性蛋白含量差異顯著;可溶性糖和可溶性蛋白含量在5—20a含量較低,而在30—40a時含量較高。游離脯氨酸含量變化未達到顯著水平,但也呈現(xiàn)和其他兩種滲透調節(jié)物質相類似的結果,在20—40a時含量處于高值。同時,本文發(fā)現(xiàn)在0—40a中新葉的滲透調節(jié)物質含量要普遍低于老葉(圖3)。
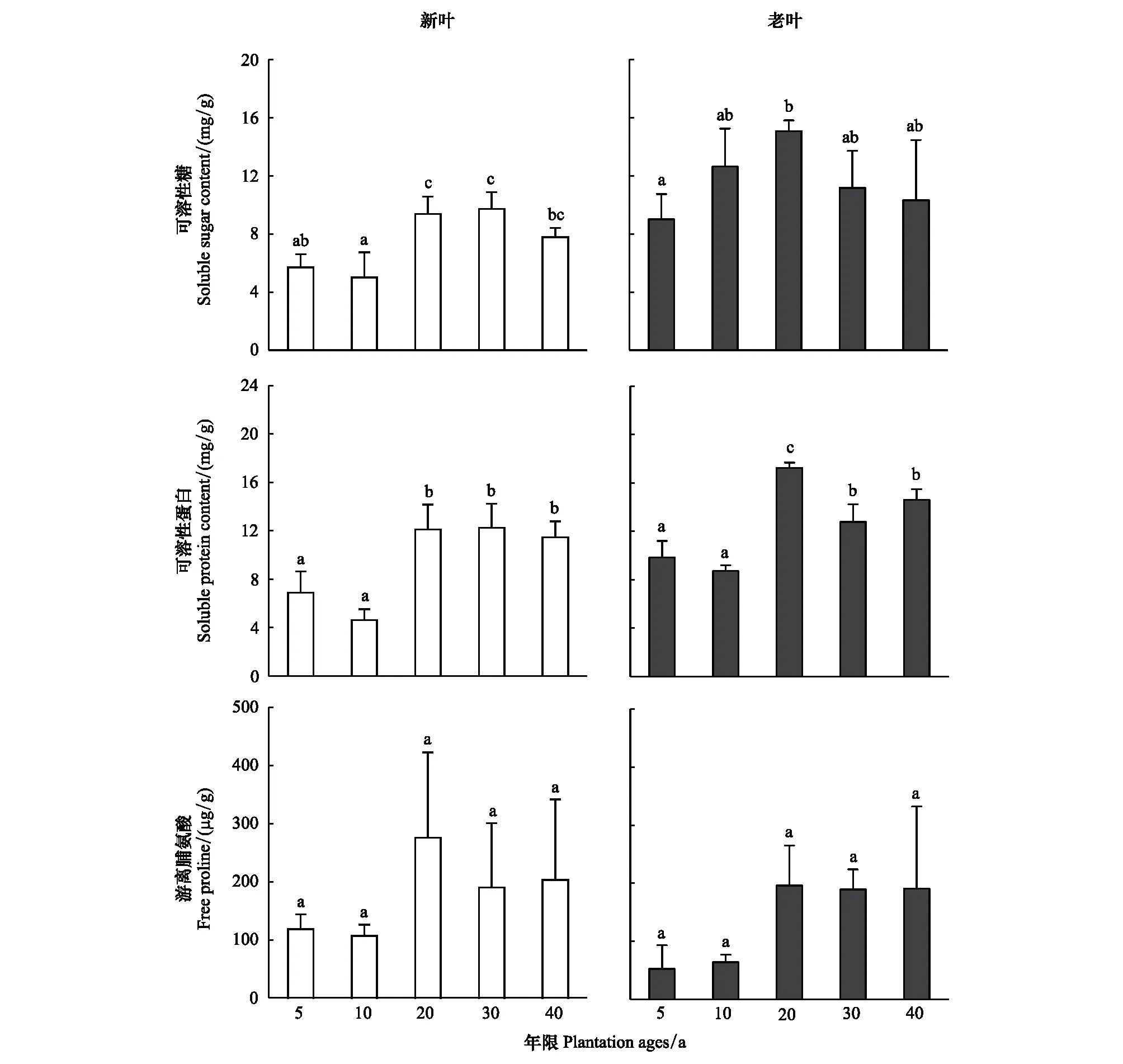
圖3 不同種植年限梭梭新葉和老葉葉片可溶性糖、可溶性蛋白和游離脯氨酸含量變化Fig.3 Changes in soluble sugar, soluble protein and free proline contents in young leaves and old leaves of Haloxylon ammodendron at different plantation ages不同字母代表葉片在不同年限的差異顯著,P<0.05
2.2.2葉片丙二醛含量變化
不同年限梭梭新葉丙二醛含量遠遠低于老葉含量,新葉丙二醛含量基本保持在0.01umol/g,而老葉除了在20a保持在較低水平,其他年限梭梭老葉丙二醛含量基本在0.16—0.02 umol/g左右(圖4)。
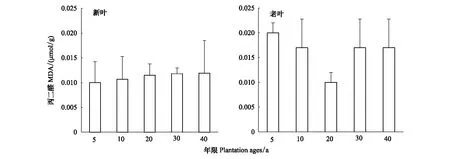
圖4 不同種植年限梭梭新葉和老葉葉片丙二醛含量變化Fig.4 Changes in MDA in young leaves and old leaves of Haloxylon ammodendron with different plantation ages
2.2.3葉片葉綠素含量變化
不同年限梭梭新葉葉綠素含量遠遠高于老葉含量,新葉總葉綠素含量保持在400—600 mg/g,而老葉總葉綠素含量低于100mg/g。同時,新葉葉綠素a、b和總含量都隨著種植年限的增加而呈現(xiàn)顯著的增加,而老葉葉綠素含量都保持在較低的水平,沒有顯著變化(圖5)。
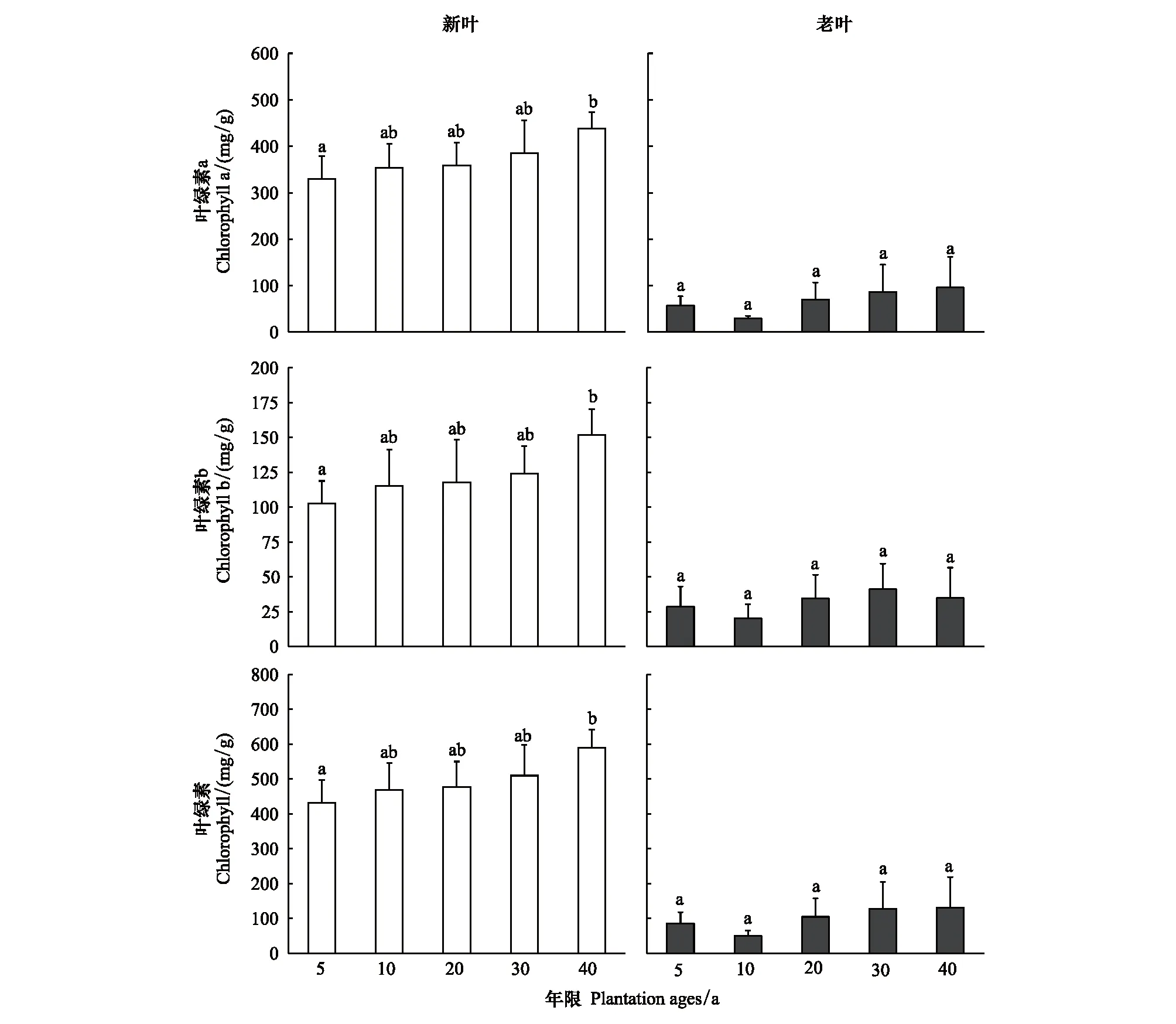
