天門冬科廣義鈴蘭族的染色體數目與核型進化研究
2021-09-16 14:49:42張籍元張代貴聶澤龍
西北植物學報 2021年8期
關鍵詞:植物
張籍元,張 成,孟 然,張代貴,聶澤龍,孟 盈
(吉首大學 生物資源與環境科學學院植物資源保護與利用重點實驗室,湖南吉首 416000)
廣義鈴蘭族(Convallarieae)包括白穗花屬(Speirantha)、鈴蘭屬(Convallaria)、萬年青屬(Rhodea)、吉祥草屬(Reineckia)、長柱開口箭屬(Tupistra)和蜘蛛抱蛋屬(Aspidistra),共約200余種。鈴蘭族植物皆為多年生草本,除鈴蘭屬冬季倒苗外,其余5屬皆為常綠。鈴蘭族中僅鈴蘭屬廣布于北半球,屬于典型的北溫帶分布,其余屬集中分布于東亞和東南亞。中國和日本是鈴蘭族植物的多樣化中心,該族70%以上的特有種分布在中國和日本[1]。鈴蘭族中大多數種具有重要的觀賞和藥用價值,如吉祥草屬、萬年青屬和蜘蛛抱蛋屬等,其中蜘蛛抱蛋屬藥理活性良好,民間應用較多,其在保護心腦血管、抗菌消炎、抗腫瘤和降血糖等方面具有廣闊的應用前景[2]。
鈴蘭族傳統隸屬于百合科,主要特征為葉簇生在根狀莖上,花葶沿葉基部側生,花序為簡單的總狀花序或間斷的穗狀花序。Krause提出廣義鈴蘭族,并將百合科分為12亞科35族[3]。在Krause的百合科系統中,鈴蘭族又被分為鈴蘭亞族(Coralarinae)和蜘蛛抱蛋亞族(Aspidistrinae)[3]。Hutchinson認為鈴蘭族具根狀莖是百合科最原始的類群,于是又把Evrardiello和開口箭屬(Compylandra)2個屬納入鈴蘭族內,并將廣義鈴蘭族劃分為狹義鈴蘭族(包括鈴蘭屬、白穗花屬、吉祥草屬和夏須草屬)和蜘蛛抱蛋族(蜘蛛抱蛋屬、萬年青屬和長柱開口箭屬)。Dalgren基于形態特征的研究結果,提出將狹義鈴蘭族、蜘蛛抱蛋族、黃精族(Polygonateae)與沿階草族(Ophiopogoneae)共同組成鈴蘭科(Convallariaceae)的觀點[4]。《中國植物志》把Campylandra并入長柱開口箭屬,早期的系統發育研究通常也將夏須草屬和Comospermum歸置于鈴蘭族[5]。鈴蘭族作為一個自然的單系類群,也得到了分子系統學證據的支持,但該族的系統位置依然存在爭議,分子證據支持將它從百合科分出置于鈴蘭科中,后來將鈴蘭科擴大到假葉樹科(Ruscaceae),APG IV系統最終將該族歸為天門冬科[6]。
染色體數目與核型反映了物種在染色體水平上的整體特征,細胞學研究有助于揭示物種間的親緣關系和起源演化,為植物系統分類提供可靠的細胞學佐證[7]。國內外學者已經對鈴蘭族各屬植物的染色體進行了大量的研究工作[8-29]。對于鈴蘭族的細胞學研究始于1961年Roy發現廣東蜘蛛抱蛋(A.lurida)的染色體為2n=36[28]。中國對鈴蘭族的研究比較晚,楊滌清在1983開展了單種屬百穗花的染色體數目為2n=38[23],隨后許炳聲又報道了吉祥草的染色體核型[22]。洪德元等[8]綜合分析整個鈴蘭族各屬的染色體結構發現白穗花屬核型與蜘蛛抱蛋核型較一致,認為兩屬可能起源于一個共同祖先。