萊州灣魚(yú)卵、仔稚魚(yú)群落結(jié)構(gòu)及多樣性的季節(jié)變化
2021-09-09 09:35:00楊艷艷朱明明徐炳慶李凡高彥潔孫春曉魏瀟王斌
海洋學(xué)報(bào) 2021年9期
楊艷艷,朱明明,徐炳慶,李凡*,高彥潔,孫春曉,魏瀟,王斌
( 1.山東省海洋資源與環(huán)境研究院 山東省海洋生態(tài)修復(fù)重點(diǎn)實(shí)驗(yàn)室,山東 煙臺(tái) 264006;2.魯東大學(xué) 濱海生態(tài)高等研究院,山東 煙臺(tái) 264025)
1 引言
魚(yú)卵和仔稚魚(yú)在海洋生態(tài)系統(tǒng)能量傳遞中起重要作用[1],是魚(yú)類(lèi)發(fā)育過(guò)程中的重要階段。同時(shí),魚(yú)卵和仔稚魚(yú)發(fā)育過(guò)程是魚(yú)類(lèi)生命周期中兩個(gè)非常短暫,但形態(tài)學(xué)、生理學(xué)和生態(tài)學(xué)等特性變化明顯的過(guò)渡時(shí)期,也是整個(gè)魚(yú)類(lèi)發(fā)育周期中最為脆弱的階段[2-3]。這一發(fā)育階段成活率的高低、剩存量的多寡將決定魚(yú)類(lèi)補(bǔ)充群體資源量的豐歉,是魚(yú)類(lèi)資源可持續(xù)利用研究中不可缺少的內(nèi)容[4]。從20世紀(jì)50代開(kāi)始,我國(guó)開(kāi)展了多次魚(yú)卵、仔稚魚(yú)的監(jiān)測(cè)調(diào)查工作,1964-1981年在開(kāi)展渤海中國(guó)明對(duì)蝦(Fenneropenaeus chinensis)產(chǎn)卵場(chǎng)調(diào)查時(shí),就收集過(guò)魚(yú)卵、仔稚魚(yú)的資料[5]。
萊州灣位于渤海南部,平均水深為10 m[6-7],是多種經(jīng)濟(jì)種類(lèi)產(chǎn)卵和育肥的場(chǎng)所[8],在漁業(yè)經(jīng)濟(jì)中占重要地位[9-10]。從20世紀(jì)50年代至21世紀(jì)初,我國(guó)生態(tài)學(xué)者、環(huán)境學(xué)者對(duì)該海域漁業(yè)資源的狀況進(jìn)行過(guò)廣泛的研究[11-13]。近60 a來(lái),受過(guò)度捕撈和環(huán)境變化的影響,該海域漁業(yè)資源結(jié)構(gòu)發(fā)生重大變遷[14],生態(tài)環(huán)境質(zhì)量呈現(xiàn)明顯下降的趨勢(shì)[7,15-16]。有報(bào)道顯示[17],2000年以來(lái),萊州灣魚(yú)卵、仔魚(yú)數(shù)量和浮游生物的種數(shù)及多樣性等均呈下降趨勢(shì)。在這種背景下,摸清萊州灣海域魚(yú)卵、仔稚魚(yú)的種類(lèi)組成、數(shù)量分布以及群落結(jié)構(gòu)和多樣性的季節(jié)變化對(duì)了解漁業(yè)資源的補(bǔ)充狀況、保護(hù)渤海生物資源有很重要的現(xiàn)實(shí)意義。近年來(lái),國(guó)內(nèi)有關(guān)萊州灣海域魚(yú)卵、仔稚魚(yú)的研究很多,但是大多數(shù)研究集中在春季和夏季,例如王愛(ài)勇等[18]和高彥潔等[19]對(duì)萊州灣春季的魚(yú)卵、仔稚魚(yú)進(jìn)行了研究;還有一些研究是集中在萊州灣的某一個(gè)特定海域,例如卞曉東等[20]對(duì)黃河口海域的魚(yú)卵和仔稚魚(yú)進(jìn)行了研究;高東奎等[21]對(duì)萊州灣人工魚(yú)礁區(qū)的魚(yú)卵和仔稚魚(yú)進(jìn)行了研究等。有關(guān)萊州灣海域不同季節(jié)的魚(yú)卵、仔稚魚(yú)群落結(jié)構(gòu)和生物多樣性變化的研究較少。
