小麥TaTDR-Like基因的克隆及表達分析
2021-09-13 01:52:08都晶晶,夏玉,任揚,李燕紅,李政,張改生,王軍衛(wèi),馬守才,宋瑜龍,齊倩,牛娜
南方農業(yè)學報 2021年9期
都晶晶,夏玉,任揚,李燕紅,李政,張改生,王軍衛(wèi),馬守才,宋瑜龍,齊倩,牛娜

摘要:【目的】克隆小麥絨氈層退化基因(Tapetum degeneration retardation,TaTDR-Like),研究其組織表達及不同育性材料間花藥不同發(fā)育時期的時空表達模式,為深入揭示小麥花藥異常發(fā)育的分子機制打下理論基礎。【方法】以普通小麥品種西農1376(MF-XN1376)為材料,通過PCR擴增克隆得到TaTDR-Like基因的3個同源拷貝,利用生物信息學分析TaTDR-Like蛋白特性及TaTDR-Like基因啟動子順式作用元件,利用實時熒光定量PCR檢測TaTDR-Like基因的組織表達情況及在可育、不育材料花藥不同時期中的時空表達模式,利用酵母雙雜交技術進行自激活檢測,并構建植物融合表達載體35S-TaTDRLD-EGFP,通過農桿菌介導轉入煙草葉片表皮細胞,觀察 TaTDRL-D蛋白亞細胞定位情況。【結果】成功克隆小麥TaTDR-Like基因,發(fā)現該基因存在3個同源拷貝即TaTDRL-A、TaTDRL-B和TaTDRL-D,分別編碼550、553、557個氨基酸,其蛋白分子量分別為58.50、58.90和59.21 kD,等電點(pI)分別為4.77、4.66和4.63。TaTDR-Like蛋白均在C端有1個bHLH保守結構域,其中TaTDRL-A與TaTDRL-B的保守結構域相似度達100%,與TaTDRL-D的保守結構域相似度為96%,TaTDRL-D與OsTDR、AtAMS保守結構域相似度分別為98%和88%;系統(tǒng)發(fā)育進化樹表明TaTDR-Like蛋白與大麥(KAE8787147.1)、二穗短柄草TDR(XP_014756384)、水稻TDR(Os02g0120500)和擬南芥AMS(AT2G16910.1)的進化關系較密切;啟動子分析表明TaTDR-Like基因啟動子處含有較多光響應元件、非生物應激反應、激素響應元件等順式作用元件;組織特異性表達分析顯示TaTDRL-D在花藥中的表達量最高,而TaTDRL-A和TaTDRL-B在幼穗表達量較高;對花藥發(fā)育不同時期表達分析顯示TaTDRL-D在花藥發(fā)育單核早期表達量最高,之后逐漸降低,單核晚期到三核期不育材料(CMS-XN1376)表達量均高于可育材料(MF-XN1376);自激活檢測結果表明TaTDRL-D具有轉錄激活活性;亞細胞定位顯示TaTDRL-D蛋白定位于細胞核。【結論】TaTDRL-D基因可能參與小麥花藥發(fā)育,負調控花藥育性。
關鍵詞: 小麥;TaTDR-Like基因;克隆;轉錄因子;表達分析
中圖分類號: S512.1? ? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2021)09-2329-10
Cloning and expression analysis of TaTDR-Like genes in wheat
DU Jing-jing1, XIA Yu1, REN Yang1, LI Yan-hong1, LI Zheng2, ZHANG Gai-sheng1,3,WANG Jun-wei1, MA Shou-cai1, SONG Yu-long1,3, QI Qian1, NIU Na1,3*
(1College of Agronomy, Northwest A & F University/National Yangling Agricultural Biotechnology Breeding Center/Yangling Branch of National Wheat Improvement Center/Engineering Research Center of Wheat Breeding of Ministry of Education, Yangling, Shaanxi? 712100, China; 2College of Life Science, Henan University/State Key Laboratory
of Crop Adversity Adaptation and Improvement, Kaifeng, Henan? 475001, China; 3Key Laboratory of
Crop Heterosis Research and Utilization in Shaanxi Province,Yangling, Shaanxi? 712100, China)
Abstract:【Objective】Wheat tapetum degeneration retardation-like(TaTDR-Like) genes were cloned and their expression patterns were analyzed between different tissues and different developmental stages of anthers in order to gain insight into the molecular mechanisms underlying abnormal wheat pollen development. 【Method】In this study, orthologues of rice (Oryza sativa L.) gene TDR(Os02t0120500) were cloned from the common wheat cultivar, MF-XN1376. Bioinformatics was used to analyze the characteristics of the proteins and cis-acting elements of the gene promoters. Moreover, real-time fluorescence quantitative PCR (qRT-PCR) was used to assess the tissue-specific expression of TaTDR-Like genes in different tissues (anther, leaf, ovary, stem and spike) and expression pattern of TaTDRL-D genes in anthers at different developmental stages. The subcellular localization of TaTDRL-D protein was observed, and yeast two-hybrid technology was used to detect its self-activation. The plant fusion expression vector 35S-TaTDRLD-EGFP was construc-ted and transferred into tobacco leaf epidermal cells through agrobacterium-mediated transfer. 【Result】It was found that there were three homologous copies, namely TaTDRL-A, TaTDRL-B and TaTDRL-D, encoding 550, 553 and 557 amino acids (aa), corresponding to molecular weights of 58.50, 58.90 and 59.21 kD and isoelectric points (pI) of 4.77, 4.66 and 4.63, respectively. TaTDR-Like proteins all have a bHLH conserved domain at the C-terminal. TaTDRL-A and TaTDRL-B had 100% and 96% similarity with TaTDRL-D, respectively, while TaTDRL-D showed 98% and 88% similarity with OsTDR and AtAMS, respectively. A phylogenetic tree analysis showed that TaTDR-Like proteins were closely rela-ted to rice TDR (Os02g0120500), Brachypodium distachyum TDR(XP014756384),barley(KAE8787147.1) and Arabidopsis AMS (AT2G16910.1). Promoter analysis results showed that TaTDRL-A, TaTDRL-B and TaTDRL-D promoters contain many cis-acting elements, such as those involved in the response to light conditions,? abiotic stress and phytohormones, which may participate in plant growth, development and the response to abiotic stress. Tissue-specific expression analysis showed that TaTDRL-D had the highest expression level in anthers, followed by young panicles and stems, while TaTDRL-A and TaTDRL-B showed higher expression levels in young panicles. Further studies revealed the expression level of TaTDRL-D to be the highest at the early mononuclear stage of anther development after which it showed a gradual decrease. The expression level in cytosolic male sterility tissues (CMS-XN1376) was higher than that in male fertile tissues (MF-XN1376) from the late mononuclear stage to the tri-nuclear stage. The results showed that TaTDRL-D had transcriptional activation activity. Subcellular localization showed that the TaTDRL-D protein was localized in the nucleus. 【Conclusion】TaTDRL-D gene may be involved in anther development and negatively regulate anther fertility in wheat.