楊滌清等[24]對栽培的萬年青與野生的萬年青作了核型研究,發現二者核型很接近,同時認為萬年青與開口箭屬的核型無明顯區別,萬年青的核型與鈴蘭屬也較為相似,通過對長柱開口箭(T.chinensis)等種的核型研究得出了長柱開口箭屬核型與吉祥草屬相近的結論。黃錦嶺在對長柱開口箭屬的國產種類進行了詳細研究后,發現長柱開口箭屬的核型結構在屬類變異相當大,不僅具有對稱性較強的單型性核型,也有高度不對稱的兩型性核型[9]。黃錦嶺等[12]依據該屬與其可能的近緣類群的核型相似性進行比較,推斷蜘蛛抱蛋屬植物染色體原始基數為 x =19,但這與其外部形態結構尤其是花部式樣及花粉形態演化趨勢并不相不符。王任翔等[19]的研究發現蜘蛛抱蛋屬染色體數目2n=36和2n=38比率基本相同,在染色體數目2n=36的種類中,花部式樣的形態結構較為簡單,可能代表其原始性狀;而染色體數目2n=38的種類中,花部式樣的形態結構較復雜和特化,可能代表其進化性狀。劉安莉等[16]通過研究屬內形態特征和核型進化結果同意上述觀點,由此推測蜘蛛抱蛋的核型可能是由不對稱向對稱性增強的方向演化。
關于廣義鈴蘭族內各屬的分子系統發育關系一直存在爭論,因為相對孤立,連續的變異性狀少,所以難以判斷性狀的演化方向和屬間系統發育關系。Yamashita等[29-30]基于trnK和rbcL對鈴蘭族進行了系統發育分析,結果表明狹義鈴蘭族不是單系群,支持鈴蘭族和蜘蛛抱蛋族共同構成廣義鈴蘭族,鈴蘭屬、百穗花屬和吉祥草屬處于基部位置,開口箭屬Compylandra與萬年青屬形成第二支,Tricalistraochracea與長柱開口箭屬Tupistra形成的一支與蜘蛛抱蛋屬互為姐妹關系,同時提議將開口箭屬歸為萬年青屬,Tricalistra歸入長柱開口箭屬。黃丹妮通過對65種蜘蛛抱蛋屬植物和12個外類群進行系統發育分析表明,蜘蛛抱蛋屬植物是單系類群,與外類群中的長柱開口箭屬關系最近,也支持將開口箭屬并入萬年青屬[31]。
植物染色體的數目和形態等是最穩定的細胞學特征之一,在不同的種間染色體有不同的數目、核型及大小等的特征區別,植物染色體的核型特征也是表明該種系統演化位置以及和近緣物種之間關系的重要依據。染色體數目可以體現自然進化的進程及方向,為植物進化和系統發育研究提供有價值的信息[7]。鈴蘭族具有較為穩定的染色體數目以及明顯的核型變化,是研究植物染色體進化非常理想的類群。關于鈴蘭族的研究涵蓋了形態學、分類學、細胞學、孢粉學和植物化學成分等[1, 11, 12, 29-35],但是該族屬間以及各屬下種間的系統關系一直存在爭議,本研究通過廣泛收集鈴蘭族植物的分子數據來重建系統發育樹,明確屬間以及各屬下種間的系統關系。因此本研究全面收集整理鈴蘭族染色體數目和核型,結合分子系統發育樹推測本族植物的染色體演化規律,也為鈴蘭族屬間關系及物種進化關系提供了細胞學證據。
1 材料和方法
1.1 細胞學實驗