本研究基于2014-2015年萊州灣海域的采樣數(shù)據(jù),分析了該海域魚(yú)卵、仔稚魚(yú)的種類(lèi)組成、數(shù)量分布、優(yōu)勢(shì)種等,探討了該海域魚(yú)卵和仔稚魚(yú)的群落結(jié)構(gòu)和生物多樣性的季節(jié)變動(dòng)規(guī)律,以期為萊州灣海域漁業(yè)資源修復(fù)和保護(hù)提供理論依據(jù)。
2 材料方法
2.1 調(diào)查時(shí)間與站位分布
于2014-2015年在萊州灣海域進(jìn)行4個(gè)航次的魚(yú)類(lèi)浮游生物拖網(wǎng)調(diào)查。調(diào)查時(shí)間分別為2014年5月(春季)、8月(夏季)、10月(秋季)和 2015年 2月(冬季)。在萊州灣海域的調(diào)查范圍(37°10′~37°45′N(xiāo),119°00′~120°10′E)內(nèi)共布設(shè) 20 個(gè)站位(圖1a),由于2015年2月(冬季)受天氣影響,實(shí)際完成調(diào)查12 個(gè)站位(圖1b)。

圖1 萊州灣調(diào)查站位Fig.1 Sample stations in the Laizhou Bay
2.2 樣品采集
樣品采集及分析方法嚴(yán)格按照《海洋調(diào)查規(guī)范》[22]進(jìn)行,租用漁業(yè)生產(chǎn)船只,使用口徑為80 cm,長(zhǎng)為280 cm的大型浮游生物網(wǎng)在海域表層進(jìn)行水平拖拽采集。每站拖拽10 min,拖速為2 n mile/h,采集到的樣品用中性甲醛溶液固定保存,帶回實(shí)驗(yàn)室,根據(jù)魚(yú)卵、仔稚魚(yú)的形態(tài)特征(包括魚(yú)卵和仔稚魚(yú)的外部形態(tài)以及不同發(fā)育時(shí)期的個(gè)體形態(tài)和組織、器官、系統(tǒng)發(fā)育特征等)利用解剖鏡進(jìn)行種類(lèi)鑒定[23-28],并計(jì)數(shù)。
2.3 數(shù)據(jù)分析方法
2.3.1 優(yōu)勢(shì)度
利用相對(duì)重要性指數(shù)(Index of Relative Importance, IRI)對(duì)魚(yú)卵和仔稚魚(yú)的優(yōu)勢(shì)種進(jìn)行計(jì)算[20],公式為

式中,N%為某一種類(lèi)個(gè)體數(shù)量占采獲總量的百分比;F%為某一種類(lèi)出現(xiàn)的站位數(shù)占總調(diào)查站位數(shù)的百分比,IRI不小于1 000的種類(lèi)為優(yōu)勢(shì)種。
2.3.2 生物多樣性
豐富度指數(shù)(D)計(jì)算公式為

香農(nóng)-威納多樣性指數(shù)(H′)計(jì)算公式為

均勻度指數(shù)(J′)計(jì)算公式為

式中,S為種類(lèi)數(shù);N為總個(gè)體數(shù);Pi為第i種個(gè)體數(shù)占總個(gè)體數(shù)的比例。
2.3.3 季節(jié)間種類(lèi)更替率
季節(jié)間種類(lèi)更替率(E)計(jì)算公式為

式中,A為季節(jié)間種類(lèi)增加與減少數(shù)之和;B為季節(jié)間相同的種類(lèi)數(shù)[20]。
2.3.4 群落結(jié)構(gòu)分析
利用R軟件的vegan、cluster等程序包計(jì)算2014-2015年不同季節(jié)間種類(lèi)Bray-Curtis相似性系數(shù)矩陣,并利用非加權(quán)組平均法進(jìn)行聚類(lèi)。
3 結(jié)果與分析
3.1 種類(lèi)組成
調(diào)查期間采集魚(yú)卵135 275粒,仔稚魚(yú)2 456尾,共22種(表1)。其中1種仔稚魚(yú)未鑒定出種,1種鰕虎魚(yú)科仔稚魚(yú)鑒定到科,其余20種隸屬于9目16科20屬。22種魚(yú)卵、仔稚魚(yú)中,魚(yú)卵13種,仔稚魚(yú)15種。鱸形目種類(lèi)最多,為7種,鯡形目4種,鰈形目3種,鯔形目2種,頜針魚(yú)目、銀漢魚(yú)目、鼠鱚目、刺魚(yú)目、鲉形目和胡瓜魚(yú)目各1種。

