紅螯螯蝦卵黃蛋白原VWD結(jié)構(gòu)域的原核表達(dá)及純化
2021-09-08 01:19:19文露婷許藝蘭童桂香黃黎明楊彥豪杜雪松黃光華韋信賢王瑞楊慧贊
南方農(nóng)業(yè)學(xué)報(bào) 2021年5期
文露婷 許藝蘭 童桂香 黃黎明 楊彥豪 杜雪松 黃光華 韋信賢 王瑞 楊慧贊

摘要:【目的】原核表達(dá)紅螯螯蝦(Cherax quadricarinatus)卵黃蛋白原(Vg)VWD結(jié)構(gòu)域,為深入開展Vg的生物學(xué)功能研究和開發(fā)相應(yīng)的生物活性物質(zhì)提供技術(shù)支持,也為改進(jìn)蝦類養(yǎng)殖催熟及獲取高質(zhì)量后代打下基礎(chǔ)。【方法】在GenBank中搜索紅螯螯蝦卵Vg的mRNA序列(AF306784.1)及查找其VWD結(jié)構(gòu)域(2345~2491 aa),采用ExPASy ProtParam、ProtScale、InterProscan及TMHMM等在線軟件進(jìn)行生物信息學(xué)分析及理論評(píng)估,結(jié)合大腸桿菌密碼子偏好優(yōu)化原始VWD氨基酸序列;然后通過(guò)全基因合成獲得目的基因,連接至載體pET-28a構(gòu)建重組質(zhì)粒pET-28a-VWD,挑取陽(yáng)性克隆轉(zhuǎn)化大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞及采用IPTG進(jìn)行誘導(dǎo)表達(dá),并以10% SDS-PAGE電泳和Wes-tern blotting檢測(cè)誘導(dǎo)表達(dá)的融合蛋白。【結(jié)果】紅螯螯蝦VWD結(jié)構(gòu)域相對(duì)分子量為19692.11 Da,理論等電點(diǎn)(pI)為9.35,分子式為C855H1334N258O261S9,屬于不穩(wěn)定的親水性蛋白,不含跨膜結(jié)構(gòu)域。延伸鏈和無(wú)規(guī)則卷曲是紅螯螯蝦VWD結(jié)構(gòu)域二級(jí)結(jié)構(gòu)的主要元件,其中,α-螺旋占7.73%,β-轉(zhuǎn)角占10.50%,延伸鏈占35.91%,無(wú)規(guī)則卷曲占45.86%。重組質(zhì)粒pET-28a-VWD經(jīng)大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞誘導(dǎo)表達(dá)即獲得融合蛋白VWD,以2 mol/L鹽酸胍溶解和Ni-NTA親和層析純化及復(fù)性后,10% SDS-PAGE電泳和Western blotting檢測(cè)均在蛋白相對(duì)分子量約20.0 kD處出現(xiàn)1條清晰的特異性條帶,融合蛋白VWD表達(dá)形式以包涵體為主。融合蛋白VWD在BL21(λDE3)感受態(tài)細(xì)胞中高效表達(dá)的最佳誘導(dǎo)條件:當(dāng)單克隆菌液OD600 nm達(dá)0.6時(shí)添加0.5 mmol/L IPTG,置于20 ℃下誘導(dǎo)培養(yǎng)16 h。純化后的融合蛋白VWD濃度為1.32 mg/mL。【結(jié)論】通過(guò)全基因合成獲得的優(yōu)化紅螯螯蝦VWD基因能在大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞中誘導(dǎo)表達(dá)出以包涵體為主要形式的融合蛋白,經(jīng)鹽酸胍溶解和Ni-NTA親和層析柱純化及復(fù)性即可獲得高純度的活性VWD蛋白,為后續(xù)開展紅螯螯蝦Vg生物學(xué)功能研究及開發(fā)相應(yīng)的生物活性物質(zhì)提供技術(shù)支持。
關(guān)鍵詞: 紅螯螯蝦;卵黃蛋白原(Vg);VWD結(jié)構(gòu)域;全基因合成;原核表達(dá);包涵體
中圖分類號(hào): S966.12? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻(xiàn)標(biāo)志碼: A 文章編號(hào):2095-1191(2021)05-1378-09
Abstract:【Objective】Prokaryotic expression of the vitellogenin(Vg) VWD domain of red claw crayfish(Cherax quadricarinatus) not only provided technical support for further research on the biological function of Vg and the development of corresponding bioactive substances, but also laid? foundation for improving C. quadricarinatus culture, accelera-ting ripening and obtaining high-quality offspring. 【Method】The protein sequence of VWD domain(2345-2491 aa) was found according to the Vg mRNA sequence(accession number:AF306784.1) of C. quadricarinatus published in GenBank database. Bioinformatics analysis and theoretical evaluation were carried out by using online softwares such as ExPASyProtParam, ProtScale, InterProscan and TMHMM. Optimization of original VWD amino acid sequence combined with codon preference of Escherichia coli. Then the target gene was obtained by whole gene synthesis,and the recombinant plasmid E. coli pET-28a-VWD was constructed by connecting to vector pET-28a,and transformed into positive clone conversion BL21(λDE3) competent cells which were induced by IPTG. The induced fusion protein was detected by 10% SDS-PAGE electrophoresis and Western blotting.【Result】The relative molecular weight of VWD domain of C. quadricarinatus was 19692.11 Da, theoretical isoelectric points(pI)was 9.35, and the molecular formula was C855H1334N258O261S9. It was an unstable hydrophilic protein and did not contain a transmembrane domain. Extended chain and irregular curl were the main elements of the secondary structure of VWD domain in C. quadricarinatus, among which, α-helix accounted for 7.73%, β-turn accounted for 10.50%, the extended chain accounted for 35.91%, and the irregular curl accounted for 45.86%. The recombinant plasmid pET-28a-VWD expressed the fusion protein VWD by inducing the competent cells E. coli BL21(λDE3). After dissolution with 2 mol/L guanidine hydrochloride and purification and renaturation on Ni-NTA affinity chromatography column, a clear specific band appeared at the relative molecular weight of the protein about 20.0 kD by 10% SDS-PAGE electrophoresis and Western blotting. The expression form of fusion protein VWD was mainly inclusion body. The best induction conditions for high expression of the fusion protein VWD in the competent cells BL21 (λDE3): when the OD600 nm of the monoclonal bacterial solution reached 0.6, 0.5 mmol/L IPTG was added and cultured at 20 ℃ for 16 h. The concentration of purified fusion protein VWD was 1.32 mg/mL. 【Conclusion】The optimized VWD gene of C. quadricarinatus obtained by whole gene synthesis can be expressed in E. coli competent cells BL21(λDE3),and the expression form of fusion protein VWD is mainly inclusion body. After dissolving with guanidine hydrochloride and purifying and renaturating with Ni-NTA affinity chromatography column, high-purity active VWD protein can be obtained, which provides technical support for the subsequent research on the biological function of C. quadricarinatus Vg and the development of corresponding bioactive substances.