Key words: wheat; TaTDR-Like gene; cloning; transcription factor; expression analysis
Foundation item:Shaanxi Key Research and Development Project(2019NY-003);Key Laboratory Project of Shaanxi (2018SZS-22) ;Postdoctoral Foundation of Shaanxi (2016BSHED113);College Students Innovation and Entrepreneurship Training Project of Northwest A & F University(X202010712224)
0 引言
【研究意義】雜種優(yōu)勢作為自然界的一種普遍現象,已在玉米、水稻、油菜等作物的高產和抗逆育種中取得顯著成果(王竹云等,2019;尤詩婷等,2019)。研究表明,小麥(Tritiaum aestivum L.)也具有明顯的雜種優(yōu)勢(趙昌平,2010)。小麥雄性不育是雜種優(yōu)勢利用的主要途徑,其不育機理是雜種優(yōu)勢利用的理論基礎。研究發(fā)現絨氈層異常發(fā)育會導致雄性不育,在花藥中的主要作用包括提供小孢子發(fā)育所需的營養(yǎng)物質、參與花粉外壁前體及孢粉素前體合成、分泌并轉運胼胝質酶、合成花粉包被等(樓悅,2012),表明小孢子的正常發(fā)育與絨氈層細胞是否正常發(fā)育密切相關。因此,克隆小麥絨氈層退化基因(Tapetum degeneration retardation,TaTDR-Like)并對其進行序列特征分析及組織表達分析,可為解析TaTDR-Like的功能及闡明小麥花藥發(fā)育過程中花粉異常的分子機制提供理論基礎。【前人研究進展】TDR基因屬于bHLH轉錄因子家族,大量研究表明bHLH轉錄因子家族主要參與調控植物生長發(fā)育(如光信號傳遞、植物激素信號和器官發(fā)育等)(Heisler et al.,2001;Fujimori et al.,2004;Oscar et al.,2004;廉騰飛等,2018)和逆境脅迫(干旱、低溫和鹽等)(Graciela et al.,2015;徐秀榮等,2019;Liu et al.,2020)。目前相關研究主要集中在擬南芥和水稻上,小麥中尚未見TDR基因的研究報道。在水稻中,TDR基因可通過影響花藥脂肪積累、脂質轉運和代謝等途徑引起絨氈層細胞程序性死亡(PCD),并在花藥發(fā)育過程中控制花粉壁發(fā)育(Li et al.,2006)。此外,TDR的功能缺失還導致236個基因的表達發(fā)生變化,其中有154個基因表達上調,82個基因表達下調,表明TDR是花粉發(fā)育過程中絨氈層PCD以及其他一些基本生物學過程的主要調控因子(Zhang et al.,2008)。擬南芥AMS基因(Aborted Microspores)(AT2G16910.1)是OsTDR的同源基因,該基因屬于bHLH基因MYC亞家族的典型轉錄因子,在花粉發(fā)育早期表達,在花粉減數分裂前后達到最高,是擬南芥孢粉素生物合成、分泌和花粉壁形成的主要調控因子(Anna et al.,2003)。ams突變體表現出小孢子缺陷,缺乏孢粉素沉積,總酚類化合物和角質單體顯著減少(Xu et al.,2014)。目前已在多種植物中鑒定到AMS基因并證實其與花藥的發(fā)育及育性相關,如在瓜類中利用育性分離群體定位到該基因(LOC103498166),同時證實敗育小孢子AMS基因在雄性不育系(ms-5)和雄性可育系(HM1-1)2 mm(四分體)及5 mm(第1次花粉有絲分裂)期間的表達量存在顯著差異(Sheng et al.,2017);在辣椒(Capsicum annuum L.)中發(fā)現并擴增2種亞型:CaAMS1和CaAMS2。研究發(fā)現CaAMS在絨氈層強烈表達,CaAMS的下調導致花絲部分縮短、雄蕊皺縮不裂及花粉敗育(Guo et al.,2018)。洋蔥(Allium cepa L.)AcAMS過表達顯示轉基因株系表現為部分不育甚至完全不育,部分不育植株的果莢變短,完全不育植株果莢不僅短小而且伴隨彎曲生長,轉基因株系花的雄蕊比雌蕊更長,花藥不飽滿且無光澤,有活力的花粉粒明顯少于野生型(袁巧玲,2019)。【本研究切入點】至今鮮見有關小麥TDR基因同源拷貝克隆及該基因在花藥育性表達模式分析的研究報道。【擬解決的關鍵問題】從小麥品種西農1376中通過PCR擴增克隆得到TaTDR-Like基因的3個同源拷貝,研究TaTDR-Like基因組織表達及不同育性材料間花藥不同發(fā)育時期的時空表達模式,對其進行生物信息學分析和啟動子順式作用元件分析,從轉錄激活活性、亞細胞定位和基因表達的角度初步探討小麥TaTDR-Like基因的功能,為深入揭示小麥花藥異常發(fā)育的分子機制打下理論基礎。
