肝細胞癌關鍵基因的生物信息學分析
2021-09-10 07:22:44隋英麗盧坤傅琳
青島大學學報(自然科學版) 2021年2期
隋英麗 盧坤 傅琳
摘要:肝細胞癌是一種高死亡率的原發(fā)性肝癌,其有限治療和較低的化療敏感性,使得迫切需要尋找潛在的臨床治療靶點和預后判斷的生物標志物.為此,采用生物信息學方法對肝細胞癌發(fā)生發(fā)展的關鍵基因進行挖掘.從基因表達數(shù)據(jù)庫(GEO)中下載數(shù)據(jù)集,并篩選差異表達基因,使用在線數(shù)據(jù)庫DAVID68對篩選到的DEGs進行基因本體(GO)富集分析和京都基因與基因百科全書(KEGG)通路分析,使用在線數(shù)據(jù)庫STRING構建蛋白質-蛋白質相互作用網絡(PPI),利用Cytoscape軟件篩選核心DEGs,并進行GO和KEGG富集分析,使用UALCAN和KaplanGMeierplotter在線數(shù)據(jù)庫篩選并驗證關鍵基因的表達以及生存預后.研究結果表明,篩選出6個關鍵基因RAD51AP1、FANCI、SMC2、POLE2、CENPN、WDHD1,與HCC的發(fā)生發(fā)展以及生存預后顯著相關,可能是HCC的潛在治療靶點,可能為HCC的內在機制的探究提供理論依據(jù).
關鍵詞:生物信息學;差異表達基因;蛋白質-蛋白質相互作用網絡;肝細胞癌;富集分析
中圖分類號:R735.7文獻標志碼:A
近年來,生物信息學分析結合基因芯片技術已經廣泛應用于基因相關腫瘤的研究當中.基因芯片可以高通量地完成基因篩選與分析,在醫(yī)學研究領域占據(jù)重要位置.肝細胞癌(HepatocellularCarcinoma,HCC)是全球最常見的癌癥之一[1],分為原發(fā)性和繼發(fā)性兩大類,是與癌癥相關死亡的第二大主要原因,也是世界范圍內的主要公共衛(wèi)生問題[2],近年來發(fā)病率仍然呈現(xiàn)不斷上升趨勢.HCC的發(fā)生發(fā)展是涉及多種因素的復雜過程,其主要危險因素與丙型肝炎后的持續(xù)病毒學反應、治療期間抑制的乙型肝炎病毒以及酒精性和非酒精性脂肪性肝病相關[3].早期肝細胞癌癥狀無特異性,且缺乏客觀的診斷標準[4],中晚期肝癌的癥狀則較多,但是當HCC患者診斷時,大多已經發(fā)展成中晚期等治療困難的階段,肝癌的有限治療和較低的化療敏感性[5],使得迫切需要尋找潛在的臨床治療靶點以及早期診斷和預后判斷的生物標志物.本研究基于生物信息學方法,從基因表達數(shù)據(jù)庫(GeneExpressionOmnibus,GEO)中下載了大量的HCC芯片數(shù)據(jù),包含HCC癌組織和癌旁正常組織,運用生物信息學方法篩選差異表達基因(DifferentiallyExpressedGenes,DEGs),對DEGs進行基因本體GeneDntology,GO)富集分析和KEGG(KyotoEncyclopediaofGenesandGenomes)通路分析,構建蛋白質—蛋白質相互作用網絡(ProteinGProteinInteractionnetwork,PPI)篩選出6個HCC發(fā)生發(fā)展相關的關鍵基因,并進一步對這些關鍵基因進行了表達和生存分析的驗證,在多個在線數(shù)據(jù)庫中進行了全面系統(tǒng)的分析.本研究可能為HCC尋找潛在的臨床治療靶點、探究HCC發(fā)生發(fā)展的內在機制提供理論基礎.
1材料與方法
1.1數(shù)據(jù)來源
從GEO數(shù)據(jù)庫(http://www.ncbi.nlm.nih.gov/GEO/)檢索并下載HCC相關數(shù)據(jù)集GSE60502[6]和GSE84402[7].其中GSE60502數(shù)據(jù)集對應的檢測平臺GPL96,GSE84402數(shù)據(jù)集對應的檢測平臺名稱為GPL570.選擇每個數(shù)據(jù)集中的HCC樣本數(shù)據(jù)及其匹配的正常樣本數(shù)據(jù)進行分析.其中GSE60502數(shù)據(jù)集包含腫瘤樣本18個和正常樣本18個,GSE84402數(shù)據(jù)集包含腫瘤樣本14個和正常樣本14個.
1.2數(shù)據(jù)的處理
利用GEO2R(http://www.ncbi.nlm.nih.gov/geo/geo2r)在線分析,導出結果并匯總,初步篩選后,刪除沒有基因名稱或探針以及同一個基因名稱對應多個基因探針的重復數(shù)據(jù).最后對篩選的DEGs進行火山圖形式的可視化分析.
1.3數(shù)據(jù)的篩選
對數(shù)據(jù)再次篩選,篩選標準:P<001,差異倍數(shù)logFC>1或logFC<-1.將2個數(shù)據(jù)集中的上調基因或下調基因分別導入在線網站Bioinformatics&EvolutionaryGenomic(http://bioinformaticspsb.ugent.be/webtools/Venn/)中,取2個數(shù)據(jù)集中上調基因或下調基因的交集.
1.4GO分析和KEGG富集分析
利用DAVID6.8在線數(shù)據(jù)庫[8](https://david.ncifcrf.gov/)分析組學和相關數(shù)據(jù)[9].GO用于分析大量注釋基因的生物學過程,分為生物學過程(BiologicalProcess,BP)、分子功能(MolecularFunction,MF)以及細胞組成(CellularComponent,CC).KEGG分析數(shù)據(jù)庫是系統(tǒng)分析基因產物在細胞中的代謝途徑以
p及這些基因產物功能的數(shù)據(jù)庫,在分子和更高水平上為基因和基因組分配功能性含義[10].將2個數(shù)據(jù)集中上調基因或下調基因的交集導入DAVID68中,將結果導出至Excel表格中進行篩選,篩選標準:P<001,count≥5,將篩選后的結果導入在線工具imageGP(http://www.ehbio.com/ImageGP/index.php/
Home/Index/index.html)中,可視化分析,P<005被認為有統(tǒng)計學意義.