表1 萊州灣魚(yú)卵、仔稚魚(yú)種類(lèi)組成Table 1 Species composition of ichthyoplankton in the Laizhou Bay
3.2 優(yōu)勢(shì)種及其季節(jié)變化
5月(春季)和8月(夏季)調(diào)查中,魚(yú)卵的優(yōu)勢(shì)種分別為鳀(Engraulis japonicus)和赤鼻棱鳀(Thryssa kammalensis),仔稚魚(yú)的優(yōu)勢(shì)種分別為鮻(Liza haematocheilus)、布氏銀漢魚(yú)(Allanetta bleekeri),仔稚魚(yú)冬季的優(yōu)勢(shì)種為長(zhǎng)綿鳚(Zoarces elongatus)。10月(秋季)和2月(冬季)未采獲到魚(yú)卵所以無(wú)法計(jì)算優(yōu)勢(shì)種,10月(秋季)調(diào)查所采集的仔稚魚(yú)樣品中無(wú)優(yōu)勢(shì)種存在,僅大銀魚(yú)(Protosalanx hyalocranius)、中頜棱鳀(Thryssa mystax)和青鱗小沙丁(Sardinella zunasi)構(gòu)成重要種(表2)。

表2 萊州灣海域魚(yú)卵、仔稚魚(yú)優(yōu)勢(shì)種和重要種組成Table 2 Dominant and important species composition of ichthyoplankton in the Laizhou Bay
3.3 豐度時(shí)間和空間變化
調(diào)查中共采集魚(yú)卵135 275粒,仔稚魚(yú)2 456尾。魚(yú)卵、仔稚魚(yú)平均密度分別為1 878.82粒/站和34.11 尾/站。
5月(春季)采獲魚(yú)卵135 118粒,平均密度為6 755.90粒/站,波動(dòng)范圍為8~112 340粒/網(wǎng)。最高值出現(xiàn)在3號(hào)站,最低值出現(xiàn)在1號(hào)和7號(hào)站位(圖2a)。其中,鳀卵數(shù)量最多,占本航次調(diào)查魚(yú)卵數(shù)量的88.32%。8月(夏季)采獲魚(yú)卵157粒,平均密度7.85粒/站,波動(dòng)范圍為0~95粒/網(wǎng),最高值出現(xiàn)在18號(hào)站位,為95粒/站,赤鼻棱鳀魚(yú)卵數(shù)量最多,占本航次調(diào)查魚(yú)卵數(shù)量的66.88%(圖2b)。秋季和冬季沒(méi)有采獲魚(yú)卵(圖2c,圖2d)。

圖2 萊州灣海域魚(yú)卵空間分布Fig.2 Distribution of fish eggs in the Laizhou Bay
5月(春季)采獲仔稚魚(yú)1 948尾,平均密度為97.4尾/站,波動(dòng)范圍為0~760尾/網(wǎng),最高值出現(xiàn)在13號(hào)站位,為760尾/站(圖3a)。其中,鮻仔稚魚(yú)的數(shù)量最多,占本航次調(diào)查仔稚魚(yú)數(shù)量的57.39%。8月(夏季)采獲仔稚魚(yú)198尾,平均密度為9.9尾/站,波動(dòng)范圍為0~35尾/網(wǎng),最高值出現(xiàn)在16號(hào)站位(圖3b)。其中,布氏銀漢魚(yú)數(shù)量最多,占本航次調(diào)查仔稚魚(yú)數(shù)量的50%。10月(秋季)采獲仔稚魚(yú)24尾,平均密度為1.2尾/站,波動(dòng)范圍為0~13尾/網(wǎng),最高值出現(xiàn)在1號(hào)站位(圖3c)。其中大銀魚(yú)仔稚魚(yú)數(shù)量最多,占本航次調(diào)查仔稚魚(yú)數(shù)量的84.62%。2月(冬季)采獲仔稚魚(yú)286尾,平均密度為23.83尾/站,最高值出現(xiàn)在4號(hào)站位(圖3d)。全部為長(zhǎng)綿鳚的仔稚魚(yú)。從全年變化范圍來(lái)看,魚(yú)卵、仔稚魚(yú)分布不均勻,資源密度總體呈現(xiàn)春夏高,秋冬低的趨勢(shì)。