Key words: Cherax quadricarinatus; vitellogenin(Vg); VWD domain; whole gene synthesis; prokaryotic expre-ssion;inclusion body
Foundation item: Guangxi Key Science and Technology Project(Guike AA17204095-5,Guike AA17204094-7); Guangxi Natural Science Foundation(2019GXNSFAA185036);Guangxi Key Research and Development Project(Guike AB182 21068)
0 引言
【研究意義】卵黃蛋白原(Vitellogenin,Vg)是卵黃蛋白(Yolk protein)的前體,是由若干個(gè)基因編碼的特異性高分子量蛋白,富含糖、磷、脂,存在于卵生和非哺乳類性成熟的動(dòng)物血液中(Avarre et al.,2007;王加偉等,2016),可為動(dòng)物胚胎發(fā)育提供和轉(zhuǎn)運(yùn)氨基酸、脂肪、維生素、硫及鈣等營(yíng)養(yǎng)物質(zhì)。除了已知的營(yíng)養(yǎng)和運(yùn)輸功能外,Vg在進(jìn)入卵細(xì)胞的過(guò)程中還發(fā)揮著其他非營(yíng)養(yǎng)性功能(盧建平和姜乃澄,2000;潘杰,2020)。在褐飛虱和煙粉虱等蟲體內(nèi),Vg可被病原微生物所利用,實(shí)現(xiàn)入卵傳播(Cheng and Hou,2005;Wei et al.,2017);在蚊子體內(nèi),Vg通過(guò)調(diào)節(jié)含硫酯鍵蛋白(Thioester-containing protein 1)表達(dá),而實(shí)現(xiàn)對(duì)瘧原蟲免疫清除的調(diào)控(Rono and Ceccarelli,2010);在蜜蜂血淋巴中,Vg通過(guò)與病原微生物細(xì)胞壁的脂磷壁酸或脂多糖互作,介導(dǎo)昆蟲對(duì)病原微生物的免疫清除等(Salmela et al.,2015)。此外,Vg可作為一種雌激素和類雌激素的標(biāo)志物,應(yīng)用于水環(huán)境中內(nèi)分泌干擾物、環(huán)境毒理與污染等狀況的調(diào)查研究(周慶祥和江桂斌,2003)。Vg具有豐富的生物功能和極高的研究?jī)r(jià)值,因此開展Vg外源表達(dá)及純化研究,對(duì)進(jìn)一步挖掘其生物學(xué)功能及綜合利用開發(fā)具有重要意義。【前人研究進(jìn)展】目前,關(guān)于Vg的研究主要集中在生物學(xué)功能(Li et al.,2009;Garcia et al.,2010)、分子結(jié)構(gòu)特征(Dalvin et al.,2011;Sun et al.,2013)及表達(dá)調(diào)控機(jī)制(Lu et al.,2016;Luo et al.,2017)等方面。大量研究表明,無(wú)論是在無(wú)脊椎動(dòng)物還是在脊椎動(dòng)物中,Vg的分子結(jié)構(gòu)及其功能均具有一定的相似性,同時(shí)又具有各自的獨(dú)特性。Vg一般含有3個(gè)非常保守的結(jié)構(gòu)域,分別是卵黃蛋白原N氨基端的結(jié)構(gòu)域(VitN)、羧基端的血管性血友病因子結(jié)構(gòu)域(VWD)及未知功能的結(jié)構(gòu)域(DUF)(Tufail and Takeda,2008)。VitN結(jié)構(gòu)域是Vg最主要的磷酸化位點(diǎn),也是重要的蛋白修飾區(qū)域,包含Vg與卵黃蛋白原受體(VgRs)的互作位點(diǎn),在Vg的剪切、Vg-VgRs識(shí)別及其所介導(dǎo)的營(yíng)養(yǎng)運(yùn)輸中發(fā)揮重要作用;VWD結(jié)構(gòu)域和DUF結(jié)構(gòu)域則具有與病毒或細(xì)菌等病原微生物互作的功能,能對(duì)病原微生物進(jìn)行識(shí)別清除,并介導(dǎo)其垂直傳播(霍巖等,2018)。已有研究發(fā)現(xiàn),通過(guò)干擾果蠅Vg與VgRs結(jié)合,能影響其體內(nèi)逆轉(zhuǎn)錄病毒進(jìn)入卵細(xì)胞(Leblanc et al.,2000;Brasset et al.,2006)。Li等(2009)研究發(fā)現(xiàn)魚類Vg對(duì)細(xì)菌的細(xì)胞壁脂多糖和磷脂壁酸具有識(shí)別功能,能促使細(xì)菌裂解。Huo等(2014)研究表明,雄性灰飛虱體內(nèi)Vg的DUF結(jié)構(gòu)域和VWD結(jié)構(gòu)域可與水稻條紋病毒(Rice stripe virus,RSV)結(jié)構(gòu)蛋白結(jié)合,再通過(guò)與營(yíng)養(yǎng)細(xì)胞表面的VgRs結(jié)合而進(jìn)入卵母細(xì)胞內(nèi),達(dá)到垂直傳播的目的。針對(duì)紅螯螯蝦(Cherax quadricarinatus)Vg的相關(guān)研究,Abdu等(2002)通過(guò)克隆及測(cè)序分析紅螯螯蝦Vg基因全長(zhǎng)cDNA序列,證實(shí)其編碼2584個(gè)氨基酸殘基,與其他甲殼類動(dòng)物的Vg序列相似性在65%左右,靠近基因3'端的區(qū)域編碼一種帶負(fù)電荷且具有結(jié)合鈣能力的蛋白質(zhì)(P106);Shechter等(2005)通過(guò)摘除雄性紅螯螯蝦的竇腺?gòu)?fù)合體(XO-SG)誘導(dǎo)Vg轉(zhuǎn)錄、翻譯和釋放,發(fā)現(xiàn)該過(guò)程與雌性個(gè)體的卵黃發(fā)生過(guò)程相似,Vg在紅螯螯蝦蛻皮間期未發(fā)生表達(dá),但在脫殼后早期高表達(dá)。