1.5PPI網絡的構建及關鍵基因的篩選
STRING數(shù)據(jù)庫旨在通過合并大量生物體的已知和預測的蛋白質———蛋白質關聯(lián)數(shù)據(jù)來收集和整合信息[11],其目標是建立一個全面,客觀的全球網絡,包括直接(物理)和間接(功能)交互[12].使用STRING數(shù)據(jù)庫(http://stringGdb.org)分析蛋白質之間的相互作用關系,將上調基因和下調基因DEGs全部導入
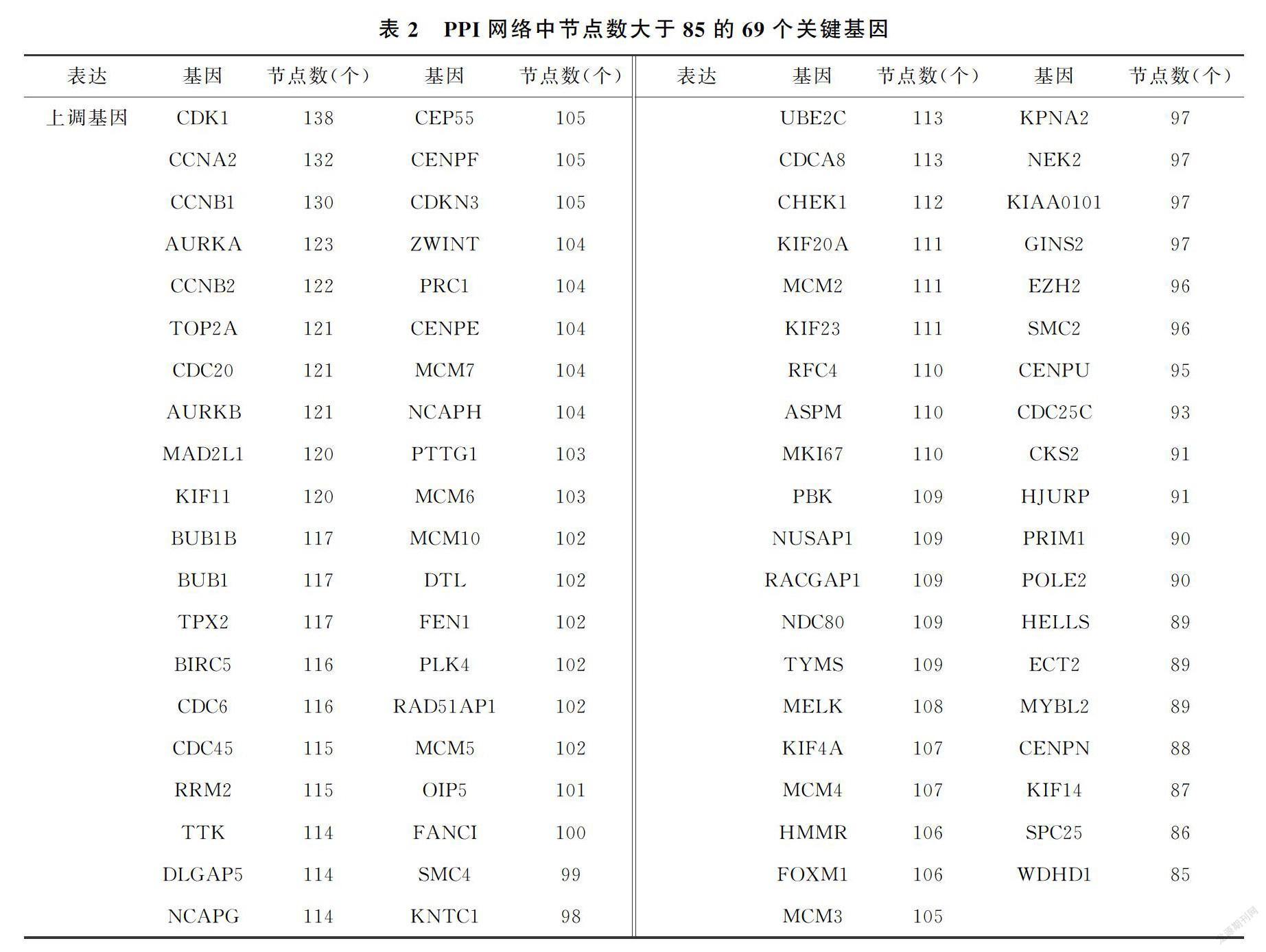
STRING數(shù)據(jù)庫,認為置信度≥04為PPI顯著.分析結果導入Cytoscape3.6.1軟件中進行可視化分析[13].利用cytoHubba插件并選擇degree[14]算法,從PPI網絡中篩選核心基因,選擇degree≥85的79個DEGs作為核心基因.
1.6關鍵基因的表達與生存預后分析
使用在線工具UALCAN[15](http://UALCAN.path.uab.edu)分析并驗證核心DEGs的表達,及KapGlanGMeierplotter(http://kmplot.com/analysis/)在線數(shù)據(jù)庫分析核心DEGs的生存預后.篩選出表達與生存預后相符合的關鍵基因,利用在線工具UALCAN分析關鍵基因的表達與HCC不同腫瘤分期關系.參數(shù)設置為默認,P<005被認為有統(tǒng)計學意義.
2結果
2.1DEGs的篩選
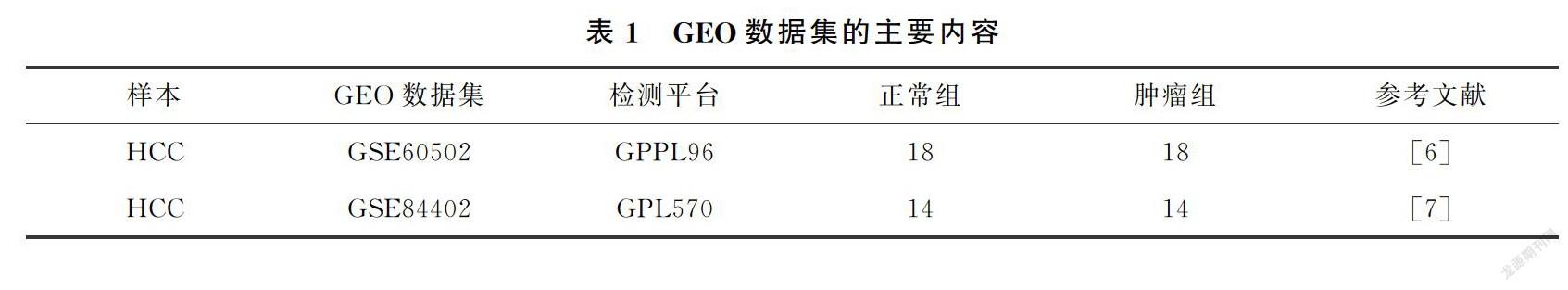
從GSE60502數(shù)據(jù)集和GSE84402數(shù)據(jù)集(表1,HCC:肝細胞癌;GEO:基因表達數(shù)據(jù)庫)中分別獲取21156和22188個基因,結果通過火山圖展示(圖1(a)),其中,紅色代表上調表達基因,綠色代表下調表達基因,黑色代表數(shù)據(jù)集中差異基因的表達水平不顯著的基因).通過Bioinformatics&EvolutionaryGenomics在線網站對GSE60502數(shù)據(jù)集和GSE84402數(shù)據(jù)集的上調基因和下調基因分別取交集,獲得這2個數(shù)據(jù)集中相同的DEGs.通過Venn圖可視化分析,發(fā)現(xiàn)這2個數(shù)據(jù)集的DEGs共同具有上調基因340個(logFC>1,P<001)和下調基因469個(logFC<-1,P<001)(圖1(b)).