1 材料與方法
1. 1 試驗材料
供試材料為普通小麥品種西農1376(MF-XN1376)和遺傳型雄性不育系西農1376(CMS-XN1376),MF-XN1376是CMS-XN1376的保持系,二者具有相同的細胞核背景。試驗在西北農林科技大學實驗站進行。于2018年10月播種,每個材料各播種10行,行長1 m,行距0.25 m,株距0.67 m。取MF-XN1376植株的花藥、莖、葉、子房和幼穗用于組織表達分析,分別取MF-XN1376和CMS-XN1376的單核早期、單核晚期、二核期和三核期的花藥用于時空表達分析。取材后所有樣品立即冷凍在液氮中,于-80 ℃保存?zhèn)溆谩?/p>
主要試劑:TRIzol和KOD FX高保真購自東洋紡(上海)生物科技有限公司,反轉試劑盒Trans-Script? One-Step gDNA Removal and cDNA Synthesis SuperMix和TransScript? Tip Green qPCR SuperMix購自北京全式金生物有限公司,pClone007 Vector Kit購自北京擎科生物科技有限公司,BM無縫克隆試劑盒購自北京博邁德生物技術有限公司。主要設備儀器:IX83-FV1200激光共聚焦熒光顯微鏡購自OLYMPUS Corporation。
1. 2 試驗方法
1. 2. 1 總RNA提取及cDNA合成 采用TRIzol法提取植物組織總RNA。利用反轉試劑盒將RNA反轉錄為cDNA。
1. 2. 2 小麥TaTDRL基因克隆 從GenBank中獲取OsTDR蛋白序列(XP_015625730.1),通過NCBI/blplants/Blast的參考基因組數據庫識別TaTDR-Like的氨基酸序列。選擇得分最高、E-val最低的預測基因作為目標基因。目的基因的CDS序列從Ensembl Plants數據庫獲得,并在NCBI數據庫中得到證實。使用Primer Premier 5.0根據TaTDR-Like的CDS序列設計擴增TaTDR-Like基因的引物(表1)。以MF-XN1376 cDNA為模板,KOD高保真酶的作用下進行PCR擴增反應。反應體系50 ?L:10×KOD-Plus-Neo PCR Buffer 5 ?L,2 mmol/L dNTPs 5 ?L,25 mmol/L MgSO4 3 ?L,上、下游引物各3 ?L,cDNA模板3 ?L,ddH2O 28 ?L。擴增程序:94 ℃預變性2 min;98 ℃ 10 s,64 ℃ 30 s,68 ℃ 1 min 30 s,進行44個循環(huán);68 ℃延伸7 min。將PCR產物連接pClone007載體,轉化至DH5α感受態(tài)細胞,在相應抗性LB培養(yǎng)基上篩選出若干個獨立的克隆,提取質粒經PCR鑒定后委托北京擎科生物科技有限公司測序。通過DNAMAN軟件將測序結果與TaTDRL序列進行比對。
1. 2. 3 生物信息學分析 利用在線網站(FGENESH-HMM-baseg gene structure prediction)將TaTDR-Like翻譯為蛋白序列,利用在線工具ExPASy(https://web.expasy.org/protparam/)預測蛋白分子量和等電點。利用在線工具SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)預測TaTDR-Like蛋白的二級結構;采用SMART(http://smart.embl-heidelberg.de/)網站在線預測TaTDR-Like蛋白的保守結構域;kinasephos在線軟件進行磷酸化位點預測(http://kinasephos.mbc.nctu.edu.tw/predict.php);使用ClustalX對TaTDRL、OsTDR和AtAMS進行多重序列比對獲得logo圖;使用MEGA 6.0進行系統(tǒng)發(fā)育進化樹分析;使用在線工具ProtScale(http://www.gravy-calculator.de/)預測親水性平均數(GRAVY);在Plant-mPLoc網站(http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/)進行TaTDR-Like的亞細胞定位分析。
1. 2. 4 啟動子分析 從EnsemblPlants(http://plants.ensembl.org/index.html)數據庫中獲得上游2000 bp的TaTDRL啟動子區(qū)域序列。使用PlantCARE(https://www.dna.affrc.go.jp/PLACE/?action=newplace)和NewPLACE(http://bioinformatics.psb.ugent.be/web-tools/plantcare/html/)數據庫分析TaTDR-Like啟動子區(qū)域的調控元件。