圖5 不同種植年限梭梭新葉和老葉葉綠素a 、b和總含量的變化Fig.5 Changes in chlorophyll a, b and total contents of young leaves and old leaves of Haloxylon ammodendron with different plantation ages
2.2.4根系活力含量變化
不同年限梭梭,0—10cm土壤深度的根系活力存在顯著差異(P<0.05),根系活力的變化趨勢近似拋物線型:5—20a間整體是增加的趨勢,20a時梭梭的根系活力達到了最大值(為8.51mg/g),30—40a逐年下降,40a時根系活力最低(1.20mg/g);10—20cm土壤深度的根系活力變化幅度較小,5—20a相對來說是高值區(qū),10a時達到最大值(為3.42mg/g),30—40a是低值區(qū)(圖6)。
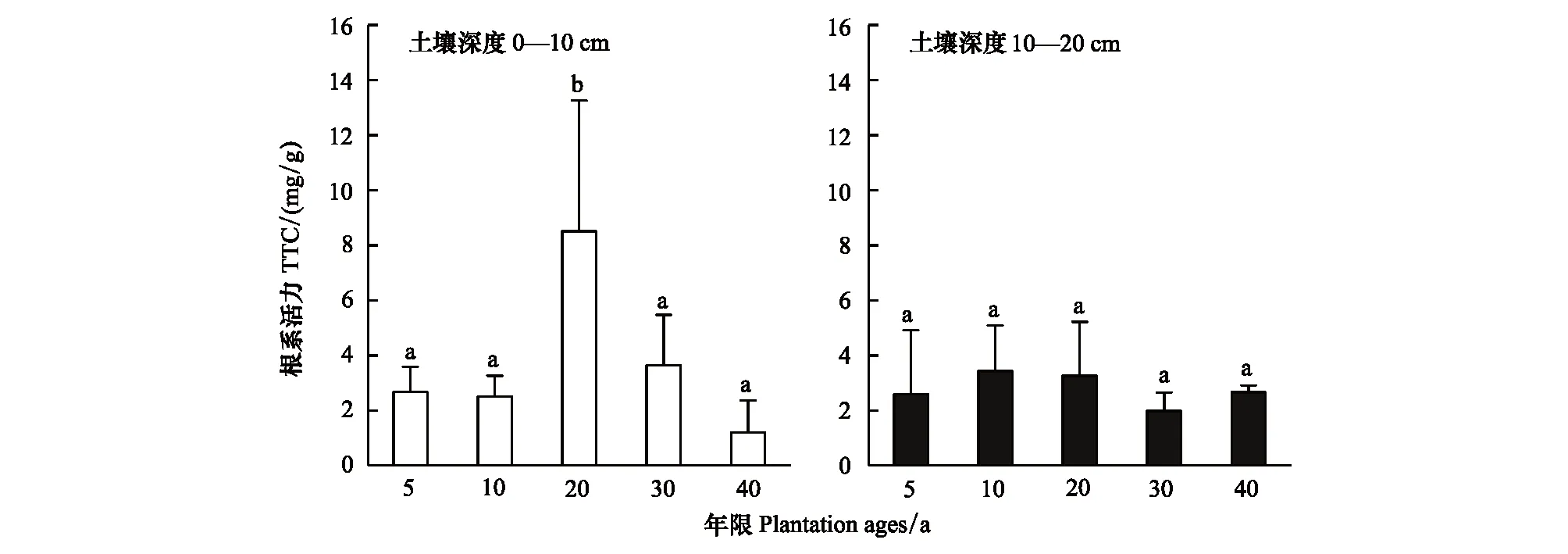
圖6 不同種植年限梭梭根系活力的變化Fig.6 Changes in feeder root activity of Haloxylon ammodendron with different plantation ages at 0—10 cm and 10—20 cm
2.3 不同年限梭梭個體生物量和形態(tài)變化
2.3.1個體生物量
單株梭梭新葉老葉總生物量的變化趨勢是:先增加后減少,5—30a不斷增加,在20—30a時達到最大值(588 g),40a迅速下降,在40a葉片總生物量最低(為218 g)。單株新葉生物量在5—30a逐年上升,在20—30a時達到了最大值(為205 g),到40a時迅速下降到了最低值(為56 g);單株老葉生物量在5—30a持續(xù)上升,30a時達到了峰值(401 g),在40a時下降到了最低值(163g)。單株梭梭枝條生物量的變化呈現(xiàn)先增加后減少的趨勢,5—20a間不斷上升,5a時生物量最小,在20a時達到了最大值,20—40a階梯式下降;單株梭梭莖干生物量的變化趨勢也是:先上升后下降,5—30a間呈現(xiàn)不斷上升的趨勢,5a時生物量最小(85g),在達到20—30a時快速生長,莖干喬木化,30a時生物量達到了最大值(8742 g),40a莖干干枯,生物量嚴重下降(891g)(圖7)。