2.2DEGs的GO分析和KEGG富集分析
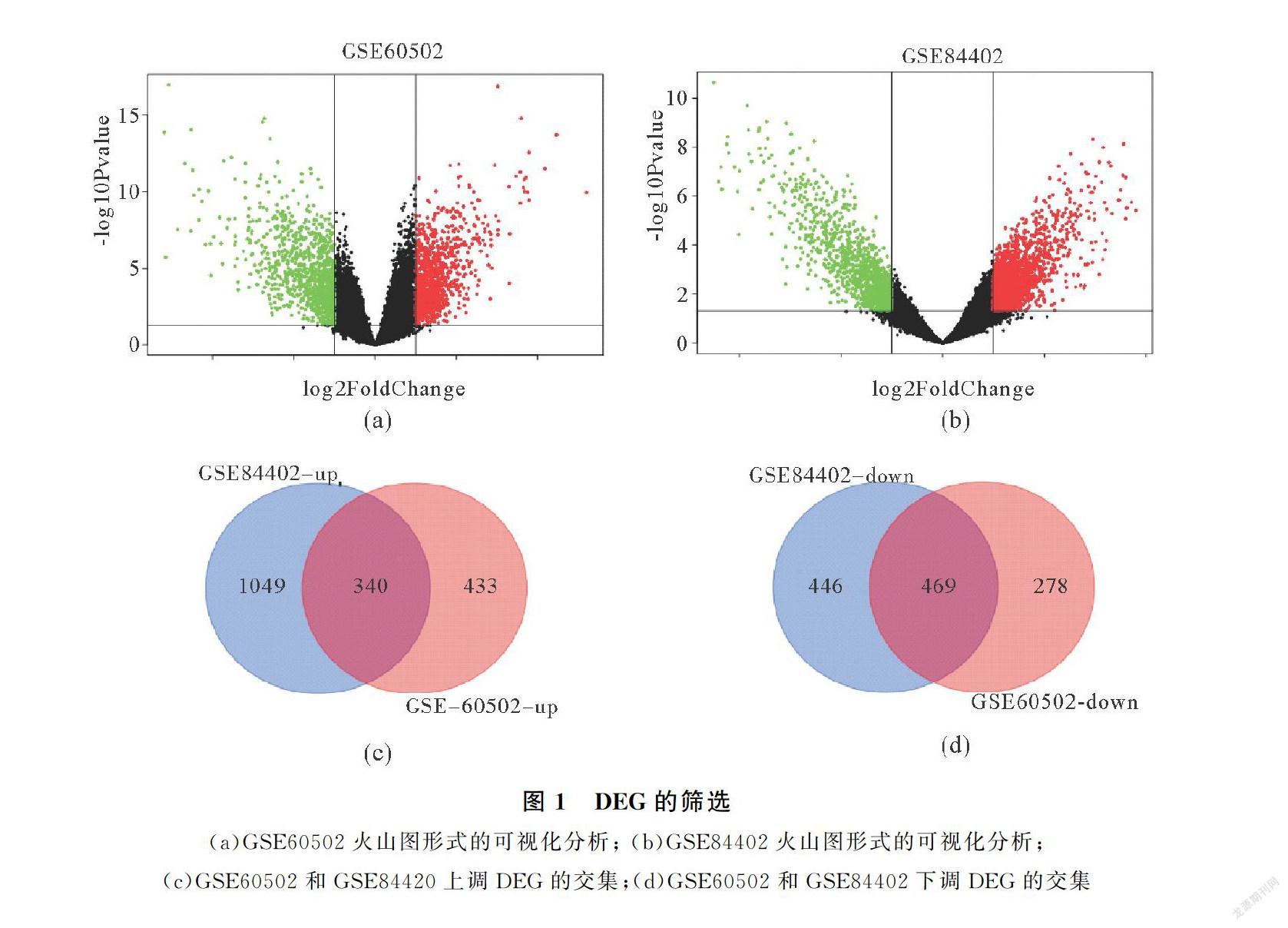
生物過程中,上調的DEGs主要參與DNA復制起始、以DNA為模板的轉錄負調控、基于微管的運動、細胞增殖以及細胞分裂過程的調控;下調的DEGs參與氧化還原過程、炎癥反應、凝血、蛋白水解以及免疫反應的調節(jié)(圖2(a)).在細胞成分上,上調的DEGs主要參與構成細胞質、細胞核、核膜、核仁以及核漿的組成成分;下調的DEGs則主要參與構成外泌體、線粒體等細胞外區(qū)域(圖2(b)).在分子功能上,上調的DEGs一般具有ATP結合能力、ATP依賴性微管運動活性、單鏈DNA結合能力以及DNA復制的起點結合能力;下調的DEGs一般具有鈣離子結合能力,鐵離子結合能力,血紅素結合能力,絲氨酸型內肽酶活性以
及與受體結合的能力(圖2(c)).在KEGG途徑上,上調的DEGs多數(shù)參與細胞周期、DNA復制、卵母細胞減數(shù)分裂以及p53信號通路;下調的DEGs主要參與代謝途徑、抗生素的生物合成、補體和凝血級聯(lián)、碳代謝以及甾體激素的生物合成信號通路(圖2(d)).