1. 2. 5 熒光定量PCR 使用INTEGRATED DNA TECHNOLOGIES網站(https://sg.idtdna.com/Scitools/Applications/RealTimePCR/?tdsourcetag=s_pctim_aiomsg)根據CDS序列設計qRT-PCR引物(表1)。分別以不同組織、不同時期花藥cDNA為模板,以小麥TaActin為內參基因,按照全式金熒光定量說明書建立實時熒光定量PCR反應體系及程序,進行3次生物學重復,采用 2-△△Ct法計算基因相對表達量。
1. 2. 6 酵母雙雜自激活檢測 分別利用TaTDRL-D-BD-F/-R、(1~294)-BD-F/-R、(295~344)-BD-F/-R為引物擴增TaTDRL-D的CDS全長(1~344)以及截取的N端(1~294)、bHLH結構域(295~344),分別連接pGBKT7(酶切位點為EcoRⅠ和SalⅠ)載體,測序正確后,將這些構建體分別轉化酵母菌株Y2H。在SD/-Trp培養(yǎng)基挑選單克隆,將菌落稀釋100、10-1和10-2接種至SD/-Trp-His-Ade培養(yǎng)基中再培養(yǎng)3 d。
1. 2. 7 亞細胞定位 以TaTDRL-D-EGFP-F/-R為引物擴增TaTDRL-D與EGFP的融合序列。利用無縫克隆Mix(北京博邁德生物技術有限公司)構建35S-TaTDRLD-EGFP融合載體,將構建成功的質粒轉至農桿菌GV3101感受態(tài)細胞,挑取PCR檢測正確的菌株擴大培養(yǎng),采用注射法將重懸農桿菌侵染入本氏煙葉片中,48 h后將注射孔附近的葉片制作成玻片,用激光共聚焦熒光顯微鏡觀察本氏煙草表皮細胞內的綠色熒光信號,明確融合蛋白的定位情況。
2 結果與分析
2. 1 TaTDRL基因的克隆及序列分析結果
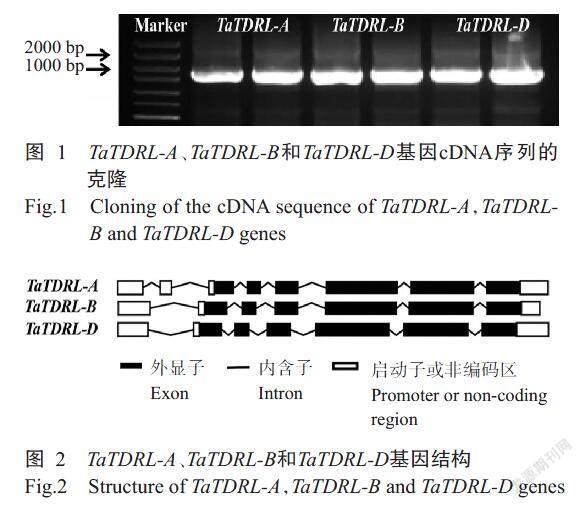
以小麥品種MF-XN1376為材料獲得cDNA第一鏈,并以此為模板進行PCR擴增獲得TaTDRL-A(TraesCS6A02G071300.1)、TaTDRL-B(TraesCS6B02 G095600.1)和TaTDRL-D(TraesCS6D02G069300.1)3個同源拷貝基因片段,瓊脂糖凝膠電泳顯示與目標條帶大小一致(圖1)。對擴增條帶進行測序和分析,結果顯示TaTDRL-A、TaTDRL-B和TaTDRL-D其完整的CDS長度分別為1653、1662和1674 bp,分別編碼550、553和557個氨基酸。對TaTDR-Like進行數據庫比對(圖2),發(fā)現TaTDRL-A、TaTDRL-B和TaTDRL-D基因全長分別為2153、2059和2150 bp,其中TaTDRL-A的基因結構含有8個外顯子和7個內含子,5'和3'未翻譯區(qū)域(UTRs)分別為301和199 bp;TaTDRL-B的基因結構含有7個外顯子和6個內含子,5'和3'未翻譯區(qū)域(UTRs)分別為130和267 bp;TaTDRL-D的基因結構含有7個外顯子和6個內含子,5'和3'未翻譯區(qū)域(UTRs)分別為251和225 bp。
2. 2 TaTDRL蛋白的生物學分析結果
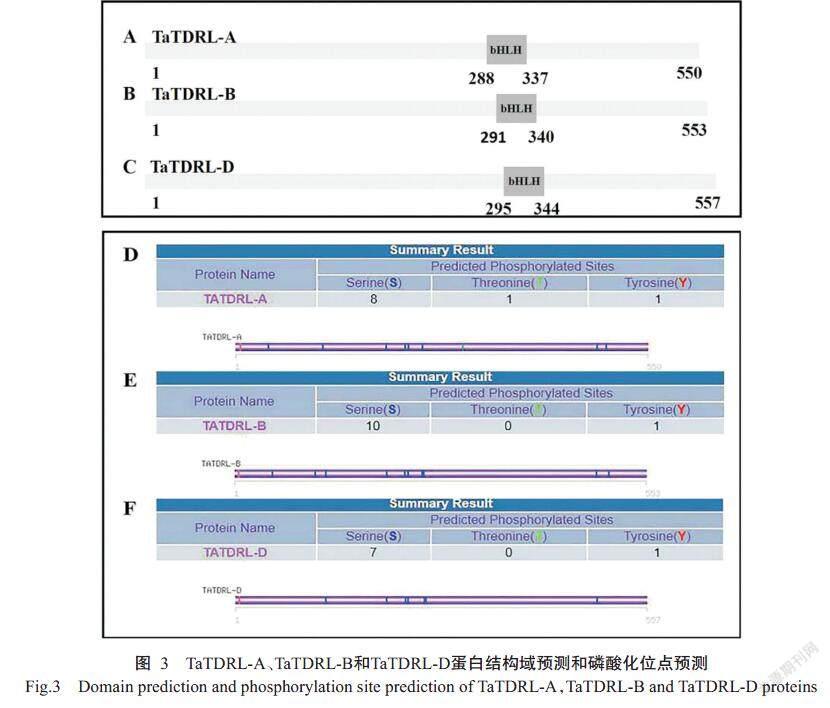
利用在線網站ExPASy預測TaTDRL-A、TaTDRL-B和TaTDRL-D蛋白分子量分別為58.50、58.90和59.21 kD,等電點(pI)分別為4.77、4.66和4.63。SMART在線軟件進行保守結構域預測發(fā)現TaTDRL-A、TaTDRL-B和TaTDRL-D均在C端含有1個bHLH保守結構域,分別跨越288~337、291~340和295~344 個氨基酸(圖3-A、3-B和3-C)。利用SOPMA對蛋白二級結構進行預測,結果顯示TaTDRL-A含有35.82%的α-螺旋、49.64%的隨機螺旋、11.09%的延伸鏈和3.45%的β-轉角。TaTDRL-B蛋白含有35.08%的α-螺旋、49.91%的隨機螺旋、10.13%的延伸鏈和4.88%的β-轉角。TaTDRL-D蛋白含有36.62%的α-螺旋、50.09%的隨機螺旋、9.16%的延伸鏈和4.13%的β-螺旋。ExPASy和Protscale在線網站預測TaTDRL-A、TaTDRL-B和TaTDRL-D的GRAVY值分別為 -0.472、 -0.513和-0.495,均為親水蛋白。利用在線軟件kina-sephos進行磷酸化分析,結果表明TaTDR-Like蛋白含有較多磷酸化位點,其中TaTDRL-A有3種類型共10個磷酸化位點,包含8個絲氨酸磷酸化位點、1個蘇氨酸磷酸化位點和1個酪氨酸磷酸化位點(圖3-D);TaTDRL-B有2種類型共11個磷酸化位點,包含10個絲氨酸磷酸化位點和1個酪氨酸磷酸化位點(圖3-E);TaTDRL-D有2種共8個磷酸化位點,包含7個絲氨酸磷酸化位點和1個酪氨酸磷酸化位點(圖3-F)。