可見,明確Vg在內(nèi)分泌調(diào)控蛻皮甲殼類動(dòng)物中的表達(dá)情況,可為研究甲殼類動(dòng)物Vg表達(dá)和翻譯的內(nèi)分泌調(diào)節(jié)機(jī)制及揭示蛻皮與生殖內(nèi)分泌軸間的關(guān)系提供誘導(dǎo)模型。【本研究切入點(diǎn)】至今,針對(duì)甲殼類動(dòng)物Vg的研究主要集中在Vg表達(dá)調(diào)控、生物學(xué)功能及其機(jī)制方面,有關(guān)Vg單個(gè)結(jié)構(gòu)域具體功能及其作用機(jī)制的研究鮮見報(bào)道,尤其是VWD結(jié)構(gòu)域。Vg各結(jié)構(gòu)域的分子結(jié)構(gòu)及其生物學(xué)功能存在明顯差異,因此有必要對(duì)各結(jié)構(gòu)域進(jìn)行單獨(dú)研究,以探明其具體的作用機(jī)制。【擬解決的關(guān)鍵問(wèn)題】對(duì)紅螯螯蝦Vg的VWD結(jié)構(gòu)域(2345~2491 aa)進(jìn)行生物信息學(xué)分析及理論評(píng)估,設(shè)計(jì)并優(yōu)化其基因序列,采用全基因合成獲得優(yōu)化基因序列,并通過(guò)原核表達(dá)載體誘導(dǎo)表達(dá)VWD蛋白,為深入開展Vg的生物學(xué)功能研究和開發(fā)相應(yīng)的生物活性物質(zhì)提供技術(shù)支持,也為改進(jìn)蝦類養(yǎng)殖催熟及獲取高質(zhì)量后代打下基礎(chǔ)。
1 材料與方法
1. 1 試驗(yàn)材料
大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞由廣西水產(chǎn)遺傳育種與健康養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室保存提供。載體pET-28a、瓊脂糖凝膠回收試劑盒、質(zhì)粒提取試劑盒、Ni-NTA純化樹脂預(yù)裝柱非預(yù)染蛋白Marker、預(yù)染蛋白Marker、蛋白濃度定量試劑盒、TMB顯色試劑盒及IPTG等購(gòu)自生工生物工程(上海)股份有限公司;兔抗His-Tag抗體和辣根過(guò)氧化物酶(HRP)標(biāo)記羊抗兔IgG抗體購(gòu)自德國(guó)Merck公司;Pfu高溫聚合酶、限制性內(nèi)切酶及T4 DNA連接酶購(gòu)自美國(guó)Promega公司;其他試劑均為國(guó)產(chǎn)分析純。
1. 2 VWD序列生物信息學(xué)分析
登錄GenBank搜索紅螯螯蝦Vg的mRNA序列(AF306784.1),導(dǎo)出其VWD結(jié)構(gòu)域(2345~2491 aa),利用NCBI ORFFinder(https://www.ncbi.nlm.nih.gov/orffinder)查找VWD基因的開放閱讀框(ORF)并推導(dǎo)其氨基酸序列。將推導(dǎo)的氨基酸序列提交至ExPASy數(shù)據(jù)庫(kù)(https://www.expasy.org/),采用ProtParam、ProtScale、InterProscan和TMHMM(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)等在線分析軟件分別進(jìn)行親/疏水性、信號(hào)肽、跨膜結(jié)構(gòu)域及基本結(jié)構(gòu)等生物學(xué)信息進(jìn)行分析和理論評(píng)估。
1. 3 序列優(yōu)化與全基因合成
根據(jù)VWD氨基酸序列的評(píng)估結(jié)果及大腸桿菌密碼子偏好性,通過(guò)OPTIMIZER(http://genomes.urv.es/OPTIMIZER/)和DNAWORKS(https://hpcwebapps.cit.nih.gov/dnaworks/)對(duì)原始VWD氨基酸序列進(jìn)行密碼子優(yōu)化及序列改造,設(shè)計(jì)優(yōu)化目的片段的全基因合成方案:將優(yōu)化后目的片段分割成18段帶有重疊序列的寡核苷酸片段(F:5'-GACACGGATCCCT TACAACCGT-3';R:5'-GTGTCCTCGAGTTAGCCG GACGGAG-3',下劃線部分分別為BamH I和Xho I酶切位點(diǎn)),委托生工生物工程(上海)股份有限公司合成。全基因合成策略是將合成的18段寡核苷酸片段混合在一個(gè)體系中,通過(guò)第一輪PCR擴(kuò)增的多個(gè)熱循環(huán)和退火實(shí)現(xiàn)目的片段的全長(zhǎng)序列拼接,再利用首尾引物通過(guò)第二輪PCR進(jìn)一步擴(kuò)增和富集全長(zhǎng)目的基因。第一輪PCR反應(yīng)體系50.0 ?L:18對(duì)引物Mix各0.5 ?L,dNTP(25 mmol/L)1.0 ?L,10×Pfu Buffer 5.0 ?L,Pfu高溫聚合酶(5 U/?L)0.4 ?L,ddH2O補(bǔ)足至50.0 ?L。擴(kuò)增程序:95 ℃預(yù)變性3 min;95 ℃ 22 s,55 ℃ 20 s,72 ℃ 30 s,進(jìn)行20個(gè)循環(huán);72 ℃延伸5 min。以第一輪PCR產(chǎn)物為模板進(jìn)行第二輪PCR擴(kuò)增,反應(yīng)體系50.0 ?L:首尾引物各2.0 ?L(引物濃度同上),第一輪PCR產(chǎn)物(模板),dNTP(25 mmol/L)1.0 ?L,10×Pfu Buffer 5.0 ?L,Pfu高溫聚合酶(5 U/?L)0.4 ?L,ddH2O補(bǔ)足至50.0 ?L。擴(kuò)增程序:95 ℃預(yù)變性3 min;95 ℃ 22 s,55 ℃ 20 s,72 ℃ 30 s,進(jìn)行22個(gè)循環(huán);72 ℃延伸5 min。第二輪PCR擴(kuò)增產(chǎn)物采用1.2%瓊脂糖凝膠電泳進(jìn)行檢測(cè),并對(duì)特異性條帶進(jìn)行純化回收。