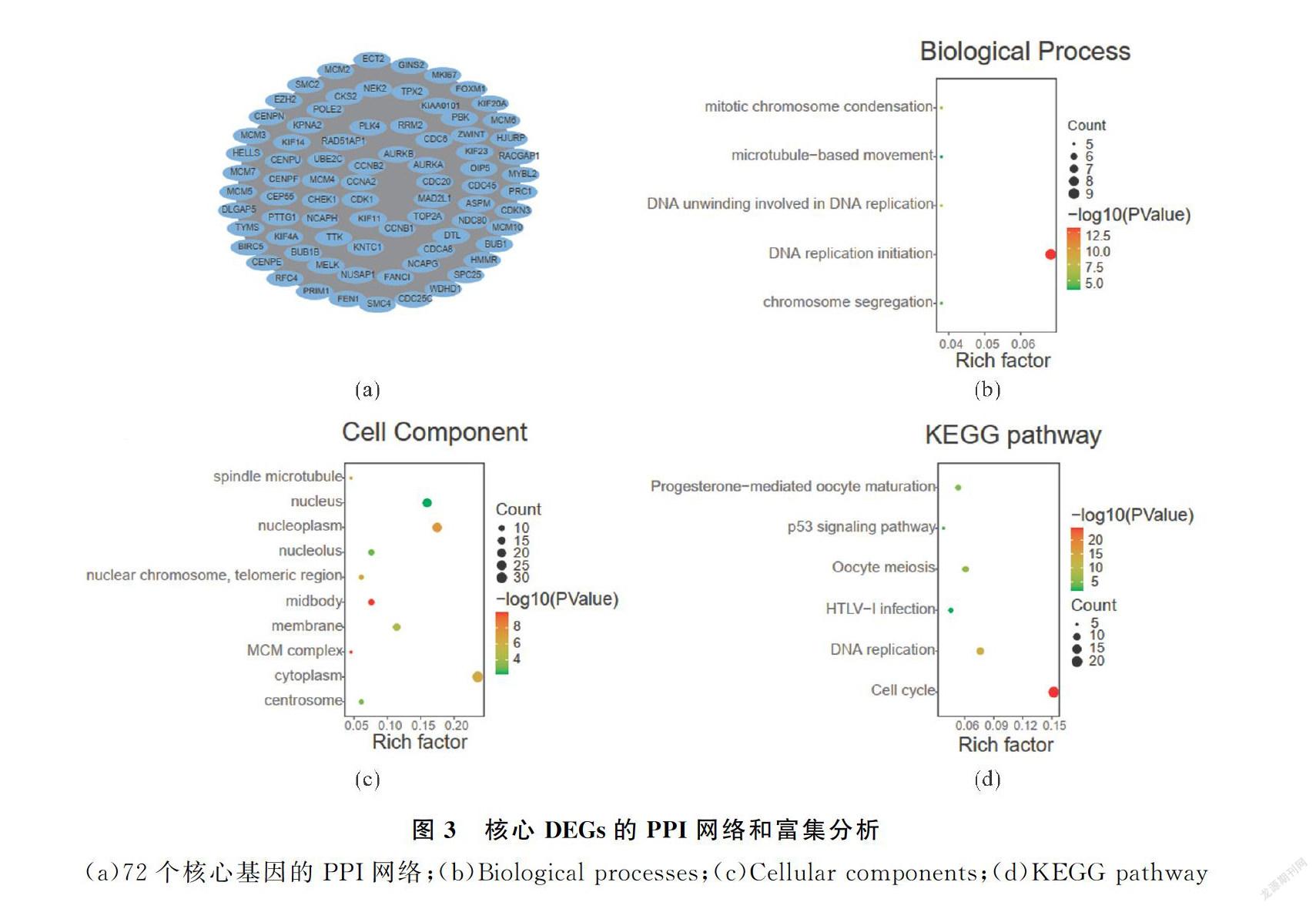
2.3PPI網絡的構建及核心基因的篩選
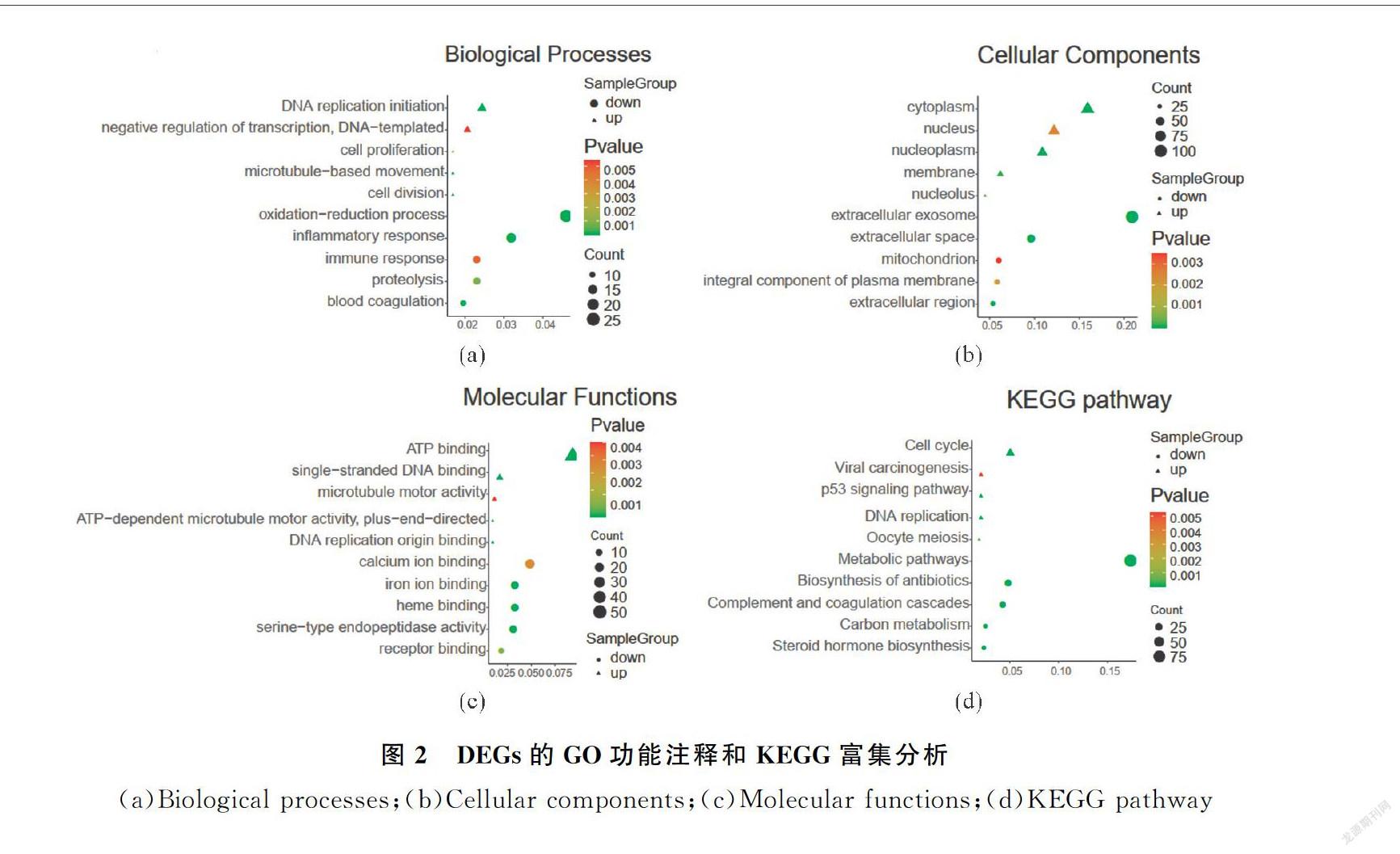
為了進一步篩選核心的DEGs,使用STRING在線數(shù)據(jù)庫分析得到2個數(shù)據(jù)集的340個上調基因和469個下調基因的蛋白調控網絡.通過Cytoscape軟件中的cytoHubba插件進一步篩選出79個核心基因(degree>85)(圖3(a),表2),且均為上調基因.接下來對這79個核心DGEs進行GO分析和KEGG富集
分析,在生物學過程中,核心DEGs主要參與DNA復制起始染色體分離以及有絲分裂染色質濃縮過程(圖3(b)).在細胞成分上,核心DEGs主要構成細胞質、細胞核以及細胞膜的組成成分(圖3(c)).在分子功能上,核心DEGs一般具有ATP結合的能力.在KEGG途徑上,核心DEGs主要參與細胞周期、DNA復制以及卵母細胞減數(shù)分裂途徑(圖3(d)).