為進一步分析TaTDRL基因功能,對TaTDRL-A、TaTDRL-B和TaTDRL-D蛋白進行在線BLASTp分析(圖4),結果表明,TaTDR-Like 3個同源拷貝間蛋白同源性較高,保守結構域基本相同,其中TaTDRL-A與TaTDRL-B的保守結構域相似度達100%,與TaTDRL-D的保守結構域相似度為96%,TaTDRL-D與OsTDR、AtAMS保守結構域相似度分別為98%和88%。與其他植物同源蛋白序列比對,發(fā)現TaTDR-Like結構域與轉錄因子AtAMS及OsTDR相似。在此基礎上,構建系統(tǒng)發(fā)育進化樹(圖5),其中AtPIF1(AT2G20180)屬于外源基因,是光敏色素作用因子(Qiu et al.,2020),結果表明TaTDR-Like同源基因聚類為同一簇,與AtPIF1親緣關系較遠。TaTDR-Like與大麥TDR(KAE8787147.1)進化親緣關系最近,與二穗短柄草TDR(XP_014756384)、OsTDR和AtAMS親緣關系較近。表明TaTDR-Like蛋白序列在進化上具有較高的保守性。
2. 3 TaTDR-Like基因啟動子分析結果
對TaTDR-Like基因啟動子順式作用元件進行分析,結果表明在TaTDRL基因啟動子上包含若干順式作用元件,包括參與植物激素響應元件、非生物應激反應和光響應的順式作用元件。對具有相同或類似功能的不同順式作用元件進行分類和統(tǒng)計,結果見圖6。TaTDR-Like基因啟動子序列含有較多的光響應元件,共計11種類型19個,并且不同成員的光反應元件種類和數量各不相同,其中TaTDRL-A、TaTDRL-B和TaTDRL-D的光反應元件數量分別為7、2和10個,說明不同染色體組的TaTDR-Like基因對光的響應程度可能不同。同時,TaTDR-Like基因啟動子序列含有許多激素響應元件,包括脫落酸、生長素、赤霉素和茉莉酸甲酯等順式作用元件,并且不同染色體組均含有脫落酸和茉莉酸甲酯的順式作用元件,其中TaTDRL-A缺少赤霉素響應元件,TaTDRL-B缺少生長素響應元件。表明TaTDR-Like基因的表達調控可能受多種激素調控,并且不同組成員間可能具有相互補充和促進作用。此外,對TaTDR-Like基因啟動子分析發(fā)現含有較多不同類型的逆境響應元件,如厭氧誘導ARE、低溫響應元件LTR及干旱相關的元件MBS等,說明該基因可能響應了多種逆境脅迫。綜合分析發(fā)現,小麥TaTDR-Like基因可能在植物生長發(fā)育及逆境響應過程中發(fā)揮重要作用。
2. 4 TaTDR-Like基因的表達模式分析結果
為分析TaTDR-Like基因在小麥不同組織中的表達模式,以小麥品種MF-XN1376的花藥、莖、葉、子房和幼穗cDNA為模板進行實時熒光定量PCR分析,結果(圖7)表明,TaTDRL在各組織內均有表達,但表達量存在明顯差異,其中,TaTDRL-A在幼穗表達量最高,在花藥和葉中表達量較高,在子房中表達最低;TaTDRL-B在幼穗表達量最高,在其他組織中表達量無明顯差異;TaTDRL-D在花藥中的表達量最高,在其他組織中表達量均較低。說明TaTDRL可能參與花藥的發(fā)育,而TaTDRL-D在花藥中高表達,推測TaTDRL-D在參與花藥發(fā)育方面起主要作用。
為進一步探究TaTDRL-D是否影響花藥育性,分別以可育材料MF-XN137和遺傳型不育材料CMS-XN1376花藥的單核早期、單核晚期、二核期及三核期的cDNA為模板進行實時熒光定量PCR分析,結果(圖8)表明,TaTDRL-D在單核早期中的表達量最高,但MF-XN1376與CMS-XN1376間無明顯差異;之后表達量逐漸降低,在可育和不育材料間呈現表達量差異,并且不育材料CMS-XN1376始終高于可育材料MF-XN1376。
2. 5 自激活檢測結果
為研究TaTDRL-D是否具有轉錄激活活性,分別截取TaTDR-Like的CDS以及N端(1~294)、bHLH結構域(295~344)3個區(qū)段,并被融合到GAL4的DNA結合結構域(GAL4-BD)上,隨后將重組質粒轉化至酵母菌株Y2H中,結果顯示TaTDRL-BD載體和N端-BD載體轉化的酵母細胞中可檢測到報告基因HIS3和ADE2的表達,表明TaTDRL-D具有轉錄激活活性(圖9)。
2. 6 亞細胞定位
使用在線軟件預測TaTDRL-D定位于細胞核。為進一步驗證該結果,利用農桿菌介導法將35S-TaTDRLD-EGFP表達載體轉化到煙草葉片表皮細胞中,瞬時表達結果(圖10)顯示空載信號飽滿,細胞膜和細胞核均有表達,而融合蛋白TaTDRLD-EGFP定位在細胞核中,與軟件預測結果一致。
3 討論
