補播羊草和黃花苜蓿對退化草甸植物群落特征的影響
2021-09-07 05:57:00劉玉玲王德平楊合龍王開麗戎郁萍
草地學報 2021年8期
孫 偉, 劉玉玲, 王德平, 趙 敏, 楊合龍, 宋 倩, 肖 紅, 王開麗, 曹 婧, 戎郁萍*
(1.中國農(nóng)業(yè)大學草業(yè)科學與技術(shù)學院, 北京 100193; 2. 包頭市園林綠化事業(yè)發(fā)展中心, 內(nèi)蒙古 包頭 014030)
補播是治理退化草地的有效手段,人為添加適應(yīng)當?shù)赝嘶莸厣鷳B(tài)系統(tǒng)自然條件的、有價值的優(yōu)良牧草,增加其種類和比例,控制雜草數(shù)量及入侵速度,達到提高草地生產(chǎn)力和改善牧草品質(zhì)的目的,在維持植被生產(chǎn)力和植物群落更新等方面具有重要作用[1-2]。補播恢復退化草地實踐過程中,適宜補播時間能為草種萌發(fā)提供良好的水熱條件,且有利于補播幼苗避開不良氣候,但由于區(qū)域氣象條件的差異,不同地區(qū)補播時間的選擇也有所不同,如地中海地區(qū)適宜的補播時間應(yīng)避開10—11月和4—5月[3],而寧夏適宜補播時間為4月30日—6月30日[4]。補播比例則主要影響植物群落生物量,比例過低會導致生長空間占據(jù)少,過高則會導致植物種群激烈競爭,抑制單個植物產(chǎn)量和生長,甚至造成植物死亡[5]。國內(nèi)外學者在不同地區(qū)探討了牧草補播比例對植物群落生物量的影響,均得到了適宜本地區(qū)最優(yōu)牧草補播比例,如張翊碧等[6]在貴州的研究發(fā)現(xiàn),以2∶1∶1的鴨茅(DactylisglomerataL.)+白三葉(TrifoliumrepensL.)+多年生黑麥草(LoliumperenneL.)混播的草地草產(chǎn)量最高;韓學明[7]在青海省的研究表明,5∶3的燕麥(AvenasativaL.)和箭筈豌豆(ViciasativaL.)混播后增產(chǎn)明顯;Hodgson[8]在阿拉斯加發(fā)現(xiàn),豌豆(PisumsativumL.)與燕麥以1∶1和13∶7的混播比例產(chǎn)草量最高。綜上,補播時間和補播比例的選擇是補播成功的關(guān)鍵。
羊草為禾本科(Gramineae)多年生草本,具有無性繁殖能力強、適口性好的特點,是北方草原區(qū)的優(yōu)勢植物[9],黃花苜蓿為豆科(Leguminosae)多年生草本,是紫花苜蓿(MedicagosativaL.)的近緣野生種,因其抗逆性強、營養(yǎng)品質(zhì)高,被廣泛種植于自然條件惡劣的地區(qū)[10]。王殿才[11]等通過補播羊草恢復齊齊哈爾重度退化草地,2—3年后植被覆蓋度和干草產(chǎn)量均比對照區(qū)增加60%以上;劉暢[12]在松嫩平原補播羊草后發(fā)現(xiàn),植物群落穩(wěn)定性增強,雜類草入侵速度降低;周冀瓊[13]在呼倫貝爾草原補播黃花苜蓿后發(fā)現(xiàn),2014—2016草地生產(chǎn)力平均提高34%,效果明顯。
呼倫貝爾草原生物多樣性豐富,生產(chǎn)力高,生態(tài)服務(wù)價值大,是我國北方重要的生態(tài)屏障,但近年來由于氣候變化、農(nóng)業(yè)開發(fā)和建設(shè)用地等原因,2012年該地區(qū)退化草地面積已達88%以上[14-15],草地修復尤為迫切。近年來關(guān)于補播恢復退化草地的研究已有諸多報道,但不同補播時間和補播比例及其交互作用對退化草地植物群落的影響未見報道,因此,本文通過系統(tǒng)性研究補播時間及補播不同比例羊草和黃花苜蓿對植物群落特征的影響,旨在為呼倫貝爾退化草地修復提供依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