圖3 萊州灣海域仔稚魚(yú)空間分布Fig.3 Distribution of larvae in the Laizhou Bay
3.4 群落多樣性的季節(jié)變化
2015年2 月(冬季),因只采獲到長(zhǎng)綿鳚1個(gè)種類(lèi),所以豐富度指數(shù)(D)和多樣性指數(shù)(H′)均為0,均勻度指數(shù)(J′)則無(wú)法計(jì)算。由圖4可知,5月(春季)的豐富度指數(shù)(D)和多樣性指數(shù)(H′)均為全年最低,均勻度指數(shù)(J′)則為除 2月(冬季)外最低。8月(夏季)的豐富度指數(shù)(D)和多樣性指數(shù)(H′)均為最高,豐富度指數(shù)(D)和多樣性指數(shù)(H′)全年呈先升高后下降的趨勢(shì);10月(秋季)的均勻度指數(shù)(J′)為除2月外全年最高,均勻度指數(shù)(J′)全年除2月外呈現(xiàn)逐漸上升的趨勢(shì)。

圖4 萊州灣海域魚(yú)卵、仔稚魚(yú)物種多樣性Fig.4 Species diversity of ichthyoplankton in the Laizhou Bay
3.5 種類(lèi)更替和群落結(jié)構(gòu)的季節(jié)間相似性聚類(lèi)
由圖5可知,2014-2015年調(diào)查期間萊州灣海域各季節(jié)間魚(yú)卵和仔稚魚(yú)的種類(lèi)更替率均在80%以上,且隨季節(jié)變化呈現(xiàn)上升的趨勢(shì),從春季到夏季的種類(lèi)更替率最小,為89.47%,其次為夏季到秋季,為92.31%,秋季到冬季以及冬季到春季的種類(lèi)更替率均為 100%(圖5)。

圖5 萊州灣海域魚(yú)卵、仔稚魚(yú)種類(lèi)更替率Fig.5 The species turnover rate of ichthyoplankton in the Laizhou Bay
利用Bray-Curtis相似性系數(shù)公式計(jì)算不同季節(jié)間種類(lèi)組成的相似性系數(shù),然后運(yùn)用R語(yǔ)言聚類(lèi)樹(shù)對(duì)其進(jìn)行聚類(lèi)分析,根據(jù)融合水平值定義劃分水平,選擇具有最大跳躍的分組水平,將4個(gè)季節(jié)的魚(yú)卵、仔稚魚(yú)劃分為3個(gè)組,進(jìn)一步聚類(lèi)分析結(jié)果顯示,3個(gè)分組分別為春組、夏秋組和冬組(圖6,圖7),其中,春組和夏秋組、春組和冬組主要分歧種均為鳀,鳀對(duì)其季節(jié)間平均相異性貢獻(xiàn)率分別為87.28%和87.22%;夏秋組和冬組間主要分歧種為長(zhǎng)綿鳚和赤鼻棱鳀,這兩個(gè)種類(lèi)對(duì)兩個(gè)組季節(jié)間平均相異性貢獻(xiàn)率為76.63%。

圖6 聚類(lèi)樹(shù)的融合水平值Fig.6 Fusion level values of cluster tree

圖7 萊州灣海域魚(yú)卵、仔稚魚(yú)群落結(jié)構(gòu)季節(jié)變化聚類(lèi)分析Fig.7 Cluster analysis of seasonal changes in the community structure of ichthyoplankton in the Laizhou Bay
4 討論
4.1 種類(lèi)組成和數(shù)量分布的季節(jié)更替
本研究調(diào)查結(jié)果顯示,萊州灣海域魚(yú)卵、仔稚魚(yú)的種類(lèi)和數(shù)量隨季節(jié)而更替,從魚(yú)卵、仔稚魚(yú)數(shù)量的季節(jié)變化來(lái)看,春季(5月)為魚(yú)類(lèi)的產(chǎn)卵盛期,魚(yú)卵和仔稚魚(yú)的數(shù)量最多,占全年總量的90%以上,夏季數(shù)量大幅減少,秋冬季節(jié)進(jìn)入產(chǎn)卵末期以后數(shù)量也隨之降至最低點(diǎn),兩季魚(yú)卵和仔稚魚(yú)數(shù)量?jī)H占全年數(shù)量的0.2%左右[7]。