絨氈層細胞作為一種營養(yǎng)組織,與發(fā)育中的配子體直接接觸,減數分裂后PCD促進絨氈層解體被認為是小孢子形成、釋放和隨后花粉成熟的物質信號,絨氈層提前或推遲降解導致小孢子發(fā)育異常,最終引起雄性不育(于世霞,2017)。前人研究表明AMS轉錄因子屬于典型的bHLH基因的MYC亞家族,可直接與花粉外壁發(fā)育相關基因的啟動子相結合(Anna et al.,2003)。已有研究表明,在擬南芥中AMS基因是孢粉素合成、分泌和花粉壁形成的關鍵調控因子,ams突變體表現為孢子體隱性雄性不育,純合突變植株完全沒有成熟花粉,敗育的主要原因是絨氈層過早降解引起小孢子退化,并伴隨著雄蕊花絲變短現象(Alison et al.,2017)。水稻OsTDR基因屬于AMS的同源基因,該基因優(yōu)先在絨氈層表達,促進絨氈層PCD(Li et al.,2006)。AMS同源基因在辣椒、洋蔥和瓜類等植物上被證實與絨氈層發(fā)育有關(Sheng et al.,2017;Guo et al.,2018;袁巧玲,2019)。綜上所見,TDR轉錄因子在絨氈層的發(fā)育中發(fā)揮了重要作用,但在小麥上未見相關報道。本研究從西農1376中克隆得到小麥TaTDR-Like基因,發(fā)現具有3個同源拷貝,分別命名為TaTDRL-A、TaTDRL-B和TaTDRL-D。蛋白結構預測TaTDR-Like的同源拷貝均含有1個bHLH轉錄因子的保守結構域。序列分析結果表明TaTDR-Like保守結構域與OsTDR及AtAMS結構域序列高度同源,其中TaTDRL-A與TaTDRL-B的保守結構域相似度達100%,與TaTDRL-D保守結構域的相似度為96%,TaTDRL-D與OsTDR、AtAMS保守結構域相似度分別為98%和88%,推測TaTDR-Like與OsTDR、AtAMS具有相似的生理功能。此外,系統(tǒng)發(fā)育進化樹結果顯示TaTDR-Like蛋白與OsTDR、大麥(AKE8787147)的進化關系較密切,同時TDR與其他物種的同源蛋白聚類為一簇,與AtPIF1(外源基因)聚類遠,諸多研究證實TDR與花粉外壁的合成和絨氈層降解有關(Li et al.,2006;Zhang et al.,2008;Xu et al.,2014;Guo et al.,2018;袁巧玲,2019),表明TDR功能十分保守,可能在物種分化之前就具有調控花藥發(fā)育的相關功能。亞細胞定位顯示TaTDRL-D蛋白定位在細胞核內并證明其具有轉錄激活活性,說明其具有轉錄因子的基本特性,為后續(xù)進一步尋找下游靶基因打下基礎。
啟動子分析結果表明,TaTDR-Like基因啟動子上游有較多激素響應元件、光響應元件和逆境響應元件,推測TaTDR-Like基因在植物生長發(fā)育及逆境響應過程中可能發(fā)揮重要作用。由于不同的同源拷貝間順式作用元件的類型和數量各異,推測由于小麥基因組龐大且復雜,不同的同源拷貝間可能存在相互補充或促進作用。在擬南芥上有研究發(fā)現CLAVATA3/ESR-RELATED19(CLE19)小肽負調控AMS基因的表達,這種細胞外配體及其受體樣蛋白激酶(RLK)介導的信號通路在花藥發(fā)育調控中發(fā)揮著重要作用(Wang et al.,2017)。本研究在TaTDR-Like蛋白上預測到較多的磷酸化位點,推測TaTDR-Like蛋白可能與某種激酶互作并受該激酶磷酸化,從而影響TaTDR-Like的功能。
本研究通過組織表達分析發(fā)現TaTDR-Like基因在不同組織中表達量差異明顯,TaTDRL-A和TaTDRL-B在花藥幾乎不表達,而TaTDRL-D在花藥表達量最高,說明TaTDRL-D可能參與花藥發(fā)育過程,并起著重要作用。進一步驗證TaTDRL-D在花藥各個時期的表達模式,發(fā)現TaTDRL-D在可育材料MF-XN1376和遺傳型不育CMS-XN1376的單核晚期開始出現表達量的差異,之后逐漸降低,三核期表達量最低,且不育材料CMS-XN1376表達量高于可育材料MF-XN1376。前人研究發(fā)現AMS和TDR基因在花藥發(fā)育早期表達量達到峰值并起作用,之后逐漸減小(Fu et al.,2014;Alison et al.,2017),TaTDRL-D同樣驗證了這一趨勢。有研究表明遺傳性雄性不育CMS-XN1376在單核晚期開始出現敗育,三核期敗育完全(Wang et al.,2015),敗育時期與本研究TaTDRL-D表達量差異相吻合,這些結果均證明TaTDRL-D基因可能影響小孢子的育性,而關于TaTDRL-D在小麥花藥發(fā)育中的作用有待進一步驗證。
4 結論
TaTDRL-D基因可能參與小麥花藥發(fā)育,負調控花藥育性。
參考文獻:
廉騰飛,徐永萍,李蘭芬,蘇曉東. 2018. 植物激素茉莉酸通路中核心轉錄因子MYC2的結構與功能研究[C]//中國生物化學與分子生物學會第十二屆全國會員代表大會暨2018年全國學術會議摘要集. 上海:中國生物化學與分子生物學會:101. [Lian T F,Xu Y P,Li L F,Su X D. 2018. Study on the structure and function of MYC2,a key transcription factor in plant hormone jasmonate pathway[C]//Abstracts of the 12th National congress and 2018 national conference of Chinese Society of Biochemistry and Molecular Biology. Shanghai:Chinese Society of Biochemistry and Molecular Biology:101.]
樓悅. 2012. 擬南芥花藥絨氈層發(fā)育轉錄調控通路及關鍵基因TDF1功能研究[D]. 上海:上海師范大學. [Lou Y. 2012. A genetic pathway for tapetum development of anther in Arabidopsis and function analysis of defective in tapetal development and function1(TDF1)[D]. Shanghai:Shanghai Normal University.]
王竹云,張耀文,趙小光,侯君利,關周博,李殿榮,史文青. 2019. 限制油菜高產水平提高的因素解析及解決途徑[J]. 江西農業(yè)學報,31(6):45-51. [Wang Z Y,Zhang Y W,Zhao X G,Hou J L,Guan Z B,Li D R,Shi W Q. 2019. Analysis and solution of factors restricting high yield of rapeseed[J]. Acta Agriculturae Jiangxi,31(6):45-51.] doi:10.19386/j.cnki.jxnyxb.2019.06.08.
徐秀榮,楊克彬,王思寧,高志民. 2019. 毛竹bHLH轉錄因子的鑒定及其在干旱和鹽脅迫條件下的表達分析[J]. 植物科學學報,37(5):610-620. [Xu X R,Yang K B,Wang S N,Gao Z M. 2019. Identification of bHLH transcription factors in moso bamboo(Phyllostachys edulis) and their expression analysis under drought and salt stress[J]. Plant Science Journal,37(5):610-620.] doi:10.11913/PSJ. 2095-0837.2019.50610.
尤詩婷,鄧策,李會敏,呂蒙,李志敏,劉惠芬. 2019. 玉米株高和穗位高的QTL定位[J]. 河南農業(yè)科學,48(6):20-25. [You S T,Deng C,Li H M,Lü M,Li Z M,Liu H F. 2019. QTL mapping of plant height and ear height in maize[J]. Journal of Henan Agricultural Sciences,48(6):20-25.] doi:10.15933/j.cnki.1004-3268.2019.06.004.
于世霞. 2017. 煙草和番茄中活性氧介導的絨氈層程序性細胞死亡影響花粉的發(fā)育[D]. 泰安:山東農業(yè)大學. [Yu S X. 2017. Reactive oxygen species mediated tapetal programmed cell death affects pollen development in toba-cco and tomato[D]. Taian:Shandong Agricultural University.]
袁巧玲. 2019. 洋蔥細胞質雄性不育相關基因AcAMS的克隆及其功能分析[D]. 哈爾濱:東北農業(yè)大學. [Yuan Q L. 2019. Cloning and functional analysis of AcAMS gene related to cytoplasmic male sterility in onion (Allium cepa L.)[D]. Harbin:Northeast Agricultural University.]
趙昌平. 2010. 中國雜交小麥研究現狀與趨勢[J]. 中國農業(yè)科技導報,12(2):5-8. [Zhao C P. 2010. Status and trends of hybrid wheat research in China[J]. Journal of Agricultural Science and Technology,12(2):5-8.] doi:10.3969/j.issn.1008-0864.2010.02.02.