1. 4 重組表達(dá)載體構(gòu)建與驗(yàn)證
根據(jù)引物設(shè)計(jì)的酶切位點(diǎn),利用BamH I和Xho I對(duì)純化回收得到的目的基因和載體pET-28a進(jìn)行雙酶切。目的基因酶切體系50.0 ?L:純化回收的目的基因20.0 ?L(1.0 ?g),10×FD Buffer 5.0 ?L,BamH I和Xho I各1.0 ?L(10 U/?L),ddH2O 23.0 ?L。載體pET-28a酶切體系50.0 ?L:pET-28a 1.0 ?g,10×FD Buffer 5.0 ?L,BamH I和Xho I各1.0 ?L(10 U/?L),ddH2O補(bǔ)足至50.0 ?L。將上述2個(gè)酶切體系分別放入37 ℃恒溫水浴鍋中反應(yīng)2 h。將酶切后純化回收的目的DNA片段和pET-28a進(jìn)行連接,連接體系20.0 ?L:目的片段酶切產(chǎn)物8.0 ?L,pET-28a酶切產(chǎn)物4.0 ?L,10×T4 DNA Ligase Buffer 2.0 ?L,T4 DNAigase 1.0 ?L(5 U/?L), ddH2O補(bǔ)足至20.0 ?L。將上述連接混合液放入22 ℃的PCR儀內(nèi)反應(yīng)1 h,然后轉(zhuǎn)化BL21(λDE3)感受態(tài)細(xì)胞,均勻涂布于含卡拉霉素的LB培養(yǎng)基上,37 ℃培養(yǎng)過(guò)夜,篩選陽(yáng)性克隆,提取質(zhì)粒,進(jìn)行酶切驗(yàn)證,反應(yīng)體系10.0 ?L:重組質(zhì)粒100 ng,10×FD Buffer 1.0 ?L,BamH I和Xho I各1.0 ?L(10 U/?L),ddH2O補(bǔ)足至10.0 ?L,置于37 ℃恒溫水浴鍋中消化2 h。經(jīng)1.2%瓊脂糖凝膠電泳檢測(cè),將陽(yáng)性克隆接種至含卡拉霉素的LB肉湯培養(yǎng)基中,250 r/min搖床培養(yǎng)過(guò)夜,菌液送至北京華大基因科技有限公司測(cè)序。測(cè)序引物為載體pET-28a的T7啟動(dòng)子和終止子通用引物(5'端測(cè)序引物5'-TAATACGACT CACTATAGGG-3',3'端測(cè)序引物5'-GCTAGTTATT GCTCAGCGGG-3')。
1. 5 VWD蛋白原核表達(dá)
成功構(gòu)建的重組質(zhì)粒命名為pET-28a-VWD,挑取陽(yáng)性克隆轉(zhuǎn)化BL21(λDE3)感受態(tài)細(xì)胞,熱激后涂布于含卡拉霉素的LB培養(yǎng)基上,37 ℃培養(yǎng)過(guò)夜;挑取單克隆菌落接種至含卡拉霉素的LB液體培養(yǎng)基中進(jìn)行37 ℃活化培養(yǎng),當(dāng)菌液OD600 nm達(dá)0.6時(shí),添加一定濃度的IPTG,分別于16和20 ℃下培養(yǎng)16 h,再置于37 ℃下培養(yǎng)4 h,設(shè)未添加IPTG的為陰性對(duì)照。取菌液12000 r/min離心5 min,棄上清液,收集菌體沉淀,加入PBS(pH 7.4)進(jìn)行懸浮,利用超聲波破碎儀使其充分溶解,離心后棄上清液,向沉淀中加入緩沖液A(8 mol/L Urea,50 mmol/L Tris-HCl,300 mmol/L NaCl,pH 8.0)進(jìn)行溶解,再次離心,向上清液和沉淀中分別加入等體積的2×Loading Buffer,煮沸后進(jìn)行10% SDS-PAGE電泳,通過(guò)考馬斯亮藍(lán)(R250)染色確定蛋白表達(dá)量及純度,進(jìn)而確定誘導(dǎo)表達(dá)所需的最佳IPTG濃度、誘導(dǎo)溫度及誘導(dǎo)時(shí)間。最后于最佳誘導(dǎo)條件下進(jìn)行大量表達(dá),菌液在4 ℃下3000 r/min離心10 min后,收集菌體沉淀。
1. 6 融合蛋白純化
向收集到的菌體中加入緩沖液B(8 mol/L Urea,50 mmol/L Tris,300 mmol/L NaCl,0.1% Triton X-100,pH 8.0),4 ℃靜置30 min后置于冰上進(jìn)行200 W的多次間歇性超聲波破碎,待菌液澄清后,4 ℃下12000 r/min離心10 min,收集上清液粗蛋白(包涵體),加入2 mol/L鹽酸胍進(jìn)行溶解。取5 mL的Ni-NTA親和層析柱,以5倍柱床體積的Binding Buffer清洗平衡柱,流速5 mL/min;將粗蛋白與平衡后的Ni-NTA親和層析柱填料孵育1 h;孵育后的產(chǎn)物上柱進(jìn)行提純和復(fù)性,收集流出液;先用Binding Buffer清洗平衡柱,接著以Washing Buffer洗柱,收集流出液;用Elution Buffer再次洗脫,收集流出液。分別對(duì)粗蛋白、洗雜流出液及洗脫流出液進(jìn)行處理,經(jīng)10% SDS-PAGE電泳后進(jìn)行R250染色分析。將純度較高的6個(gè)組分分別加入到緩沖液C(50 mmol/L Tris,300 mmol/L NaCl,0.1% SKL,pH 8.5)中進(jìn)行透析,透析結(jié)束采用PEG-20000進(jìn)行濃縮,0.45 μm濾膜過(guò)濾后分裝為1 mL/管,-80 ℃保存。
1. 7 融合蛋白表達(dá)驗(yàn)證
對(duì)復(fù)性且純化后的融合蛋白進(jìn)行制樣,經(jīng)10% SDS-PAGE電泳后,在200 mA恒定電流及低溫條件下轉(zhuǎn)膜(PVDF)2 h,再以含5%脫脂奶粉的TBST封閉2 h,將兔抗His-Tag抗體進(jìn)行1∶2000稀釋,HRP標(biāo)記羊抗兔IgG抗體進(jìn)行1∶5000稀釋,常溫孵育過(guò)夜,洗膜3次,將TMB顯色液滴加到PVDF膜上顯色,進(jìn)行Western blotting特異性驗(yàn)證。
2 結(jié)果與分析
2. 1 紅螯螯蝦VWD結(jié)構(gòu)域生物信息學(xué)分析結(jié)果