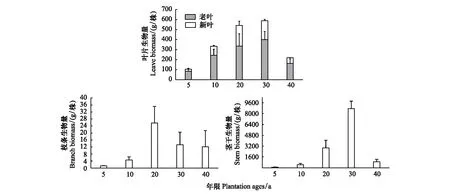
圖7 不同年限梭梭葉片、枝條、莖干的單株生物量(平均值±標準誤差)Fig.7 The biomass of leaves, branches and stemsof Haloxylon ammodendron with different ages (mean±SD)
2.3.2個體形態(tài)變化
梭梭株高的變化呈現(xiàn)不斷上升的趨勢,5—20a上升幅度較大,20—40a上升幅度較緩,最大值是在40a(為379cm);梭梭冠幅的變化趨勢,先上升后下降,5—20a間呈上升趨勢,20a時是最大冠幅(為4.94m2),20—40a間梭梭冠幅緩慢下降;梭梭枯枝比的變化呈現(xiàn)不斷上升的趨勢,5a時梭梭枯枝比最低(35%),40a時達到最大比例(74%)(圖8)。
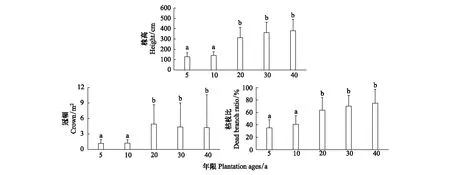
圖8 不同年限梭梭個體形態(tài)(高度、冠幅、枯枝比)的變化(平均值±標準誤差)Fig.8 Changes of individual morphological characteristics (height, crown and ratio of dead branches) of Haloxylon ammodendron with different plantation ages (mean ± SD)
3 討論
3.1 不同種植年限人工梭梭林土壤水分變化
本研究發(fā)現(xiàn)種植30a后,雨養(yǎng)梭梭林內100—120cm和180—200cm土壤含水量下降明顯,從3%—4%下降到1%—2%;而20—40cm土壤含水量呈波動狀態(tài),無明顯下降。和本研究相類似,馬全林[36]等研究發(fā)現(xiàn)在民勤人工梭梭林土壤含水量在種植30a后,林地土壤含水量保持在1%左右。一般認為,人工梭梭林土壤干燥化的原因主要有兩個;一方面,干旱荒漠地區(qū)降水稀少,而蒸發(fā)強烈,同時降水主要以小降水事件為主,降水入滲主要分布在60cm以內,淺層土壤能夠得到降水補給,而深層土壤水分補給不顯著[37];同時,梭梭的蒸騰耗水量遠大于降水量[38],尤其是在梭梭種植30a后,梭梭根系對根際土壤水分的過度消耗利用,進一步導致人工梭梭林土壤水分狀況惡化[39]。