Alison C F,Simon P,Leah R B,Yang C Y,Ferjentsikova I,King J,Zheng Y,Zhang D B,Wilson Z A. 2017. Biphasic regulation of the transcription factor ABORTED MICROSPORES(AMS) is essential for tapetum and pollen development in Arabidopsis[J]. New Phytologist,213(2):778-790. doi:10.1111/nph.14200.
Anna M S,Sandra K,Ulrike S U,Peter H,Koen D,Heinz S. 2003. The Arabidopsis ABORTED MICROSPORES(AMS) gene encodes a MYC class transcription factor[J]. The Plant Journal,33(2):413-423. doi:10.1046/j.1365-313X. 2003.01644.x.
Fu Z Z,Yu J,Cheng X W,Zong X,Xu J,Chen M J,Li Z Y,Zhang D B,Liang W Q. 2014. The rice basic helix-loop-helix transcription factor TDR INTERACTING PROTEIN2 is a central switch in early anther development[J]. Plant Cell,26(4):1512. doi:https://doi.org/10.1105/tpc.114.123745.
Fujimori T,yamashino T,Kato T,Mizuno T. 2004. Circadian-controlled basic/helix-loop-helix factor,PIL6,implicated in light-signal transduction in Arabidopsis thaliana[J]. Plant & Cell Physiology,45(8):1078-1086. doi:10.1093/pcp/pch124.
Graciela C,Feranda L,Leila S F,Maria H B Z,Márcia M P. 2015. Possible roles of basic helix-loop-helix transcription factors in adaptation to drought[J]. Plant Science an International Journal of Experimental Plant Biology,223:1-14. doi:10.1016/j.plantsci.2015.03.012.
Guo J J,Liu C,Wang P,Chen Q,Sun L,Yang W C,Shen H L. 2018. The Aborted microspores (AMS)-Like gene is required for anther and microspore development in Pe-pper(Capsicum annuum L.)[J]. International Journal of Molecular Sciences,19(5):1341-1355. doi:10.3390/ijms 19051341.
Heisler M G,Atkinson A,Bylstra Y H,Walsh R,Smyth D R. 2001. SPATULA,a gene that controls development of carpel margin tissues in Arabidopsis,encodes a bHLH protein[J]. Development,128(7):1089-1098. doi:/10.1242/ dev.128.7.1089.
Li N,Zhang D S,Liu H S,Yin C S,Li X X,Liang W Q,Yuan Z,Xu B,Chu H W,Wang J,Wen T Q,Huang H,Luo D,Ma H,Zhang D B. 2006. The rice tapetum degeneration retardation gene is required for tapetum degradation and anther development[J]. Plant Cell,18(11):2999-3014. doi:10.1105/tpc.106.044107.
Liu H,Yang Y,Liu D D,Wang X Y,Zhang L S,2020. Transcription factor TabHLH49 positively regulates dehydrin WZY2 gene expression and enhances drought stress tolerance in wheat[J]. BMC Plant Biology,20(1):259-270. doi:10.1186/s12870-020-02474-5.
Oscar L,Jose M C,Jose J,Sánchez S,Roberto S. 2004. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis[J]. Plant Cell,16(7):1938-1950. doi:10.1105/tpc.022319.
Qiu J R,Xiang X Y,Wang J T,Xu W X,Chen J,Xiao Y,Jiang C Z,Huang Z. 2020. MfPIF1 of resurrection plant myrothamnus flabellifolia plays a positive regulatory role in responding to drought and salinity stresses in Arabidopsis[J]. International Journal of Molecular Sciences,21(8):3011-3030. doi:10.3390/ijms21083011.
Sheng Y Y,Wang Y D,Jiao S Q,Jin Y Z,Ji P,Luan F S. 2017. Mapping and preliminary analysis of ABORTED MICROSPORES(AMS) as the candidate gene underlying the male sterility(MS-5) mutant in melon(Cucumis melo L.)[J]. Frontiers in Plant Science,8:902-916. doi:10. 3389/fpls.2017.00902.
Wang S P,Zhang G S,Zhang Y X,Song Q L,Chen Z,Wang J S,Guo J L,Niu N,Wang J W,Ma S C. 2015. Comparative studies of mitochondrial proteomics reveal an intimate protein network of male sterility in wheat(Triticum aestivum L.)[J]. Journal of Experimental Botany,66(20):6191-6203. doi:10.1093/jxb/erv322.
Wang S S,Lu J J,Song X F,Ren S C,You C J,Xu J,Liu C M,Ma H,Chang F. 2017. Cytological and transcriptomic analyses reveal important roles of CLE19 in pollen exine formation[J]. Plant Physiolog,175(3):1186-1202. doi:https://doi.org/10.1104/pp.17.00439.
Xu J,Ding Z W,Barrena J V,Shi J X,Liang W Q,Yuan Z,Reichhart D W,Schreiber L,Wilson Z A,Zhang D B. 2014. ABORTED MICROSPORES acts as a master regulator of pollen wall formation in Arabidopsis[J]. The Plant Cell Online,26(4):1544-1556. doi:10.1105/tpc.114.122986.
Zhang D S,Liang W Q,Yuan Z,Li N,Shi J,Wang J,Liu Y M,Yu W J,Zhang D B. 2008. Tapetum degeneration retardation is critical for aliphatic metabolism and gene re-gulation during rice pollen development[J]. Molecular Plant,1(4):599-610. doi: 10.1093/mp/ssn028.
(責任編輯 麻小燕)