2. 1. 1 紅螯螯蝦VWD結(jié)構(gòu)域的理化性質(zhì) 紅螯螯蝦VWD結(jié)構(gòu)域相對(duì)分子量為19692.11 Da,理論等電點(diǎn)(pI)為9.35,負(fù)電荷殘基(Asp+Glu)總數(shù)為12個(gè),正電荷殘基(Arg+Lys)總數(shù)為15個(gè),共計(jì)2717個(gè)原子,分子式為C855H1334N258O261S9,不穩(wěn)定系數(shù)為40.74,大于40,即為不穩(wěn)定蛋白;其脂肪指數(shù)為63.98,親水性平均系數(shù)(GRAVY)為-0.367,ProtScale預(yù)測(cè)結(jié)果(圖1)也表明該蛋白的親水氨基酸數(shù)量分布明顯多于疏水氨基酸,故推測(cè)為親水性蛋白。經(jīng)TMHMM預(yù)測(cè)顯示,編碼VWD結(jié)構(gòu)域的181個(gè)氨基酸殘基全部位于細(xì)胞膜外,即不含跨膜結(jié)構(gòu)域(圖2)。
2. 1. 2 紅螯螯蝦VWD結(jié)構(gòu)域的結(jié)構(gòu)特征 利用SOPMA對(duì)紅螯螯蝦VWD結(jié)構(gòu)域二級(jí)結(jié)構(gòu)進(jìn)行預(yù)測(cè),結(jié)果如圖3所示。紅螯螯蝦VWD結(jié)構(gòu)域二級(jí)結(jié)構(gòu)由α-螺旋(Alpha helix)、延伸鏈(Extended strand)、β-轉(zhuǎn)角(Beta turn)及無(wú)規(guī)則卷曲(Random coil)4種類型構(gòu)成,其中,α-螺旋有14個(gè)(占7.73%),β-轉(zhuǎn)角有19個(gè)(占10.50%),延伸鏈有65條(占35.91%),無(wú)規(guī)則卷曲有83個(gè)(占45.86%),即延伸鏈和無(wú)規(guī)則卷曲是VWD結(jié)構(gòu)域二級(jí)結(jié)構(gòu)的主要元件。α-螺旋聚集在第20~40位氨基酸間,β-轉(zhuǎn)角則零星散布于整個(gè)氨基酸序列中。
紅螯螯蝦VWD結(jié)構(gòu)域三級(jí)結(jié)構(gòu)是以c6rbfA蛋白結(jié)構(gòu)為基礎(chǔ)進(jìn)行同源建模,其中對(duì)齊殘基數(shù)為137個(gè),覆蓋率為76%,一致性為17%,預(yù)測(cè)可信度為100%。紅螯螯蝦VWD結(jié)構(gòu)域三級(jí)結(jié)構(gòu)(圖4)由上下兩層Sheet片段通過(guò)長(zhǎng)鏈卷曲相連接而成,其下層還包含一段螺旋結(jié)構(gòu)。
2. 2 重組質(zhì)粒pET-28a-VWD驗(yàn)證結(jié)果
2. 2. 1 全基因合成驗(yàn)證結(jié)果 通過(guò)全基因合成獲得的PCR產(chǎn)物經(jīng)1.2%瓊脂糖凝膠電泳檢測(cè),發(fā)現(xiàn)在450 bp附近出現(xiàn)單一、明亮的特異性條帶(圖5),與預(yù)期設(shè)計(jì)的目的基因大小相符,說(shuō)明已成功合成獲得紅螯螯蝦VWD基因。
2. 2. 2 重組質(zhì)粒pET-28a-VWD鑒定結(jié)果? 提取重組質(zhì)粒pET-28a-VWD,經(jīng)BamH I和Xho I雙酶切后進(jìn)行1.2%瓊脂糖凝膠電泳檢測(cè),結(jié)果(圖6)顯示,酶切產(chǎn)物為2條大小約5300和450 bp的片段,與預(yù)期結(jié)果相符。對(duì)篩選出的陽(yáng)性克隆(重組質(zhì)粒pET-28a-VWD)進(jìn)行增菌培養(yǎng),菌液送至北京華大基因科技有限公司測(cè)序,其測(cè)序結(jié)果與NCBI(https://www.ncbi.nlm.nih.gov/)已公布紅螯螯蝦Vg基因的核苷酸序列(AAG17936.1)相似性為100.0%,而推導(dǎo)氨基酸序列與紅螯螯蝦Vg氨基酸序列(AAS68191.1)的相似性為75%,說(shuō)明通過(guò)全基因合成并克隆轉(zhuǎn)化獲得的氨基酸序列即為紅螯螯蝦Vg的VWD結(jié)構(gòu)域,同時(shí)確認(rèn)重組質(zhì)粒pET-28a-VWD構(gòu)建成功。
2. 3 融合蛋白VWD表達(dá)形式鑒定結(jié)果
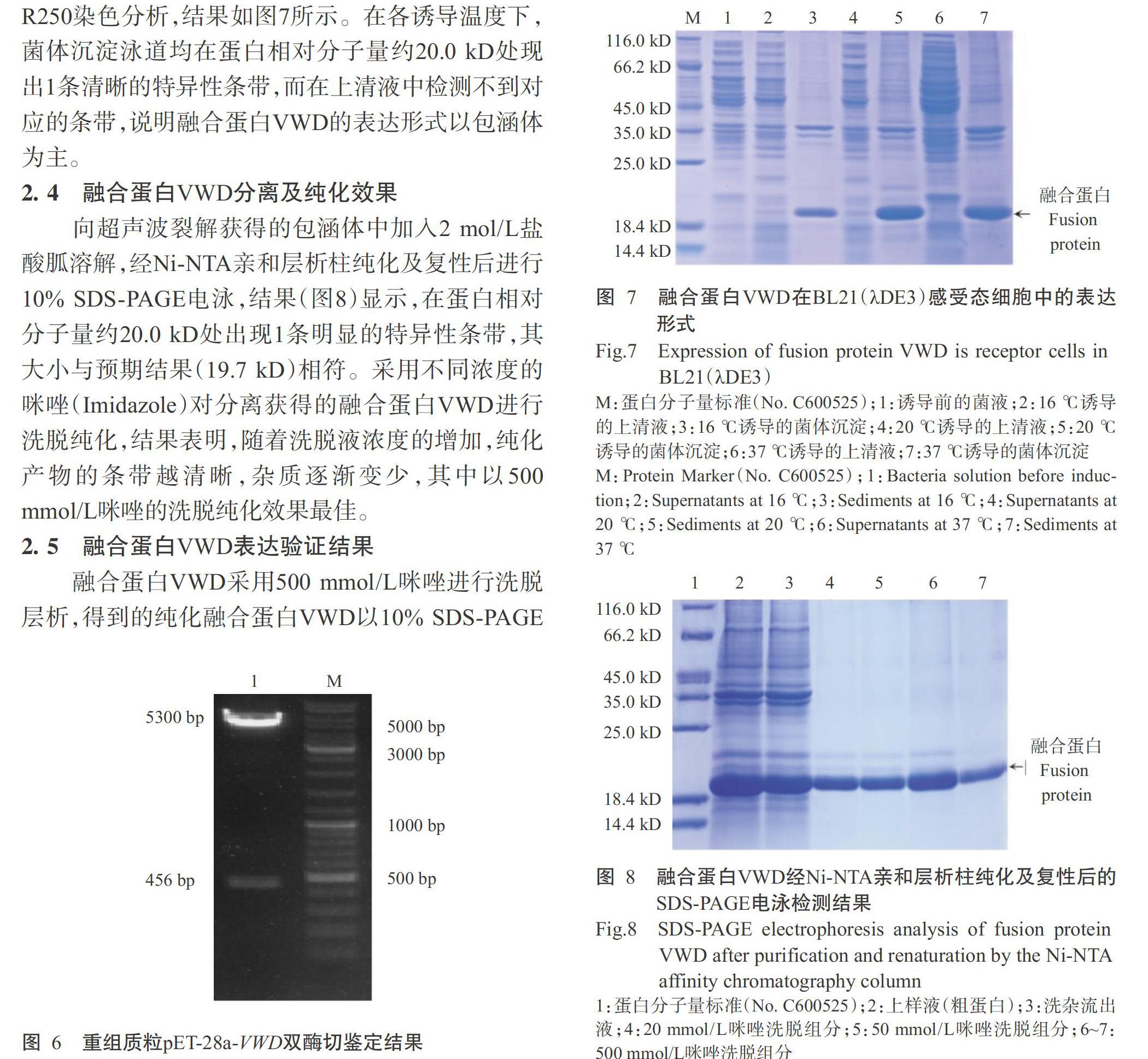
為確定融合蛋白VWD的表達(dá)形式,將擴(kuò)培菌液經(jīng)超聲波裂解后進(jìn)行離心,收集上清液和菌體沉淀,分別采用10% SDS-PAGE電泳進(jìn)行檢測(cè),并進(jìn)行R250染色分析,結(jié)果如圖7所示。在各誘導(dǎo)溫度下,菌體沉淀泳道均在蛋白相對(duì)分子量約20.0 kD處現(xiàn)出1條清晰的特異性條帶,而在上清液中檢測(cè)不到對(duì)應(yīng)的條帶,說(shuō)明融合蛋白VWD的表達(dá)形式以包涵體為主。
2. 4 融合蛋白VWD分離及純化效果
向超聲波裂解獲得的包涵體中加入2 mol/L鹽酸胍溶解,經(jīng)Ni-NTA親和層析柱純化及復(fù)性后進(jìn)行10% SDS-PAGE電泳,結(jié)果(圖8)顯示,在蛋白相對(duì)分子量約20.0 kD處出現(xiàn)1條明顯的特異性條帶,其大小與預(yù)期結(jié)果(19.7 kD)相符。采用不同濃度的咪唑(Imidazole)對(duì)分離獲得的融合蛋白VWD進(jìn)行洗脫純化,結(jié)果表明,隨著洗脫液濃度的增加,純化產(chǎn)物的條帶越清晰,雜質(zhì)逐漸變少,其中以500 mmol/L咪唑的洗脫純化效果最佳。
2. 5 融合蛋白VWD表達(dá)驗(yàn)證結(jié)果
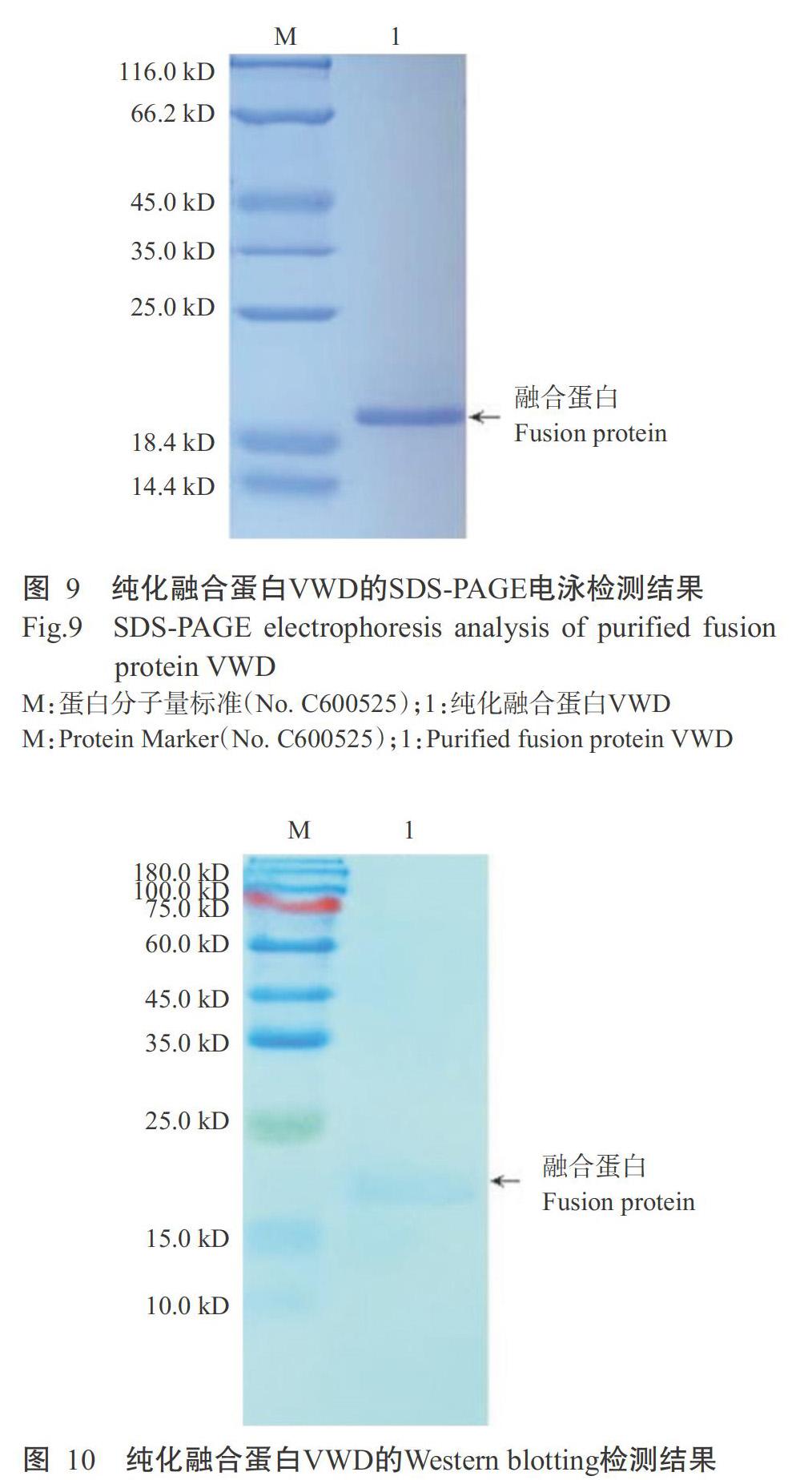