3.2 不同種植年限梭梭生理特征
葉片,作為植物與空氣接觸總面積最大的器官,對環(huán)境脅迫因子的敏感性和感知力最強,對抵抗外界脅迫和實現(xiàn)抵抗后的功能恢復有著重要的作用[40]。本研究發(fā)現(xiàn),5—20a梭梭生長前期,土壤水分保持在3%—4%,梭梭葉片各滲透調節(jié)物質的含量整體不斷上升積累,在20a的時候達到峰值,而從30a土壤水分下降到1%—2% 時,葉片滲透調節(jié)物質并沒有顯著提高,這暗示著在前期5—20a,梭梭葉片主要通過滲透調節(jié)抵御干旱脅迫,而到了后期30—40a葉片滲透調節(jié)功能下降,由此可以看出,梭梭葉片滲透調節(jié)能力是有限的,只能在一定的干旱脅迫閾值范圍內發(fā)揮緩沖作用。
同時,本研究發(fā)現(xiàn)在種植前期5—20a,梭梭老葉的可溶性糖、可溶性蛋白的含量是新葉含量的近2倍,這說明老葉水分含量較低,滲透調節(jié)主要發(fā)生在老葉。雖然很多研究認為滲透調節(jié)物質可溶性糖、可溶性蛋白對植物是有益的,但過度的滲透調節(jié)物質可能會導致植物葉片中蛋白質、葉綠素和酶等其他有機物質減少,特別是與光合作用相關的物質,例如葉綠素a、b,因此,葉片滲透調節(jié)物質可溶性糖、可溶性蛋白積累的過程其實也是葉片退化的過程,即新葉(綠葉)向老葉(黃葉)轉化的過程。
丙二醛是植物在逆境下產生的膜質過氧化的主要產物之一,對細胞產生毒害,造成細胞膜的功能紊亂。它的積累會對植物造成一定的傷害,因此其含量可以反映植物遭受干旱傷害的程度,即膜脂過氧化作用越強,丙二醛的含量越高,則對葉片的損傷就越大[41]。本研究發(fā)現(xiàn):不同年限梭梭新葉丙二醛含量保持低水平,說明梭梭在新葉狀態(tài)下,由于葉片滲透調節(jié)物質的作用,梭梭葉片的細胞膜是完整的,而在老葉中丙二醛積累比新葉高60%—100%,說明老葉受到干旱脅迫的影響更大,老葉原生質膜結構受到破壞遠遠大于新葉。