研究區(qū)位于內(nèi)蒙古呼倫貝爾市陳巴爾虎旗特尼河九隊(49° 26′ 17″N,120° 09′ 42″ E,海拔690 m),溫帶大陸性季風氣候,土壤類型為黑鈣土。年均氣溫-1.2℃,年均總降水量354.5 mm,其中75%降水量集中在6—9月,2017年總降水量為180.8 mm,與歷史總降水量相比表現(xiàn)為缺水,2018年總降水量為277.0 mm,缺水情況得到緩解,月平均氣溫與歷史相差不大(圖1)。在研究區(qū)域植被組成中,羊草和貝加爾針茅(StipabaicalensisRoshev.)為建群種,主要伴生種有斜莖黃芪(AstragalusadsurgensPall.)、多葉棘豆(Oxytropismyriophylla(Pall.) DC.)、寸草苔(CarexduriusculaC.A.Mey.)、蒲公英(TaraxacummongolicumHand.-Mazz.)等。

圖1 試驗區(qū)月均氣溫和月降水量(1951—2018年)Fig.1 Monthly mean temperature and total precipitation in the experimental area (1951—2018)
1.2 試驗設(shè)計
黃花苜蓿種子來源于內(nèi)蒙古呼倫貝爾草原生態(tài)系統(tǒng)國家野外科學觀測研究站,凈度93%,千粒重2.27 g,25℃種子萌發(fā)率88%;羊草種子來源于東北天然草地,種子萌發(fā)率12%,凈度70%,千粒重2.72 g。按照種子實際用價,播量設(shè)置為羊草64.5 kg·hm-2,黃花苜蓿30 kg·hm-2。
補播時間和補播比例按雙因素隨機區(qū)組設(shè)計,補播時間有3個水平,夏播(2017年7月1日),秋播(2017年9月30日,臨冬寄籽)和春播(2018年5月1日)。補播比例有5個水平,按種子播量設(shè)羊草和黃花苜蓿1∶1(RS1),1∶2 (RS2),1∶3(RS3)分行條播和羊草單播(MS1)黃花苜蓿單播(MS2),天然不處理草地為對照處理(CK)。同一補播時間內(nèi)設(shè)上述6個處理,4個區(qū)組。不同補播時間樣地間隔10 m,小區(qū)面積為4 m×3.5 m,小區(qū)間隔2 m,樣地總面積約3 000 m2。播種前刈割原生植被,留茬高度5 cm,播種開溝深度5~7 cm,人工播種,播種深度2~3 cm,播種行距30 cm,原位土回填并鎮(zhèn)壓。

圖2 補播小區(qū)示意圖Fig.2 The reseeding plots注:RS1,羊草∶黃花苜蓿1∶1;RS2,羊草∶黃花苜蓿1∶2;RS3,羊草∶黃花苜蓿1∶3;MS1,羊草單播;MS2,黃花苜蓿單播;CK,對照,下同Note:RS1,mixture sowing of Leymus chinensis and Medicago falcata with ratio of 1∶1;RS2,mixture sowing of Leymus chinensis and Medicago falcata with ratio of 1∶2;RS3,mixture sowing of Leymus chinensis and Medicago falcata with ratio of 1∶3;MS1,monoculture sowing of Leymus chinensis;MS2,monoculture sowing of Medicago falcata;CK,control without reseeding,the same as below