融合蛋白VWD采用500 mmol/L咪唑進(jìn)行洗脫層析,得到的純化融合蛋白VWD以10% SDS-PAGE電泳進(jìn)行檢測(cè),結(jié)果(圖9)顯示,在蛋白相對(duì)分子量約20.0 kD的相應(yīng)位置出現(xiàn)單一、清晰的特異性條帶,無(wú)雜帶,可初步確定融合蛋白VWD在復(fù)性的同時(shí)得到高效純化。Western blotting檢測(cè)結(jié)果(圖10)也發(fā)現(xiàn),在約20.0 kD的相應(yīng)位置出現(xiàn)明顯的特異性條帶,說(shuō)明純化獲得的融合蛋白VWD即為紅螯螯蝦VWD結(jié)構(gòu)域的原核表達(dá)蛋白。采用非干擾型蛋白定量試劑盒測(cè)定純化融合蛋白VWD濃度,得知其濃度為1.32 mg/mL。
3 討論
Vg的VWD結(jié)構(gòu)域具有與病毒或細(xì)菌互作的特性,對(duì)病原微生物進(jìn)行識(shí)別及介導(dǎo)其清除或垂直傳播均有密切聯(lián)系。Vg作為卵黃蛋白的前體,是卵生動(dòng)物胚胎發(fā)育的主要營(yíng)養(yǎng)來(lái)源(羅明坤等,2015;潘賢輝等,2018);Vg作為卵內(nèi)的重要營(yíng)養(yǎng)蛋白,除了已知的營(yíng)養(yǎng)運(yùn)輸功能之外,在進(jìn)入卵細(xì)胞的過(guò)程中還發(fā)揮著其他非營(yíng)養(yǎng)性功能(霍巖等,2018)。近年來(lái)研究發(fā)現(xiàn),Vg以多種功能參與宿主的免疫防御(Salmela et al.,2015),但不同Vg基因編碼的所有Vg蛋白是否都發(fā)揮免疫作用尚未明確,Vg不同結(jié)構(gòu)域與各種免疫功能的相關(guān)性也不清楚。Sun等(2013)從斑馬魚肝臟組織中提取RNA并逆轉(zhuǎn)錄合成cDNA,采用RACE克隆斑馬魚rDUF1943、rDUF1944和rVWD基因cDNA序列,然后構(gòu)建表達(dá)載體進(jìn)行表達(dá)與純化,結(jié)果發(fā)現(xiàn)誘導(dǎo)表達(dá)獲得的融合蛋白rDUF1943、rDUF1944和rVWD均能結(jié)合金黃色葡萄球菌、枯草芽孢桿菌和藤黃微球菌等革蘭氏陽(yáng)性菌及大腸桿菌和鰻弧菌等革蘭氏陰性菌,即DUF1943、DUF1944和VWD可能在Vg發(fā)揮模式識(shí)別受體功能時(shí)扮演著重要角色。本研究通過(guò)對(duì)紅螯螯蝦Vg的VWD結(jié)構(gòu)域進(jìn)行單獨(dú)克隆、表達(dá)及純化,以期制備獲得VWD蛋白用于后續(xù)相關(guān)的生物學(xué)功能及機(jī)制研究。與Sun等(2013)的研究不同,本研究是基于NCBI已公布紅螯螯蝦Vg的VWD結(jié)構(gòu)域(2345~2491 aa)進(jìn)行生物信息學(xué)分析及理論評(píng)估,并結(jié)合大腸桿菌密碼子偏好對(duì)原始VWD氨基酸序列進(jìn)行優(yōu)化及改造,然后通過(guò)全基因合成獲得目的基因,省去了模板制備和文庫(kù)構(gòu)建的相關(guān)步驟。
全基因合成技術(shù)是通過(guò)對(duì)基因序列進(jìn)行改造,優(yōu)化后的目的基因編碼序列更符合宿主細(xì)胞密碼子的偏好性,能有效解決原核生物和真核生物在翻譯前后修飾過(guò)程差異所造成真核生物無(wú)法在大腸桿菌上表達(dá)及正確折疊的問(wèn)題,使目的基因在大腸桿菌中得以正確轉(zhuǎn)錄和翻譯(Papamichail et al.,2016)。本研究結(jié)果表明,重組質(zhì)粒pET-28a-VWD在大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞中高效表達(dá)的最佳誘導(dǎo)條件:當(dāng)單克隆菌液OD600 nm達(dá)0.6時(shí)添加0.5 mmol/L IPTG,置于20 ℃下誘導(dǎo)培養(yǎng)16 h能獲得最大的融合蛋白表達(dá)量;獲得的融合蛋白VWD主要以包涵體形式表達(dá),經(jīng)2 mol/L鹽酸胍溶解和Ni-NTA親和層析純化后可獲得高純度的活性VWD蛋白。大腸桿菌是目前用于重組蛋白異源表達(dá)最成熟的原核表達(dá)系統(tǒng)(謝磊等,2004),是借助胞質(zhì)將外源蛋白通過(guò)可溶性(活性肽)或非可溶性(包涵體)的形式進(jìn)行表達(dá)(張永德等,2018)。大腸桿菌胞質(zhì)是一個(gè)還原性較強(qiáng)的環(huán)境,極易阻礙胞質(zhì)內(nèi)所表達(dá)外源蛋白或多肽中二硫鍵的正確形成,進(jìn)而折疊形成不具生物活性的包涵體(Rosano and Ceccarelli,2014;張永德等,2018)。因此,要獲得活性蛋白用于生物學(xué)功能研究,可對(duì)目的基因進(jìn)行改造,接入信號(hào)肽分子引導(dǎo)外源蛋白分泌到周質(zhì)空間進(jìn)行折疊,或?qū)⒛康牡鞍着c大腸桿菌的外膜蛋白融合表達(dá)也可誘導(dǎo)其分泌到周質(zhì)空間中(Meng et al.,2011;Klint et al.,2013)。本研究通過(guò)密碼子優(yōu)化大量表達(dá)獲得融合蛋白VWD包涵體,后續(xù)使用2 mol/L鹽酸胍對(duì)包涵體進(jìn)行溶解,以降低蛋白結(jié)構(gòu)柔韌性,干擾其聚集,接著采用攜帶His-Tag標(biāo)簽的Ni-NTA親和層析柱對(duì)包涵體進(jìn)行純化及復(fù)性,最后通過(guò)透析去除變性劑以促進(jìn)折疊。
4 結(jié)論
通過(guò)全基因合成獲得的優(yōu)化紅螯螯蝦VWD基因能在大腸桿菌BL21(λDE3)感受態(tài)細(xì)胞中誘導(dǎo)表達(dá)出以包涵體為主要形式的融合蛋白,經(jīng)鹽酸胍溶解和Ni-NTA親和層析柱純化即可獲得高純度的活性VWD蛋白,為后續(xù)開展紅螯螯蝦Vg生物學(xué)功能研究及開發(fā)相應(yīng)的生物活性物質(zhì)提供技術(shù)支持。