葉綠素作為植物葉綠體類囊體膜上重要的光合吸收和轉化分子,是植物光合結構的重要組成部分,能靈敏反映光合作用的變化情況, 為植物抗逆生理、作物增產潛力預測等方面的研究提供依據(jù), 因而被視為揭示植物光合作用與環(huán)境關系的內在探針[42]。通常,干旱脅迫會直接損傷植株的生理代謝, 導致葉綠素含量減少、光合作用下降、植株生長受阻,進而影響植物正常生長發(fā)育。本研究發(fā)現(xiàn),梭梭新葉和老葉的葉綠素a、葉綠素b及總葉綠素含量隨著年限的增加不斷增加,說明梭梭在生長過程中葉片光合能力不斷提高;但同一年限的梭梭新葉老葉的葉綠素a、葉綠素b及總葉綠素含量對比差異明顯,新葉的葉綠素含量接近于老葉的5倍,這說明梭梭主要光合發(fā)生在新葉,而老葉的光合作用能力相對來講是很低的。同時通常植物葉綠素a∶b的值約為3∶1[43],而本研究發(fā)現(xiàn),不同年限梭梭新葉的葉綠素a∶b的值約為3∶1,而老葉的葉綠素a∶b的值下降到了2∶1,這也說明老葉葉綠素比例失調,光合作用下降。這些現(xiàn)象表明了梭梭應對土壤干燥的策略,即盡量保持或提高植株上部新葉的葉綠素含量,保持新葉較高的光合能力,而生理脅迫主要發(fā)生在光合作用較弱的老葉,這樣一方面可以減少蒸騰水分消耗,抵御干旱,另一方面提高梭梭個體光合作用水分利用效率,保證梭梭正常生長和生理過程所需要的水分和生物量積累。
本研究還發(fā)現(xiàn),在種植前期5—20a梭梭隨著種植年限的增加,表層土壤(0—10cm)根系活力明顯增加,在20a時達到了最大值,說明20a的梭梭根系在表層土壤中是最有活力,對降水吸收能力增強,而在后期30—40a,表層根系活力下降,對降水吸收能力減弱,因此干旱脅迫進一步加劇。
3.3 不同種植年限梭梭個體生長和形態(tài)特征