1.3 群落調(diào)查
2017年8月對夏播黃花苜蓿和羊草出苗數(shù)進行統(tǒng)計,2018年8月1日開始測定全部小區(qū)植物群落特征(高度、蓋度、生物量),每個小區(qū)隨機設(shè)置一個1 m×1 m的樣方,共72個;隨機選取同種植物5株(不足5株的全部測量),測定其自然高度。在1 m×1 m的樣方內(nèi)隨機選取一個1 m×0.25 m小樣方,將全部植物按種類齊地面刈割后裝入紙質(zhì)信封袋,105℃殺青30 min后置于65℃烘箱中于48 h后稱重。避開上述樣方測定區(qū)域,隨機放置一個由100個10 cm×10 cm小方格組成的樣方框,共72個,針刺法測定植物物種蓋度。
1.4 數(shù)據(jù)處理與分析
Shannon-Weiner指數(shù)、Simpson指數(shù)和Pielou指數(shù)用R3.4.3“vegan”包計算,重要值按功能群分為豆科、禾本科和其他科,計算公式為:重要值(Importance value,IV)=(相對蓋度+相對高度+相對生物量)/3。
所有數(shù)據(jù)通過正態(tài)分布性檢驗(Kolmogorov-Smirnov test)和方差齊性檢驗(Levene′s test)后,得出全部數(shù)據(jù)符合正態(tài)分布且方差齊性。用兩因素方差分析程序(Two-way ANOVA)分析不同補播時間和比例及其交互作用對羊草和黃花苜蓿幼苗數(shù)和植物群落(生物量、多樣性指數(shù))的影響。用單因素方差分析程序(One-way ANOVA)分析相同時間不同處理對上述指標及重要值的影響,統(tǒng)計分析軟件為SAS V8,差異顯著性檢驗水平P<0.05,SigmaPlot 12.5繪圖。利用各類群植物生物量、Simpson指數(shù)和重要值等7個指標綜合評價補播效果,采用相對值對上述指標進行標準化,通過主成分分析(Principal component analysis,PCA)計算各指標權(quán)重(Wj),最后計算綜合評價值(D),統(tǒng)計分析軟件為SPSS 19.0。計算公式如下[16]:
相對值=某一指標值/該指標對照值
隸屬函數(shù)值(uj)=(xj-xmin)/(xmax-xmin)
式中,Xj表示第j個綜合指標,j=1,2,3…n,Xmax與Xmin分別表示第j個綜合指標的最大值和最小值。
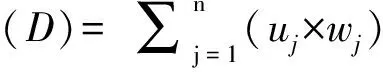
2 結(jié)果與分析
2.1 補播時間和比例對黃花苜蓿、羊草幼苗數(shù)的影響
補播時間和比例極顯著影響黃花苜蓿幼苗數(shù)(P<0.0001),兩者交互作用顯著(圖3),秋播黃花苜蓿幼苗數(shù)量顯著低于春播和夏播(P<0.05)。夏播、秋播和春播黃花苜蓿幼苗均為RS1最低,分別為99,14和86株·m-2;MS2最高,分別為212,44和258株·m-2,分別是RS1的2.1,3.2和3.0倍。夏播MS2和RS3處理顯著高于RS1和RS2處理(P<0.05);秋播除MS2顯著高于RS1外(P<0.05),與其他處理差異不顯著;春播MS2顯著高于其他處理(P<0.05)。