2.4關鍵基因的表達、生存預后以及與不同腫瘤分期的關系
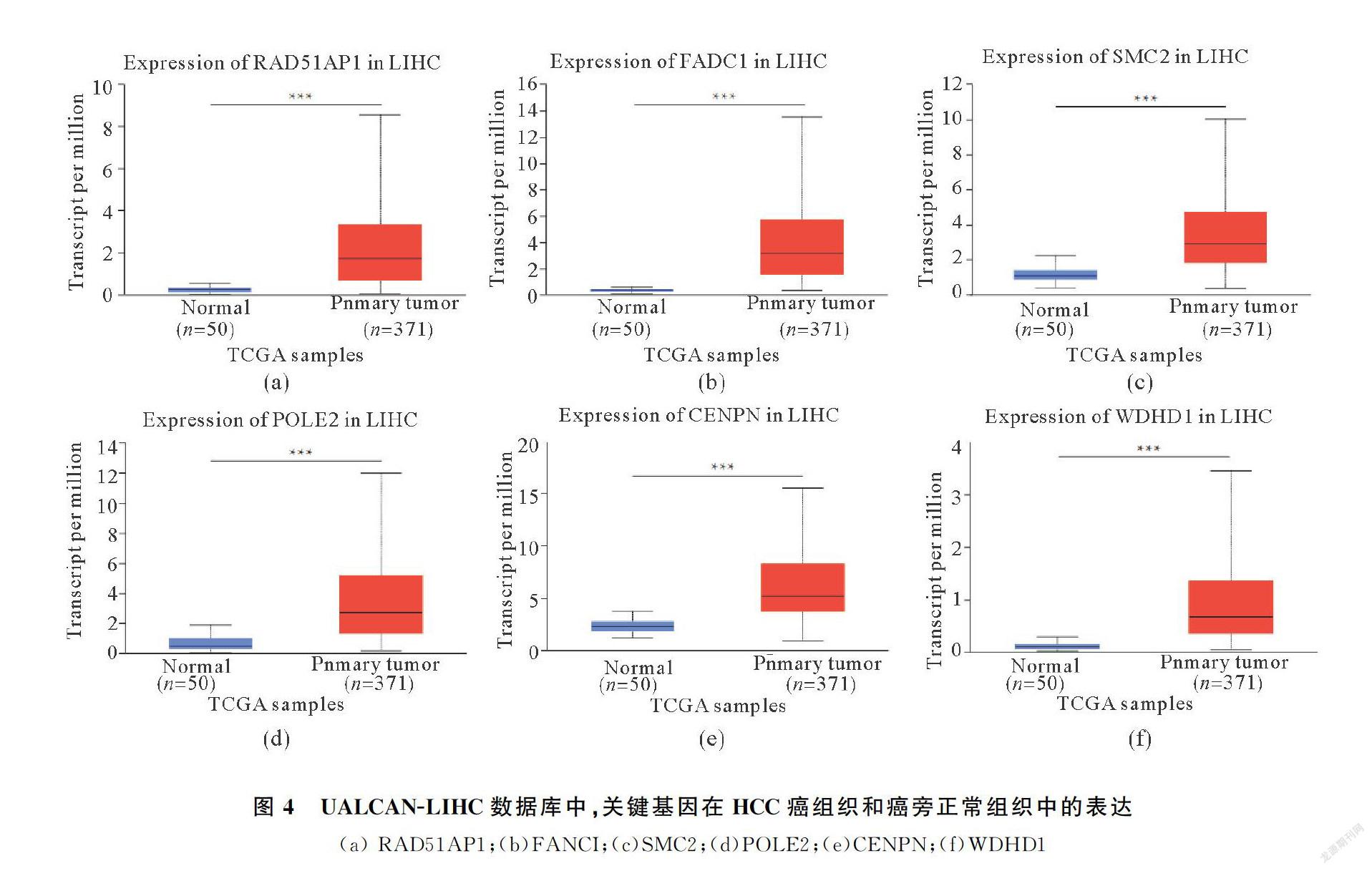
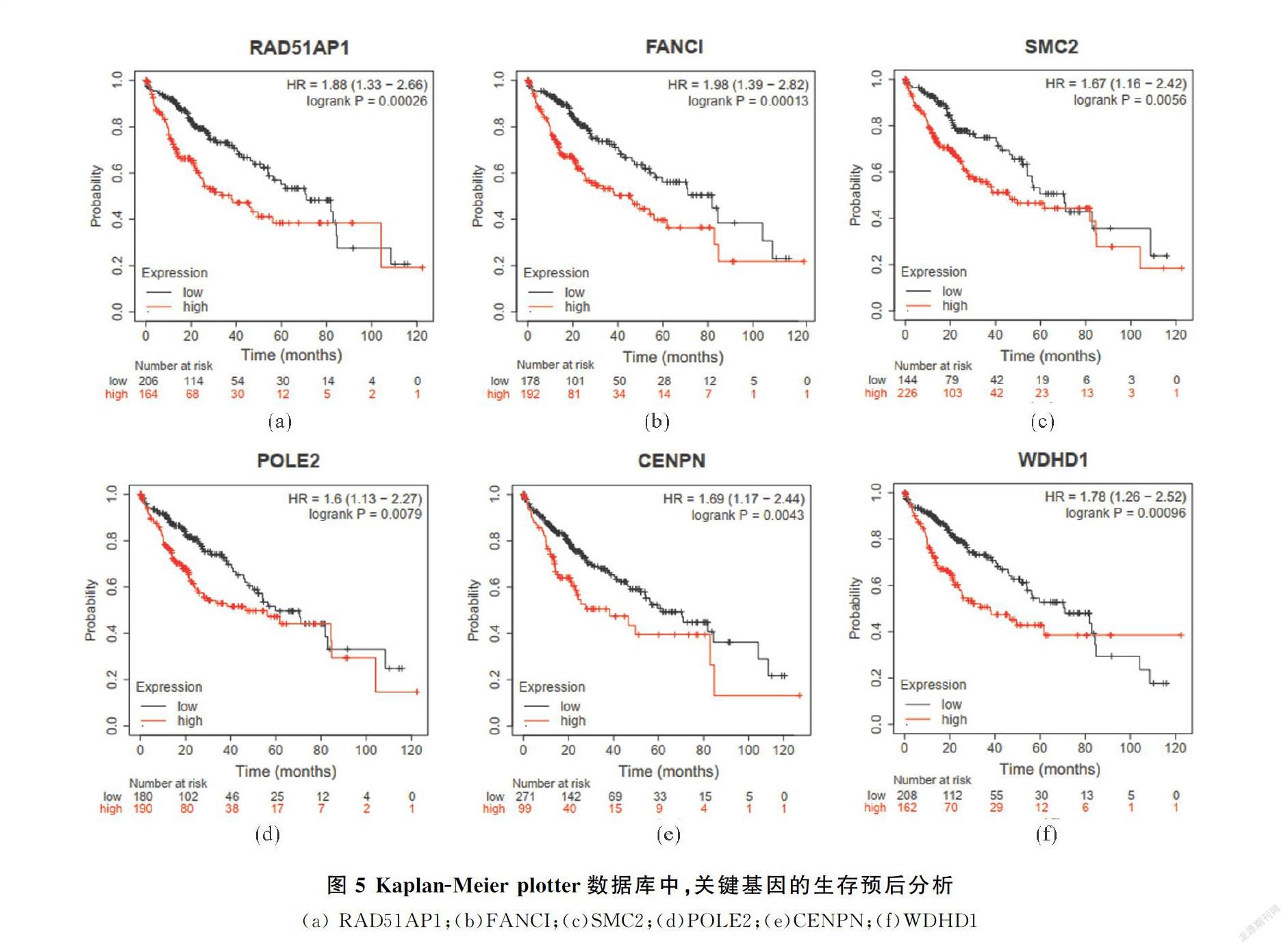
使用UALCAN以及KaplanMeierGplotter網站進行表達和生存預后分析,篩選到6個高表達的基因(圖4,?P<0.05,??P<0.01,???P<0.001),且在HCC中有顯著預后差異(圖5),分別為:
RAD51AP1(r=1.88,P<0.05)、FANCI(r=1.98,P<0.05)、SMC2(r=1.67,P<0.05)、POLE2(r=1.6,P
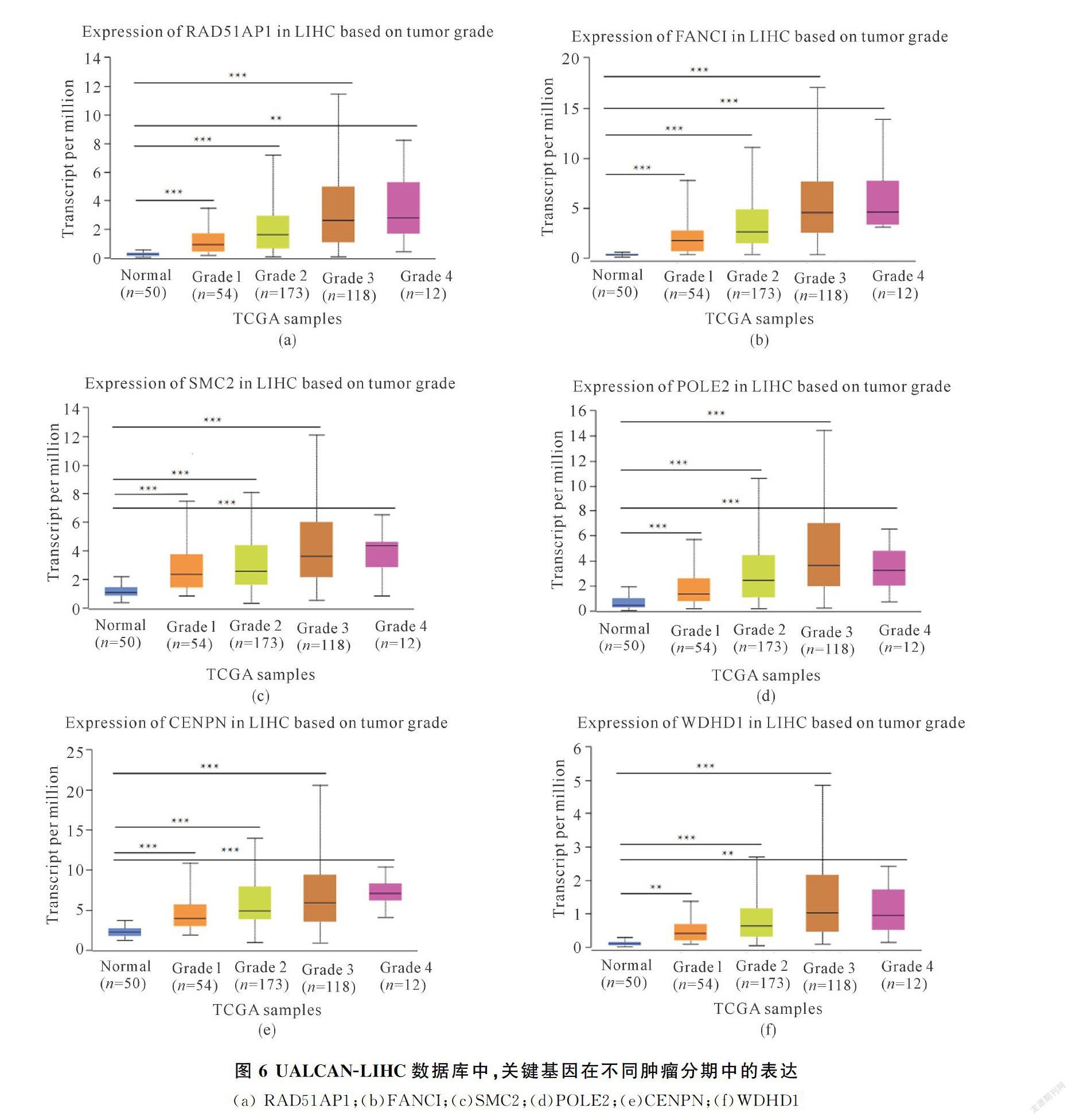
<0.05)、CENPN(r=1.69,P<0.05)、WDHD1(r=1.78,P<0.05),這些上調基因的預后生存分析結果表明,高表達的關鍵基因會顯著降低HCC患者的生存率,且與不同腫瘤分期(正常,一級,二級,三級和四級)Grade1G3呈現(xiàn)正相關趨勢(圖6,?P<0.05,??P<0.01,???P<0.001)).