在干旱脅迫下, 植物的生長和個體形態(tài)會發(fā)生一系列調整。很多的相關研究表明,干旱脅迫會對樹木的生長形態(tài)產生抑制作用,其株高、基徑、冠幅面積等指標成為衡量抗旱的能力的重要參考[44- 45]。本研究發(fā)現(xiàn),在種植前期,梭梭株高、冠幅、生物量隨著種植年限的增加而增加,20年的梭梭是其生長過程中的最佳時期,在種植后期30—40年,梭梭老葉(黃葉)比例增加,葉片和枝條也出現(xiàn)了大量脫落的現(xiàn)象。梭梭老葉黃化和同化枝脫落,一方面保證梭梭正常的水分生長代謝,并減少低效的光合器官(老葉)水分消耗,另一方面保證新葉(綠葉)生物量,提高光合水分利用效率,這與許皓[26]的觀點較為一致。
梭梭在干旱荒漠環(huán)境下長期的生態(tài)適應過程中,不斷進化,逐漸形成了適應荒漠干旱生境的生態(tài)型。在本研究中,我們發(fā)現(xiàn)當梭梭種植30—40a土壤水分下降到1%—2%時,梭梭枝條葉片大量脫落減緩了蒸騰作用水分過度消耗,維持植物體內水碳平衡。但同時同化枝條和葉片大面積脫落也導致光合作用急劇下降,個體生物量顯著降低。這與趙興梁對沙坡頭固沙植物沙土水分研究結果類似,即在沙漠環(huán)境下土壤水分在2%以上時,固沙植物生長正常;而當土壤水分在1%—2%時,固沙植物出現(xiàn)衰退現(xiàn)象;而在1%左右固沙植物大量死亡。盡管梭梭可以用凋落葉片和枝條的方法來降低老化光合器官水分消耗,但老葉、枝條以及莖干水分匱缺極其嚴重,基本已經達到了極限,而大量葉片和枝條脫落進一步減少其個體生物量,惡化其生存能力,最后莖干也由于植株體內的水分過少也出現(xiàn)干枯斷裂。因此,基于以上研究結果,我們提出在梭梭種植30—40a后通過適度人工干預,例如修剪枯枝、適當灌溉、一定程度的平茬或間伐,可能會對人工雨養(yǎng)梭梭復活和復壯具有積極的意義。
4 結論
本文研究發(fā)現(xiàn),在典型荒漠綠洲過渡帶種植梭梭5—20a,雨養(yǎng)林土壤水分保持在3%—4%,梭梭葉片滲透調節(jié)物質和葉綠素含量的顯著增加,說明梭梭內在水分利用效率(WUE)隨著干旱程度增加而明顯提高。但是,種植后期30—40a,土壤水分降低到1%—2%,嚴重干旱造成葉片葉肉結構不可復原的破壞,當葉片水勢下降到一定閾值范圍內,葉片葉綠素含量下降,葉片生理調節(jié)失效,葉片開始死亡并隨后脫落。最后在種植40a后,多數(shù)梭梭進入休眠狀態(tài)(假死)。