圖3 補播時間和比例對黃花苜蓿、羊草幼苗數(shù)量的影響Fig.3 Effects of different reseeding time and ratio on the number of seedings of Leymus chinensis (Trin.)Tzvel. and Medicago falcata L.注:數(shù)據(jù)代表平均值±標準誤。不同小寫字母表示出苗數(shù)在相同時間不同處理間差異顯著(P<0.05)。*表示不同補播時間出苗數(shù)差異顯著(P<0.05)Note:Values are shown as means±SE. Different lowercase letters showed significant differences in seedling emergence among different treatments in same reseeding time at the 0.05 level. *indicate that there were significant differences in seedling emergence between different reseeding time at the 0.05 level
補播時間和比例顯著影響羊草幼苗數(shù)(P=0.0040,P=0.0382),兩者交互作用不顯著,夏播羊草幼苗數(shù)量顯著高于春播和秋播(P<0.05)。夏播和春播羊草幼苗均為RS1最低,分別為6和4株·m-2,而秋播為RS3最低,為4株·m-2。不同補播時間羊草幼苗數(shù)均為MS1最高,分別為16,8和8株·m-2。夏播除MS1顯著高于RS1外(P<0.05),其余各處理差異不顯著。
2.2 補播對植物群落生物量的影響
補播時間顯著影響群落總生物量(P<0.05),補播比例對總生物量無顯著影響,但兩者交互作用顯著(P<0.05,圖4a),春播總生物量顯著低于夏播和秋播(P<0.05)。與CK相比,夏播各處理下總生物量均表現(xiàn)為增加趨勢,但差異不顯著;與CK相比,秋播RS3顯著增加96 g·m-2(P<0.05);而春播各處理與CK相比均表現(xiàn)為降低,其中RS1,RS2,MS2分別顯著降低了72,55和73 g·m-2(P<0.05)。
補播時間和比例對豆科和其他科植物生物量無顯著影響,兩者交互作用不顯著(圖4b,圖4 d)。補播時間顯著影響禾本科生物量(P<0.05),補播比例對禾本科總生物量無顯著影響,兩者交互作用不顯著(圖4c)。

圖4 補播對生物量的影響Fig.4 Effects of reseeding on biomass注:數(shù)據(jù)代表平均值±標準誤。不同小寫字母表示生物量相同時間不同處理間差異顯著(P<0.05)。*表示不同補播時間生物量差異顯著(P<0.05)Note:Values are shown as means±SE. Different lowercase letters showed significant differences in plant biomass among different treatments in same reseeding time at the 0.05 level. * indicate that there were significant differences in plant biomass between different reseeding time at the 0.05 level
2.3 補播對植物功能群重要值的影響