3討論
隨著大數(shù)據(jù)時代正式到來,產生了大量的共享生物數(shù)據(jù),大數(shù)據(jù)應用的關鍵在于挖掘其中的重要信息并進行分析解釋[16].HCC是常見的惡性腫瘤,通常在慢性肝病的背景下出現(xiàn).手術切除和移植是早期肝細胞癌治療的基礎[17].不幸的是,肝癌患者通常被診斷為晚期[18],采用現(xiàn)代治療方法,晚期肝細胞癌患者治療選擇很少,并且預后很差,中位生存率低,HCC的全球負擔正在增加,可能會超過每年100萬例的發(fā)病率[19],因此,HCC潛在治療靶點的探究顯得格外重要.
本研究使用生物信息學的方法,對2個GEO數(shù)據(jù)集進行分析,篩選出HCC中癌組織和癌旁正常組織中的809個DEGs,包括340個上調DEGs,469個下調DEGs;GO和KEGG富集分析顯示,上調的DEGs主要作為細胞質的組成成分,與ATP結合和DNA復制過程有關,參與細胞周期的調控;下調的DEGs是細胞
外泌體的主要組成成分,與鈣離子結合和氧化還原過程有關,參與細胞代謝的調控.提示HCC的發(fā)生發(fā)展以及早期診斷和預后生存可能與調控細胞周期和DNA復制的DEGs有關.通過PPI網絡分析、表達分析以及預后生存分析進一步篩選出了與HCC進展高度相關的6個新的關鍵基因,分別為RAD51AP1、FANGCI、SMC2、POLE2、CENPN、WDHD1.RAD51AP1是端粒選擇性延長的重要介質[20],端粒延長與DNA復制和細胞周期密切相關[21G22],端粒的伸長受到細胞周期的調節(jié),并在S期與DNA復制相關[23].有報道表明,RAD51AP1是神經膠質瘤中的促癌基因[24],沉默RAD51AP1可抑制非小細胞肺癌上皮—間質轉化和轉移[25].FANCI是Akt激活的負調節(jié)劑[26],在核糖體生物發(fā)生中也起作用[27].SMC2是膀胱癌的癌基因[28],POLE2可以參與調控肺腺癌[29].CENPN是著絲粒蛋白家族的重要成員,對于動粒組裝和染色體分離至關重要,通過調控細胞周期調控口腔癌,CENPN還參與人間期細胞核質復合體的形成,WDHD1參與調控肺腺癌,并與膽管癌的上皮—間質轉化,腫瘤生長和轉移相關[30].這6個基因多數(shù)在不同腫瘤中作為癌基因發(fā)揮作用,調控細胞周期和DNA復制過程,與本研究的生物信息學分析的結果一致.
4結論
本研究通過運用生物信息學方法對2個GEO數(shù)據(jù)集GSE60502和GSE84402進行全面系統(tǒng)分析,最終篩選出6個新的可能與HCC密切相關的關鍵基因RAD51AP1、FANCI、SMC2、POLE2、CENPN、WDHD1,這些基因可能是HCC潛在的治療靶點,有望為HCC的內在機制的探究提供理論依據(jù).隨著大數(shù)據(jù)共享以及生物信息學技術的不斷進步,生物信息學在基因芯片研究的基礎上,能全面系統(tǒng)地為疾病的研究提供更可靠的理論基礎.
參考文獻
[1]STEFANODEF,CHACONE,TURCIOSL,etal.Novelbiomarkersinhepatocellularcarcinoma[J].DigLiverDis,2018;50(11):1115G1123.
[2]JIANGHY,CHENJ,XIACC,etal.Noninvasiveimagingofhepatocellularcarcinoma:Fromdiagnosistoprognosis[J].WorldJGastroGenterol,2018;24(22):2348G2362.
[3]KULIKL,ELGSERAGHB.Epidemiologyandmanagementofhepatocellularcarcinoma[J].Gastroenterology,2019;156(2):477G491.e471.
[4]李海強,趙希梅,魏賓,等.基于多特征融合和ELM的肝病多分類識別[J].青島大學學報(自然科學版),2018;31(4):42G48.
[5]LIUFF,LIUYN,CHENZ.TimG3expressionanditsroleinhepatocellularcarcinoma[J].JournalofHematologyOncology,2018,11(1):126.doi:10.1186/s13045G018G0667G4.
[6]WANGYH,CHENGTY,CHENTY,etal.Plasmalemmalvesicleassociatedprotein(PLVAP)asatherapeutictargetfortreatmentofhepatocellularcarcinoma[J].BMCCancer,2014,14:815.doi:10.1186/1471G2407G14G815
[7]WANGH,HUOX,YANGXR,etal.STAT3GmediatedupregulationoflncRNAHOXDGAS1asaceRNAfacilitateslivercancermetastaGsisbyregulatingSOX4[J].MolCancer,2017,16(1):136.doi:10.1186/s12943G017G0680G1
[8]DENNISJRG,SHERMANBT,HOSACKDA,etal.DAVID:Databaseforannotation,visualization,andintegrateddiscovery[J].GeGnomeBiol,2003,4(9):R60.