從種類(lèi)數(shù)的季節(jié)變化來(lái)看,5月(春季)捕獲魚(yú)卵、仔稚魚(yú) 11種,8月(夏季)9種,10月(秋季)5種,2月(冬季)1種,種類(lèi)數(shù)呈現(xiàn)隨季節(jié)轉(zhuǎn)變而下降的趨勢(shì),這種季節(jié)變化主要是洄游性種類(lèi)的季節(jié)性分布引起的:春季洄游性的種類(lèi)(主要為暖水種和暖溫種) 進(jìn)入萊州灣產(chǎn)卵、索餌,這一時(shí)期種數(shù)較冬季增加;秋末,隨著水溫的降低,這些種類(lèi)離開(kāi)萊州灣至越冬場(chǎng),僅定居種留下[29]。
從種類(lèi)組成的季節(jié)變化來(lái)看,5月(春季)為一年中的產(chǎn)卵盛期,此時(shí)魚(yú)卵集中在灣口海域,仔稚魚(yú)則集中在西部海域,魚(yú)卵、仔稚魚(yú)的優(yōu)勢(shì)種為陸續(xù)進(jìn)入灣口等近岸水域產(chǎn)卵的鳀和從深水區(qū)回到河口產(chǎn)卵的鮻,5月調(diào)查中捕獲鮻仔稚魚(yú)1 118尾,魚(yú)卵89粒,且鮻多為初孵的仔魚(yú)(初孵仔魚(yú)占鮻仔稚魚(yú)總數(shù)的98.2%),而鮻的孵化時(shí)間為42 h,所以分析可能是調(diào)查時(shí)間錯(cuò)過(guò)了鮻產(chǎn)卵盛期;春季到夏季的種類(lèi)更替率為89.47%,魚(yú)卵、仔稚魚(yú)的優(yōu)勢(shì)種從春季暖溫性中上層洄游魚(yú)類(lèi)的鳀和暖溫性河口定居魚(yú)類(lèi)鮻變?yōu)橄募九灾猩蠈愉в昔~(yú)類(lèi)赤鼻棱鳀和布氏銀漢魚(yú),且魚(yú)卵優(yōu)勢(shì)種優(yōu)勢(shì)度下降,仔稚魚(yú)優(yōu)勢(shì)種優(yōu)勢(shì)度上升。因?yàn)橄募窘端疁乇冗h(yuǎn)岸高,所以此時(shí)魚(yú)卵、仔稚魚(yú)的分布向?yàn)车捉都小O募竞颓锛局g的種類(lèi)更替率為92.31%,秋季和冬季之間的種類(lèi)更替率為100%,夏季到秋季再到冬季主要種類(lèi)變化依次為暖水性中上層洄游魚(yú)類(lèi)的赤鼻棱鳀和布氏銀漢魚(yú)變?yōu)榍锛九瘻匦灾猩蠈铀莺愉в涡贼~(yú)類(lèi)大銀魚(yú),再到冬季冷溫性底層魚(yú)類(lèi)長(zhǎng)綿鳚,形成了種類(lèi)與環(huán)境和季節(jié)相對(duì)應(yīng)的動(dòng)態(tài)格局[20],同時(shí)也說(shuō)明在不同的季節(jié),產(chǎn)卵群體的組成和結(jié)構(gòu)也發(fā)生了顯著的變化,而季節(jié)間種類(lèi)的高更替率則說(shuō)明魚(yú)類(lèi)群落結(jié)構(gòu)的時(shí)序相對(duì)不穩(wěn)定[20]。分析這種不穩(wěn)定產(chǎn)生的原因,一方面與萊州灣物種洄游性季節(jié)分布有關(guān),另一方面由于近年來(lái)萊州灣海域的環(huán)境變化和人類(lèi)活動(dòng)等因素,使種類(lèi)更替率明顯加快[30-31]。另外,秋冬季節(jié)沒(méi)有捕獲到魚(yú)卵,這是因?yàn)楸狙芯恐姓{(diào)查網(wǎng)具為浮游生物網(wǎng),調(diào)查方法為表層拖網(wǎng),而萊州灣海域秋冬季節(jié)僅有極少數(shù)產(chǎn)浮性卵的底棲魚(yú)類(lèi),細(xì)紋獅子魚(yú)(Liparis tanakae)、大瀧六線魚(yú)(Hexagrammos otakii)等秋冬季節(jié)繁殖的魚(yú)類(lèi)均產(chǎn)黏著沉性卵,不適于浮游生物網(wǎng)采集。