參考文獻(xiàn):
霍巖,陳曉英,方榮祥,張莉莉. 2018. 卵黃蛋白原的產(chǎn)生及其非營(yíng)養(yǎng)功能的研究現(xiàn)狀[J]. 生物技術(shù)通報(bào),34(2):66-73. doi:10.13560/j.cnki.biotech.bull.1985.2018-0003. [Huo Y,Chen X Y,F(xiàn)ang R X,Zhang L L. 2018. Study on the production of vitellogenin and its non-nutritional functions[J]. Biotechnology Bulletin,34(2):66-73.]
盧建平,姜乃澄. 2000. 羅氏沼蝦胚胎發(fā)育過(guò)程中同工酶的研究[J]. 東海海洋,18(3):34-39. doi:10.3969/j.issn.1001-909X.2000.03.006. [Lu J P,Jiang N C. 2000. The isozyme patterns in embryo development of the freshwater prawn Macrobrachium rosenbergh[J]. Donghai Marine Science,18(3):34-39.]
羅明坤,趙金良,Thammaratsuntorn Jeerawat,李傳陽(yáng),魏繼海,趙永華. 2015. 尼羅羅非魚卵巢發(fā)育過(guò)程中性類固醇激素與卵黃蛋白原含量變化及Vtg mRNA表達(dá)特征[J].? 中國(guó)水產(chǎn)科學(xué),22(6):1123-1132. doi:10.3724/SP.J.1118. 2015.14467. [Luo M K,Zhao J L,Thammaratsuntorn J,Li C Y,Wei J H,Zhao Y H. 2015. Changes in sex steroid hormones,vitellogenin content,and Vtg mRNA expression during ovarian development in Oreochromis niloticus[J]. Journal of Fishery Sciences of China,22(6):1123-1132.]
潘杰. 2020. 兩種類固醇對(duì)中華絨螯蟹卵黃蛋白原及其受體基因的表達(dá)調(diào)控[D]. 上海:上海海洋大學(xué). [Pan J. 2020. Study on the expression of vitellogenesis-related genes regulated by two steroids in Eriocheir sinensis[D].? Shanghai:Shanghai Ocean University.]
潘賢輝,劉奕,周康奇,胡隱昌,鄭曙明,宋紅梅,牟希東,楊葉欣. 2018. 雙須骨舌魚卵黃蛋白原基因vtg的克隆、組織表達(dá)和原核表達(dá)分析[J]. 動(dòng)物學(xué)雜志,53(4):597-614. doi:10.13859/j.cjz.201804011. [Pan X H,Liu Y,Zhou K Q,Hu Y C,Zheng S M,Song H M,Mu X D,Yang Y X. 2018. Molecular cloning,expression pattern and prokaryo-tic expression of vitellogenin gene(vtg) from Osteoglossum bicirrhosum[J]. Chinese Journal of Zoology,53(4):597-614.]
王加偉,彭露,鄒明民,楊一帆,汪蕾,尤民生. 2016. 昆蟲卵黃原蛋白受體(VgRs)及其主要功能綜述[J]. 環(huán)境昆蟲學(xué)報(bào),38(4):831-842. doi:10.3969/j.issn.1674-0858.2016. 04.25. [Wang J W,Peng L,Zou M M,Yang Y F,Wang L,You M S. 2016. A review of insect vitellogenin receptors(VgRs) and their fundamental functions[J]. Journal of Environmental Entomology,38(4):831-842.]
謝磊,孫建波,張世清,黃俊生. 2004. 大腸桿菌表達(dá)系統(tǒng)及其研究進(jìn)展[J]. 華南熱帶農(nóng)業(yè)大學(xué)學(xué)報(bào),10(2):16-20. doi:10.3969/j.issn.1674-7054.2004.02.005. [Xie L,Sun J B,Zhang S Q,Huang J S. 2004. Research progress in the E. coli expression system[J]. Journal of South China University of Tropical Agriculture,10(2):16-20.]
張永德,林勇,馮鵬霏,陳忠,杜雪松,黃姻,潘傳燕,羅洪林. 2018. 尼羅羅非魚Lck多克隆抗體的制備及鑒定[J]. 南方農(nóng)業(yè)學(xué)報(bào),49(11):2304-2310. doi:10.3969/j.issn.2095-1191.2018.11.27. [Zhang Y D,Lin Y,F(xiàn)eng P F,Chen Z,Du X S,Huang Y,Pan C Y,Luo H L. 2018. Preparation and identification of Lck polyclonal antibody in Nile tilapia[J]. Journal of Southern Agriculture,49(11):2304-2310.]