[9]KANEHISAM,F(xiàn)URUMICHIM,TANABEM,etal.Expansionofthegeneontologyknowledgebaseandresources[J].NucleicAcidsRes,2017,45(D1):D331GD338.
[10]KANEHISAM,F(xiàn)URUMICHIM,TANABEM,etal.KEGG:Newperspectivesongenomes,pathways,diseasesanddrugs[J].NucleicAcidsRes,2017,45(D1):D353GD361.
[11]SZKLARCZYKD,MORRISJH,COOKH,etal.TheSTRINGdatabasein2017:QualityGcontrolledproteinGproteinassociationnetGworks,madebroadlyaccessible[J].NucleicAcidsRes,2017,45(D1):D362GD368.
[12]SZKLARCZYKD,GABLEAL,LYOND,etal.STRINGv11:proteinGproteinassociationnetworkswithincreasedcoverage,supportingfunctionaldiscoveryingenomeGwideexperimentaldatasets[J].NucleicAcidsRes,2019,47(D1):D607GD613.
[13]DONCHEVANT,MORRISJH,GORODKINJ,etal.CytoscapestringApp:Networkanalysisandvisualizationofproteomicsdata[J].JProteomeRes,2019,18(2):623G632.
[16]易曙光,孟昕,李詠沙.基于大數(shù)據(jù)的檔案管理及挑戰(zhàn)[J].青島大學學報(自然科學版),2019;32(3):44G48.
[17]GRANDHIMS,KIMAK,RONNEKLEIVGKELLYSM,etal.Hepatocellularcarcinoma:Fromdiagnosistotreatment[J].SurgOnGcol,2016,25(2):74G85.
[18]OGUNWOBIOO,HARRICHARRANT,HUAMANJ,etal.Mechanismsofhepatocellularcarcinomaprogression[J].WorldJGastroGenterol,2019,25(19):2279G2293.
[19]LLOVETJM,MONTALR,SIAD,etal.Moleculartherapiesandprecisionmedicineforhepatocellularcarcinoma[J].NatRevClinOnGcol,2018,15(10):599G616.
[20]BARROSOGGONZLEZJ,GARCAGEXPSITOL,HOANGSM,etal.RAD51AP1isanessentialmediatorofalternativelengtheningoftelomeres[J].MolCell,2019,76(1):11G26.e17.
[21]HASEGAWAY,YAMAMOTOM,MIYAMORIJ,etal.TelomereDNAlengthGdependentregulationofDNAreplicationtimingatinGternallatereplicationorigins[J].SciRep,2019,9(1):9946.
[22]POOLELA,ZHAOR,GLICKGG,etal.SMARCAL1maintainstelomereintegrityduringDNAreplication[J].ProcNatlAcadSciUSA,2015,112(48):14864G14869.
[23]LIS.CellGcycleGdependenttelomereelongationbytelomeraseinbuddingyeast[J].BiosciRep,2011,31(3):169G177.
[24]WANGQ,TANY,F(xiàn)ANGC,etal.SingleGcellRNAGseqrevealsRAD51AP1asapotentmediatorofEGFRvIIIinhumanglioblastomas[J].Aging(AlbanyNY),2019,11(18):7707G7722.
[25]WUY,WANGH,QIAOL,etal.SilencingofRAD51AP1suppressesepithelialGmesenchymaltransitionandmetastasisinnonGsmallcelllungcancer[J].ThoracCancer,2019,10(9):1748G1763.
[26]ZHANGX,LUX,AKHTERS,etal.FANCIisanegativeregulatorofAktactivation[J].CellCycle,2016,15(8):1134G1143.[27]SONDALLESB,LONGERICHS,OGAWALM,etal.FanconianemiaproteinFANCIfunctionsinribosomebiogenesis[J].ProcNatlAcadSciUSA,2019,116(7):2561G2570.
[28]HANYH,WANY,XIONGH,etal.Structuralmaintenanceofchromosomes2isidentifiedasanoncogeneinbladdercancerinvitroandinvivo[J].Neoplasma,2020,67(2):364G370.
[29]LIJ,WANGJ,YUJ,etal.KnockdownofPOLE2expressionsuppresseslungadenocarcinomacellmalignantphenotypesinvitro[J].OnGcolRep,2018,40(5):2477G2486.
[30]LIUB,HUY,QINL,etal.MicroRNAG494GdependentWDHDIinhibitionsuppressesepithelialGesenchymaltransition,tumorgrowthandmetastasisincholangiocarcinoma[J].DigLiverDis,2019,51(3):397G411.BioinformaticsAnalysisofKeyGenesinHepatocellularCarcinoma
SUIYingGli,LUKun,F(xiàn)ULin
(InstituteofChronicDiseases,SchoolofBasicMedicine,DepartmentofMedicine,
QingdaoUniversity,Qingdao266071,China)
Abstract:HepatocellularCarcinoma(HCC)isaprimaryhepatocellularcarcinomawithhighmortality.Duetoitslimitedtreatmentandlowchemotherapeuticsensitivity,itisurgenttofindpotentialclinicaltheraGpeutictargetsandbiomarkersforprognosis.Therefore,bioinformaticsmethodwasusedtominethekeygenesintheoccurrenceanddevelopmentofHCC.DatasetsweredownloadedfromGeneExpressionOmniGbus(GEO)andDifferentiallyExpressedGenes(DEGs)werescreened.OnlinedatabaseDAVID6.8wasusedforGeneOntology(GO)enrichmentanalysisandKEGGpathwayanalysis.OnlinedatabaseSTRINGwasusedtoconstructProteinGProteinInteractionnetwork(PPI),CytoscapsoftwarewasusedtoscreencoreDEGs,andGOandKEGGenrichmentanalysiswasperformed,UALCANandKaplanGMeierplotter
onlinedatabaseswereusedtoscreenandverifytheexpressionofkeygenesandsurvivalprognosis.ThereGsultsshowedthatsixkeygenesRAD51AP1、FANCI、SMC2、POLE2、CENPN、WDHD1werescreenedout,whichweresignificantlyrelatedtotheoccurrence,developmentandsurvivalprognosisofHCC.TheymaybepotentialtherapeutictargetsforHCCandprovidetheoreticalbasisfortheexplorationoftheinternalmechanismofHCC.
Keywords:bioinformatics;differentiallyexpressedgenes;proteinGproteininteractionnetwork;hepatocelGlularcarcinoma;enrichmentanalysi