4.2 物種多樣性
由于春、夏季是萊州灣海域魚(yú)類(lèi)繁殖的重要季節(jié),有關(guān)秋、冬季魚(yú)卵、仔稚魚(yú)多樣性指數(shù)(H′)可參考的數(shù)據(jù)很少,所以本研究中僅對(duì)萊州灣海域春、夏季不同調(diào)查時(shí)期的多樣性指數(shù)(H′)進(jìn)行對(duì)比。
馬克平和劉玉明[32]提出,多樣性指數(shù)(H′)的合理范圍為1.5~3.5,本研究中萊州灣海域魚(yú)卵、仔稚魚(yú)群落的多樣性指數(shù)(H′)只有夏季處于此范圍之內(nèi),春季、秋季和冬季均低于該范圍,將春季多樣性指數(shù)(H′)與萊州灣以及鄰近海域不同時(shí)期調(diào)查相比較(表3),發(fā)現(xiàn)其均處于較低水平。同時(shí),魚(yú)卵、仔稚魚(yú)群落D、J′也都較低,均低于夏季和秋季,說(shuō)明春季魚(yú)卵、仔稚魚(yú)的結(jié)構(gòu)相對(duì)不穩(wěn)定,種間分布不均勻[20]。魚(yú)類(lèi)群落在長(zhǎng)期嚴(yán)重的外界干擾下,物種多樣性會(huì)喪失[33],而人類(lèi)的捕撈是對(duì)魚(yú)類(lèi)群落最主要的外界干擾因素,在人類(lèi)捕撈活動(dòng)的長(zhǎng)期干擾下,魚(yú)類(lèi)群落多樣性不斷下降,從而導(dǎo)致作為其補(bǔ)充群體的魚(yú)卵、仔稚魚(yú)的群落多樣性也不斷下降[34],另外,根據(jù)多樣性指數(shù)的生態(tài)含義分析,在種類(lèi)數(shù)較多,但是種間分布極不均勻的情況下,多樣性偏低。某些優(yōu)勢(shì)種在個(gè)別海區(qū)的聚集,使這些站位個(gè)體數(shù)量在不同種間分配差異加大,也會(huì)使多樣性偏低[35-36]。本研究符合這種情況:春季航次魚(yú)卵種類(lèi)雖多,但是除鳀以外的其他種類(lèi)數(shù)量較少,鳀占該航次魚(yú)卵總數(shù)的88.3%,成為該航次的絕對(duì)優(yōu)勢(shì)種;而鳀仔稚魚(yú)雖然沒(méi)有成為春季航次仔稚魚(yú)的絕對(duì)優(yōu)勢(shì)種,但是其數(shù)量占該航次仔稚魚(yú)總數(shù)量的近1/3,并且分布極不均勻,只在1號(hào)站位出現(xiàn),這就導(dǎo)致了春季魚(yú)卵、仔稚魚(yú)的H′較低。
夏季H′在與2007年夏季黃河口以南和以北海域的比較中均處于較高水平(表3),但是因?yàn)閰⒖嘉墨I(xiàn)的局限性,只有1 a的數(shù)據(jù)作為參考,并且其調(diào)查區(qū)域的H′可能會(huì)受到黃河調(diào)水調(diào)沙的影響而導(dǎo)致偏小,所以不能確定2014年夏季H′與萊州灣及鄰近海域不同時(shí)期調(diào)查相比是否處于較高水平,有待于今后的調(diào)查和研究中進(jìn)一步驗(yàn)證。另外,夏季的D和H′均大于秋季,但是J′略小于秋季。因?yàn)榍锛痉N類(lèi)和數(shù)量分布比較均勻,魚(yú)卵和仔稚魚(yú)均沒(méi)有出現(xiàn)優(yōu)勢(shì)種;夏季種類(lèi)和數(shù)量雖超過(guò)秋季,但是出現(xiàn)了赤鼻棱鳀和布氏銀漢魚(yú)兩個(gè)相對(duì)高優(yōu)勢(shì)度的物種,所以導(dǎo)致J′略小于秋季。魚(yú)類(lèi)的繁殖生長(zhǎng)和捕撈活動(dòng)也會(huì)造成多樣性的季節(jié)變化[37],8月的休漁,使萊州灣海域的外界干擾減小,而10月秋季所處時(shí)間段為開(kāi)捕后,人類(lèi)捕撈干擾增強(qiáng),加上水溫降低,導(dǎo)致洄游性魚(yú)類(lèi)離開(kāi)近岸水域,魚(yú)類(lèi)種類(lèi)數(shù)和數(shù)量均減少,所以D和H′均出現(xiàn)下降趨勢(shì)。