與CK相比,豆科植物重要值表現(xiàn)為增加趨勢,其中夏播RS2,RS3分別顯著增加了7%和9%(P<0.05),春播RS1,RS3和MS2分別顯著增加10%,10%和9%(P<0.05)。禾本科植物重要值與CK相比,多數(shù)處理表現(xiàn)為增加趨勢,但差異均不顯著。與豆科和禾本科不同,其他科植物重要值與CK相比則表現(xiàn)為下降趨勢(表1)。
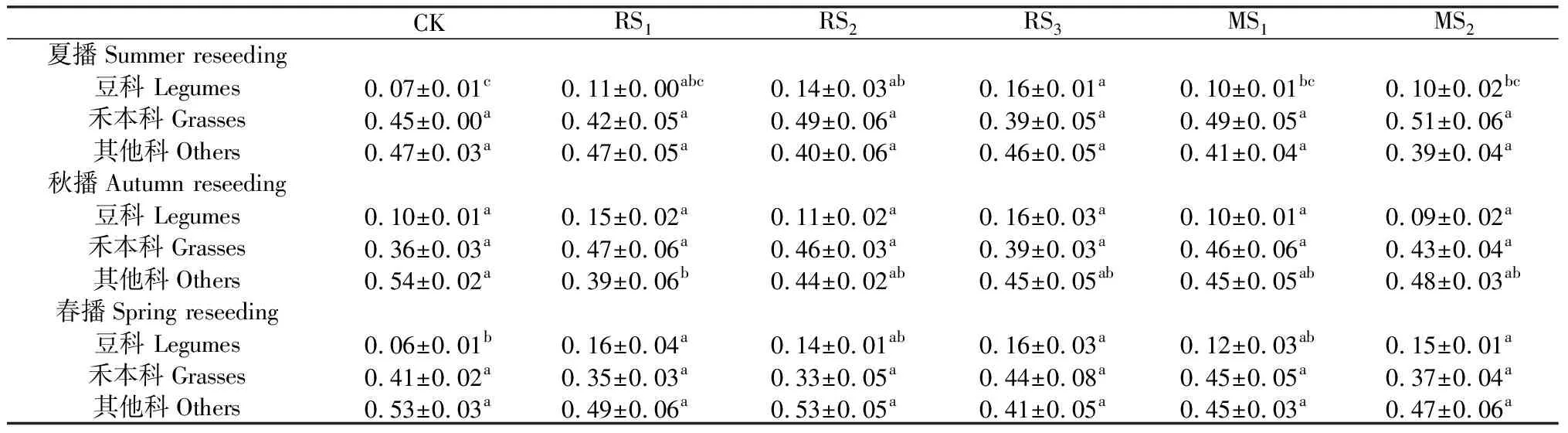
表1 補播對植物功能群重要值的影響Table 1 Effects of reseeding the important values of plant functional groups
2.4補播對植物群落多樣性的影響
如圖5所示,補播時間和比例對群落的Shannon-Weiner指數(shù)無顯著影響,兩者交互作用不顯著。補播時間對Simpson指數(shù)和Pielou指數(shù)無顯著影響,但補播比例顯著影響Simpson指數(shù)和Pielou指數(shù)(P<0.05),兩者交互作用不顯著。與CK相比,除春播RS2Shannon-Weiner指數(shù)顯著增加了0.48外(P<0.05),其余各補播處理均與CK無明顯差異,但表現(xiàn)為增加趨勢。
2.5 補播效果綜合評價
利用各類群植物生物量、Simpson指數(shù)和重要值等7個指標綜合評價補播效果,不同補播時間綜合效果表現(xiàn)為:夏播(D=0.51)>春播(D=0.46)>秋播(D=0.45),不同補播比例平均值綜合表現(xiàn)為:RS3(D=0.560)>RS2(D=0.473)>RS1(D=0.456)>MS2(D=0.453)>MS1(D=0.436,表2),由此可見,夏播RS3補播效果較好。

表2 不同補播時間和比例隸屬函數(shù)值、權(quán)重、D值及排序Table 2 Membership function value,weight,D Value and order of different reseeding time and ratio
3 討論
3.1 補播時間和比例對羊草、黃花苜蓿建植的影響
水分和溫度直接影響植物種子萌發(fā)和生長速率,是我國北方草原區(qū)的主要限制因素,決定補播能否成功[17],秋播羊草和黃花苜蓿種子在早春萌發(fā),本研究區(qū)早春氣溫波動較大,容易出現(xiàn)“倒春寒”,威脅幼苗生存。此外,即使補播幼苗能正常發(fā)芽,原生植被中返青早的闊葉類植物易對幼苗造成“遮蔭效應(yīng)”,有研究表明植物鮮重、根鮮重和分株數(shù)會隨遮光度增大表現(xiàn)為逐漸減小趨勢[18]。秋播幼苗對光競爭處于劣勢,致使秋播植株矮小、黃化,生長發(fā)育受限,Culleton[19]認為,秋播可能由于苗床準備不足、土壤肥力低下及昆蟲襲擊等原因而導致失敗,而春播和夏播前的刈割處理降低了原生植被與補播幼苗的資源競爭,為幼苗存活提供了有利條件,因此秋季補播效果差。
本研究發(fā)現(xiàn)春播和秋播黃花苜蓿建植率均為單播處理最高,而夏播則較低,可能是2017年夏季干旱所致,高密度播種導致黃花苜蓿種內(nèi)對水資源競爭激烈,而低密度下競爭相對較小,幼苗更易存活。此外,研究區(qū)2017年總降水量較低導致蝗蟲(LocustamigratoriamanilensisMeyen)大量繁殖,黃花苜蓿幼苗生長時的“獨特香味”更易誘導昆蟲襲擊,致其出現(xiàn)損傷甚至死亡。但土壤含水量低可能利于羊草種子萌發(fā),馬紅媛等[20]、楊焜等[21]發(fā)現(xiàn)羊草種子在土壤含水量較低條件下更易萌發(fā),且羊草種子萌發(fā)率隨土壤含水量上升而表現(xiàn)為下降趨勢。