周慶祥,江桂斌. 2003. 卵黃蛋白原的分離測(cè)定及其在環(huán)境內(nèi)分泌干擾物質(zhì)篩選中的應(yīng)用[J]. 化學(xué)進(jìn)展,15(1):67-73. doi:10.3321/j.issn:1005-281X.2003.01.010. [Zhou Q X,Jiang G B. 2003. Separation and determination and vitellongenin and its application as a bionmarker for the screening of environment endocrine disrupting chemicals[J]. Progress in Chemistry,15(1):67-73.]
Abdu U,Davis C,Khalaila I,Sagi A. 2002. The vitellogenin cDNA of Cherax quadricarinatus encodes a lipoprotein with calcium binding ability,and its expression is induced following the removal of the androgenic gland in a sexually plastic system[J]. General and Comparative Endocriology,127(3):263-272. doi:10.1016/S0016-6480(02)00053-9.
Avarre J C,Lubzens E,Babin P J. 2007. Apolipocrustacein,formerly vitellogenin,is the major egg yolk precursor protein in decapod crustaceans and is homologous to insect apolipophorin II/I and vertebrate apolipoprotein B[J]. BMC Evolutionary Biology,7:3. doi:10.1186/1471-2148-7-3.
Brasset E,Taddei A R,Arnaud F,F(xiàn)aye B,F(xiàn)austo A M,Mazzini M,Giorgi F,Vaury C. 2006. Viral particles of the endogenous retrovirus ZAM from Drosophila melanogaster use a pre-existing endosome/exosome pathway for transfer to the oocyte[J]. Retrovirology,3:25. doi:10.1186/1742-4690-3-25.
Cheng D J,Hou R F. 2005. Determination and distribution of a femalespecific protein in the brown planthopper,Nila-parvata lugens St?l(Homoptera:Delphacidae)[J]. Tissue & Cell,37(1): 37-45. doi:10.1016/j.tice.2004. 09.003.
Dalvin S,F(xiàn)rost P,Loeffen P,Skern-Mauritzen R,Baban J,R?nnestad I,Nilsen F. 2011. Characterisation of two vitellogenins in the salmon louse Lepeophtheirus salmonis:Molecular,functional and evolutional analysis[J]. Disea-ses of Aquatic Organisms,94(3):211-224. doi:10.3354/dao02331.
Garcia J,Munro E S,Monte M M,F(xiàn)ourrier M C S,Whitelaw J,Smail D A,Ellis A E. 2010. Atlantic salmon(Salmo salar L.) serum vitellogenin neutralises infectivity of infectious pancreatic necrosis virus(IPNV)[J]. Fish & Shellfish Immunology,29(2):293-297. doi:10.1016/j.fsi.2010.04.010.
Huo Y,Liu W W,Zhang F J,Chen X Y,Li L,Liu Q F,Zhou Y J,Wei T Y,F(xiàn)ang R X,Wang X F. 2014. Transovarial transmission of a plant virus is mediated by vitellogenin of its insect vector[J]. PLoS Pathogens,10(3):e1003949. doi:10.1371/journal. ppat.1003949.
Klint J K,Senff S,Saez N J,Seshadri R,Lau H Y,Bende N S,Undheim E A B,Rash L D,Mobli M,King G F. 2013. Production of recombinant disulfide-rich venom peptides for structural and functional analysis via expression in the periplasm of E. coli[J]. PLoS One,8(5):e63865. doi:10.1371/journal.pone.0063865.
Leblanc P,Desset S,Giorgi F,Taddei A R,F(xiàn)austo A M,Mazzini M,Dastugue B,Vaury C. 2000. Life cycle of an endogenous retrovirus,ZAM,in Drosophila melanogaster[J]. Journal of Virology,74(22):10658-10669. doi:10. 1128/jvi.74.22.10658-10669.2000.
Li Z J,Zhang S C,Zhang J,Liu M,Liu Z H. 2009. Vitellogenin is a cidal factor capable of killing bacteria via interaction with lipopolysaccharide and lipoteichoic acid[J]. Molecular Immunology,46(16):3232-3239. doi:10. 1016/j.molimm.2009.08.006.
Lu K,Chen X,Liu W T,Zhang X Y,Chen M X,Zhou Q. 2016. Nutritional signaling regulates vitellogenin synthesis and egg development through juvenile hormone in Nilaparvata lugens(St?l)[J]. International Journal of Molecular Sciences,17(3):269. doi:10.3390/ijms17030269.
Luo M W,Li D,Wang Z M,Guo W,Kang L,Zhou S T. 2017. Juvenile hormone differentially regulates two Grp78 genes encoding protein chaperones required for insect fat body cell homeostasis and vitellogenesis[J]. The Journal of Biological Chemistry,292(21):8823-8834. doi: 10.1074/jbc.M117.780957.
Meng E,Cai T F,Li W Y,Zhang H,Liu Y B,Peng K,Liang S P,Zhang D Y. 2011. Functional expression of spider neurotoxic peptide huwentoxin-I in E. coli[J]. PLoS One,6(6):e21608. doi:10.1371/journal.pone.0021608.
Papamichail D,Liu H M,Machado V,Gould N,Coleman J R,Papamichail G. 2016. Codon Context optimization in synthetic gene design[J]. IEEE/ACM Transactions on Computational Biology and Bioinformatics,15(2):452-459. doi:10.1109/TCBB. 2016.2542808.
Rono M K,Whitten M M A,Oulad-Abdelghani M,Levashina E A,Marois E. 2010. The major yolk protein vitellogenin interferes with the anti-plasmodium response in the malaria mosquito Anopheles gambiae[J]. PLoS Biology,8(7):e1000434. doi:10.1371/journal.pbio.1000434.
Rosano G L,Ceccarelli E A. 2014. Recombinant protein expression in Escherichia coli:Advances and challenges[J]. Fronters in Microbiology,5:172. doi:10.3389/fmicb. 2014.00172.
Salmela H,Amdam G V,F(xiàn)reitak D. 2015. Transfer of immunity from mother to offspring is mediated via egg-yolk protein vitellogenin[J]. PLoS Pathogens,11(7):e1005015. doi:10.1371/journal.ppat.1005015.
Shechter A,Aflalo E D,Davis C,Sagi A. 2005. Expression of the reproductive female-specific vitellogenin gene in endocrinologically induced male and intersex Cherax quadricarinatus crayfish[J]. Biology of Reproduction,73(1):72-79. doi:10.1095/biolreprod.104.038554.
Sun C,Hu L L,Liu S S,Gao Z,Zhang S C. 2013. Functional analysis of domain of unknown function(DUF)1943,DUF1944 and von Willebrand factor type D domain (VWD) in vitellogenin 2 in zebrafish[J]. Developmental and Comparative Immunology,41(4):469-476. doi:10. 1016/j.dci.2013.07.005.
Tufail M,Takeda M. 2008. Molecular characteristics of insect vitellogenins[J]. Journal of Insect Physiology,54(12):1447-1458. doi:10.1016/j.jinsphys.2008.08.007.
Wei J,He Y Z,Guo Q,Guo T,Liu Y Q,Zhou X P,Liu S S,Wang X W. 2017. Vector development and vitellogenin determine the transovarial transmission of begomoviruses[J]. Proceedings of the National Academy of Sciences of the United States of America,14(26):6746-6751. doi:10.1073/pnas.1701720114.
(責(zé)任編輯 蘭宗寶)