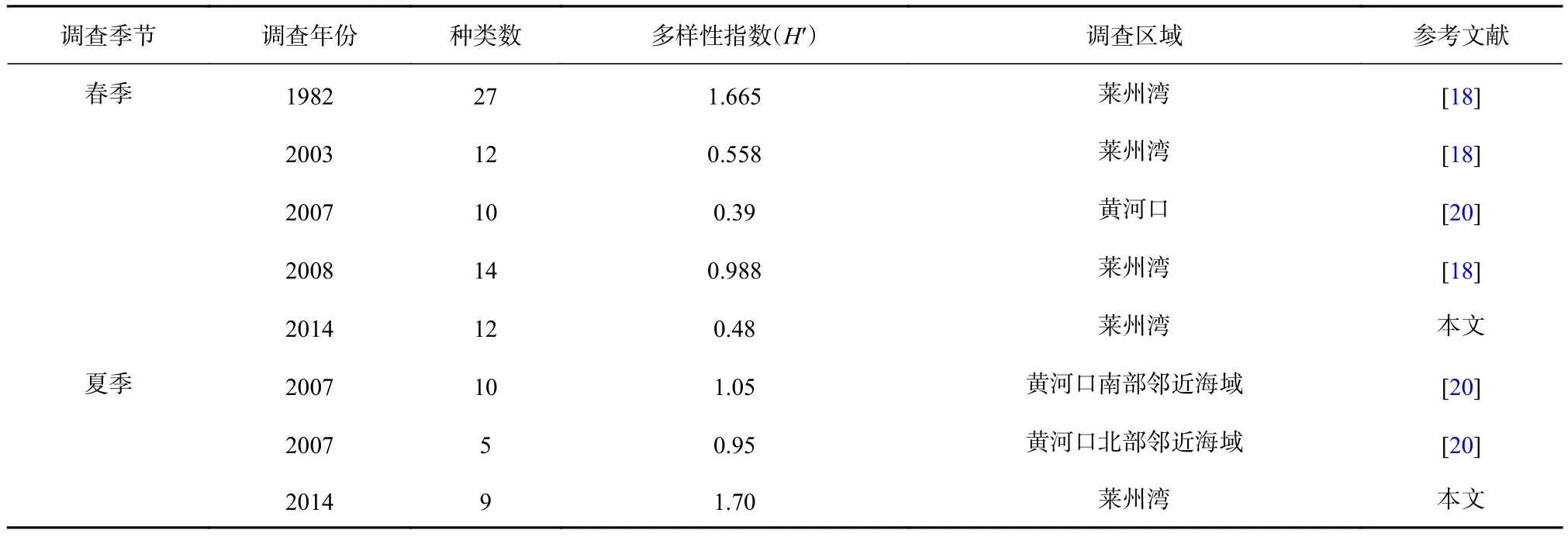
表3 萊州灣及鄰近海域不同時(shí)期調(diào)查春、夏季魚(yú)卵、仔稚魚(yú)的群落多樣性( H′ )Table 3 Community diversity of ichthyoplankton in the Laizhou Bay and its adjacent waters in different periods (H′)
4.3 群落結(jié)構(gòu)
萊州灣水域是渤海的傳統(tǒng)漁場(chǎng),也是黃、渤海眾多漁業(yè)生物的產(chǎn)卵場(chǎng)、育幼場(chǎng)和索餌場(chǎng)[14],對(duì)黃、渤海漁業(yè)資源的補(bǔ)充具有重要意義[10-11]。近年來(lái),受人類(lèi)活動(dòng)以及環(huán)境變化等的共同影響,使得萊州灣的漁業(yè)資源結(jié)構(gòu)發(fā)生了重大變遷。
從魚(yú)卵、仔稚魚(yú)種類(lèi)組成來(lái)看,本研究與黃、渤海海域大面調(diào)查的結(jié)果存在一定的差異,程濟(jì)生[38]于1998年5-11月在渤海海域進(jìn)行調(diào)查,采獲魚(yú)卵、仔稚魚(yú)共41種。萬(wàn)瑞景和姜言偉[39-40]于1982年4月至1983年4月和1992年8月至1993年6月在渤海海域的調(diào)查中,共采獲魚(yú)卵61種、仔稚魚(yú)49種;1985-1988年在北黃海和南黃海的調(diào)查中采獲68種魚(yú)卵、仔稚魚(yú)。從與黃、渤海大面調(diào)查的歷史資料縱向?qū)Ρ葋?lái)看,萊州灣海域魚(yú)卵、仔稚魚(yú)的種類(lèi)數(shù)僅為或者不足黃、渤海海域20世紀(jì)80、90年代的1/2。說(shuō)明萊州灣海域魚(yú)卵、仔稚魚(yú)的種類(lèi)在大幅減少,主要與近年來(lái)的過(guò)度捕撈和環(huán)境退化有關(guān)。而于旭光等[41]2008年在遼東灣近岸海域的調(diào)查以及郭書(shū)新等[4]2014年和2015年在青島嶗山青山灣人工魚(yú)礁區(qū)調(diào)查的種類(lèi)數(shù)均低于本研究;肖歡歡等[42]2015年在海州灣及鄰近海域3個(gè)航次的調(diào)查采獲魚(yú)卵25種,仔稚魚(yú)13種,高于本研究采獲的種類(lèi)數(shù)。從與山東海域其他海灣的橫向?qū)Ρ葋?lái)看,種類(lèi)數(shù)目高低不同,種類(lèi)組成也存在一定的差異,說(shuō)明魚(yú)卵、仔稚魚(yú)的種類(lèi)組成因調(diào)查海域范圍、調(diào)查時(shí)間以及調(diào)查海域的生態(tài)環(huán)境等因素的不同而存在差異,這種差異也反映了不同海域不同時(shí)期魚(yú)類(lèi)群落結(jié)構(gòu)的差異[4]。另外,將本研究中優(yōu)勢(shì)種種類(lèi)與1998年渤海調(diào)查相比,油魣(Sphyraena pinguis)、棘頭梅童魚(yú)(Collichthys lucidus)、叫姑魚(yú)(Johnius belangerii)、小黃魚(yú)(Larimichthys polyactis)、藍(lán)點(diǎn)馬鮫(Scomberomorus niphonius)、玉筋魚(yú)(Ammodytes personatus)、短鰭紅娘魚(yú)(Lepidotrigla micropterus)、許氏平鲉(Sebastods schlegelii)、大瀧六線魚(yú)等均缺失[7]。而鳀、鮻、赤鼻棱鳀、斑鰶(Konosirus punctatus)等種類(lèi)比例有所上升。并且2014-2015年間,高優(yōu)勢(shì)度物種的季節(jié)演替較為明顯,不存在高優(yōu)勢(shì)度物種的季節(jié)間重疊,與膠州灣和青山灣的研究結(jié)果一致,證明種類(lèi)更替呈現(xiàn)加快的趨勢(shì)[4,43]。