此外,楊偉光等[22]發(fā)現(xiàn),羊草種子在變溫條件下萌發(fā)率較高,夏播處理部分羊草種子萌發(fā),其余種子在經(jīng)過嚴寒后打破種子休眠在第二年萌發(fā),因此,夏播羊草幼苗數(shù)較高。本研究發(fā)現(xiàn),羊草建植率在播種密度低時較高,這與羊草繁殖策略有關(guān),羊草低密度播種有利于其種群數(shù)量增加,使群落中羊草未來種群數(shù)量維持在適宜水平[23]。
3.2 補播對植物群落特征影響
本研究發(fā)現(xiàn)多數(shù)補播比例處理下的總生物量與CK無顯著差異,這是由于新補播的草種還未完全定居,僅表現(xiàn)為補充退化草地空缺生態(tài)位,不同補播比例對群落生物量的影響可能會隨補播年限的增加有所變化[24]。補播后草地生態(tài)系統(tǒng)植物群落總生物量的變化主要由禾本科植物生物量決定[25],本研究發(fā)現(xiàn)夏播禾本科植物生物量最高,這是由于補播羊草種子在該時期建植較好,羊草幼苗數(shù)增加。另一方面,天然草甸易形成土壤通透性低和水汽含量小的緊實土壤表層,導致羊草無性繁殖受阻[26],補播能打破致密草層提高土壤通透性,刺激羊草無性繁殖,夏播恢復時間相對于秋播和春播最長,劃破草皮的“效應(yīng)”也最長,而春播由于恢復時間短,原生植被未完全恢復,補播效應(yīng)也沒有體現(xiàn)出來[27],導致春播各補播比例總生物量均低于CK,因此有待于進一步觀察。楊增增等[28]在退化高寒草地補播試驗表明,群落中禾本科植物重要值由0.40上升到0.51,其他科植物重要值則從0.66下降到0.49,這與本研究發(fā)現(xiàn)一致,補播后羊草和黃花苜蓿迅速占領(lǐng)退化草地空缺生態(tài)位,促使植物群落競爭機制和競爭格局發(fā)生改變[29],導致其他科植物生長和繁殖受限,重要值下降。
補播能降低退化草地生態(tài)系統(tǒng)中種子缺乏的限制,使植物群落豐富度增加,打破原有群落競爭格局,而補播前劃破草皮改變了原有植物空間格局,抑制雜類草生長,二者共同影響植物群落多樣性[30-32]。本研究發(fā)現(xiàn)補播比例顯著影響Simpson指數(shù)和Pielou指數(shù),可能是不同補播比例間各植物種內(nèi)競爭和種間競爭大小不同而導致,但同一補播時間多數(shù)處理間群落多樣性指數(shù)差異不顯著,這是由于補播雖然改變了原有群落競爭格局,但由于補播恢復時間短,群落處于不穩(wěn)定狀態(tài),補播物種與原有植被之間需要通過長時間物種競爭重新調(diào)整生長格局[26],有研究表明補播8年后退化草地植物群落能達到較好的恢復效果,18年以后才能達到未退化效果[33],因此,補播對多樣性指數(shù)的影響也需要長時間深入觀察。
4 結(jié)論
本研究以羊草和黃花苜蓿為補播對象,設(shè)置不同補播時間和不同比例,對呼倫貝爾退化草甸草原進行人工恢復,研究結(jié)果顯示該地區(qū)夏播羊草和黃花苜蓿幼苗數(shù)量顯著高于秋播和春播,夏播和秋播植物群落總生物量呈增加趨勢,而春播呈下降趨勢。無論是從補播時間還是從補播比例來看,人工補播措施能提高退化草地植物群落中豆科和禾本科植物的重要值,降低其他科植物重要值,植物群落多樣性指數(shù)呈上升趨勢,但需要長時間深入觀察。從綜合效果來看,夏季補播羊草和黃花苜蓿比例為1∶3綜合得分最高,可為該地區(qū)退化草地恢復提供參考。