從空間生態(tài)類(lèi)型來(lái)看,萊州灣海域目前以小型中上層低經(jīng)濟(jì)價(jià)值魚(yú)類(lèi)的魚(yú)卵、仔稚魚(yú)為主,中上層魚(yú)類(lèi)和經(jīng)濟(jì)價(jià)值較低魚(yú)類(lèi)的魚(yú)卵、仔稚魚(yú)均超過(guò)全年總量的95%,而底層魚(yú)類(lèi)和經(jīng)濟(jì)價(jià)值較高魚(yú)類(lèi)的魚(yú)卵、仔稚魚(yú)分別僅占全年數(shù)量的3.7%和2.1%。進(jìn)一步驗(yàn)證了目前萊州灣漁業(yè)資源結(jié)構(gòu)正在向中上層、小型低值魚(yú)類(lèi)類(lèi)群演化的趨勢(shì)[11,44-45]。這種演變趨勢(shì)主要與環(huán)境條件的變化和人類(lèi)的捕撈活動(dòng)有關(guān)。持續(xù)的全球變暖使渤海的水溫不斷升高,造成魚(yú)類(lèi)早期補(bǔ)充群體的生理周期提前,導(dǎo)致其與餌料的錯(cuò)配,使魚(yú)類(lèi)早期補(bǔ)充能力下降,進(jìn)而導(dǎo)致漁業(yè)資源的衰退[46]。另外,有研究表明,渤海內(nèi)捕撈強(qiáng)度自1950年以來(lái)增長(zhǎng)了近40倍[47],在環(huán)境變化和捕撈活動(dòng)的雙重干擾下,渤海魚(yú)類(lèi)資源急劇下降[16,48-49],小黃魚(yú)、帶魚(yú)(Trichiurus japonicas)等種類(lèi),曾是山東近海最重要的底層經(jīng)濟(jì)魚(yú)類(lèi),自1962年秋捕對(duì)蝦以來(lái),大量損害了它們的幼魚(yú),不論在育肥場(chǎng)還是越冬場(chǎng)的密集區(qū),均遭到大量的捕撈,極大破壞了底層經(jīng)濟(jì)魚(yú)類(lèi)資源。進(jìn)入21世紀(jì)以來(lái),底層魚(yú)類(lèi)比例仍處在較低水平上。而小型中上層魚(yú)類(lèi)的數(shù)量開(kāi)始增加,20世紀(jì)90年代以后,鳀、斑鰶等小型中上層魚(yú)類(lèi)逐漸成為黃、渤海主要的捕撈品種[9,16,50-51],從而導(dǎo)致作為魚(yú)類(lèi)早期補(bǔ)充群體的魚(yú)卵、仔稚魚(yú)的結(jié)構(gòu)也發(fā)生相應(yīng)的改變[31]。
從攝食習(xí)性來(lái)看,本研究中親體為浮游動(dòng)物食性和底棲動(dòng)物食性的魚(yú)卵、仔稚魚(yú)大約占采集樣品種數(shù)的70%,說(shuō)明萊州灣目前以浮游動(dòng)物食性和底棲動(dòng)物食性的魚(yú)類(lèi)為主[52]。Jin等[51]發(fā)現(xiàn)萊州灣的優(yōu)勢(shì)種從高營(yíng)養(yǎng)級(jí)和廣食性魚(yú)類(lèi),向壽命短、浮游生物食性的低營(yíng)養(yǎng)級(jí)中上層魚(yú)類(lèi)演替。同時(shí),渤海生態(tài)系統(tǒng)捕撈漁獲物的平均營(yíng)養(yǎng)級(jí)由1982年的2.92下降到2014-2015年的2.68,總的變化趨勢(shì)為低營(yíng)養(yǎng)層次種類(lèi)的生物量百分比增加,高營(yíng)養(yǎng)層次種類(lèi)的生物量百分比減少[53-55]。過(guò)度捕撈和環(huán)境退化導(dǎo)致渤海生態(tài)系統(tǒng)穩(wěn)定性不斷降低,萊州灣生態(tài)系統(tǒng)壓力巨大,而海灣區(qū)域多是水生生物重要的棲息地和重要的產(chǎn)卵場(chǎng)、育幼場(chǎng)、索餌場(chǎng)所,對(duì)漁業(yè)資源的補(bǔ)充非常重要。因此,加強(qiáng)對(duì)海灣生態(tài)系統(tǒng)的恢復(fù)和保護(hù)是當(dāng)前非常緊迫的任務(wù)。