本實驗5種鈴蘭族植物材料及采集信息見表1,憑證標本存放于湖南省湘西自治州吉首大學生物資源與環境科學學院標本館(JSU)。在天氣晴朗的上午9:00-10:00從活體上取1~2 cm長的幼嫩根尖浸泡于飽和對二氯苯水溶液中,于室溫下預處理4 h,然后蒸餾水洗凈,再用卡諾固定液I(無水乙醇∶冰醋酸 = 3∶1)于4 ℃冰箱中固定30 min,然后用1 mol·L-1HCl∶45%的冰醋酸 = 1∶1混合溶液在60 ℃的水浴鍋中解離1 min,1%的地衣紅染液染色,過夜,常規方法壓片,顯微鏡下觀察、拍照。每個物種選擇10個以上有絲分裂中期染色體分散良好的細胞拍照,計數,最后選擇最好的細胞圖用Photoshop CS6軟件上進行處理,核型分析及命名按照Levan等[36]的方法進行,核型類型按照Stebbins的標準處理[37]。此外,也用Arano提出的不對稱系數As.K%來評估核型不對稱性,As.K%值越高,核型越不對稱[38]。

表1 實驗材料來源與憑證標本
1.2 染色體數據收集與核型劃分標準
本研究對鈴蘭族植物已報道的染色體資料進行全面的收集整理及核型統計,推測鈴蘭族植物染色體非整倍化和多倍化的進化規律和地位,這對全面了解鈴蘭族及其各屬的起源和演化有積極的推動作用。染色體數據的收集主要根據《中國有文獻報道的植物染色體數目索引》和“Index to Plant Chromosome Numbers”數據庫進行查詢,前者主要包括發表在國內外雜志和期刊的資料,后者包括全世界發表的所有植物的染色體索引,并且可在網站上(http://www.tropicos.org/Project/IPCN)查詢,同時重點查閱最近分散在國內外各主要出版物上的文章等。
本研究對鈴蘭族的核型劃分標準和方法參考Yamashita等[29]觀點長度上有兩種變異模式:單型(表現為染色體間在長度上沒有明顯的差異,呈逐漸過渡的趨勢)和三型(由3組不同長度的染色體組成,并結合中期染色體的基數和長度變異模式將鈴蘭族分為:單型,染色體基數x=19的鈴蘭型;三型,基數為x = 19時為長柱開口箭型,基數為x = 18時則被稱為蜘蛛抱蛋型。
1.3 基于系統發育的染色體進化分析
基于前期鈴蘭族的分子系統學研究[29],本研究選取代表鈴蘭科各個屬和主要分支的64個種,天門冬屬Asparagus作為外類群。選取常用于該類群系統發育分析的基因片段,包括4個葉綠體基因片段(rbcL、matK、psbA-trnH和trnL-F)和1個核基因片段(ITS)進行測序分析。本研究對天門冬科10個種進行PCR及測序,DNA提取、PCR引物及反應程序和測序方法參考Wang等[39]。同時從國際生物數據庫(NCBI)下載了54個種的基因序列。所有用于本研究的類群憑證標本和GenBank登錄號等詳細信息見表2和3。分子序列首先用MUSCLE進行比對,然后用PhyED軟件手動調整[40]。預分析表明葉綠體與核基因序列的結果不存在明顯的沖突,并且與所有序列聯合的結果非常相似,故我們最終將5個片段進行串聯進行最大似然法(Maximum likelihood,ML)分析,并且不同序列片段采取單獨的進化模型。ML分析利用RAxML7.2.6進行運算,進行1 000次Bootstrap搜索,最后得到一致樹[41]。運算產生的樹文件使用軟件Figtree1.4.2進行可視化,最后把染色體數目和核型映射到系統進化樹上來探討它們的進化關系。
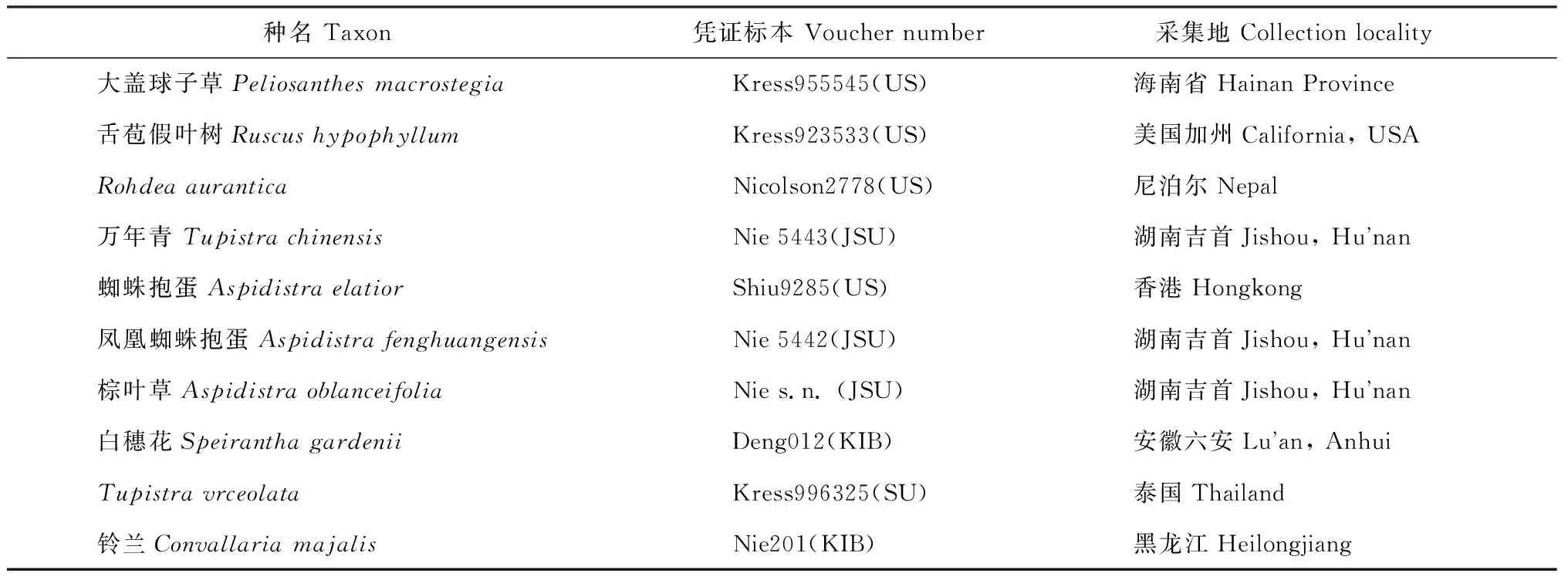
表2 實驗材料來源與憑證標本

表3 本研究從GenBank下載序列的相關信息
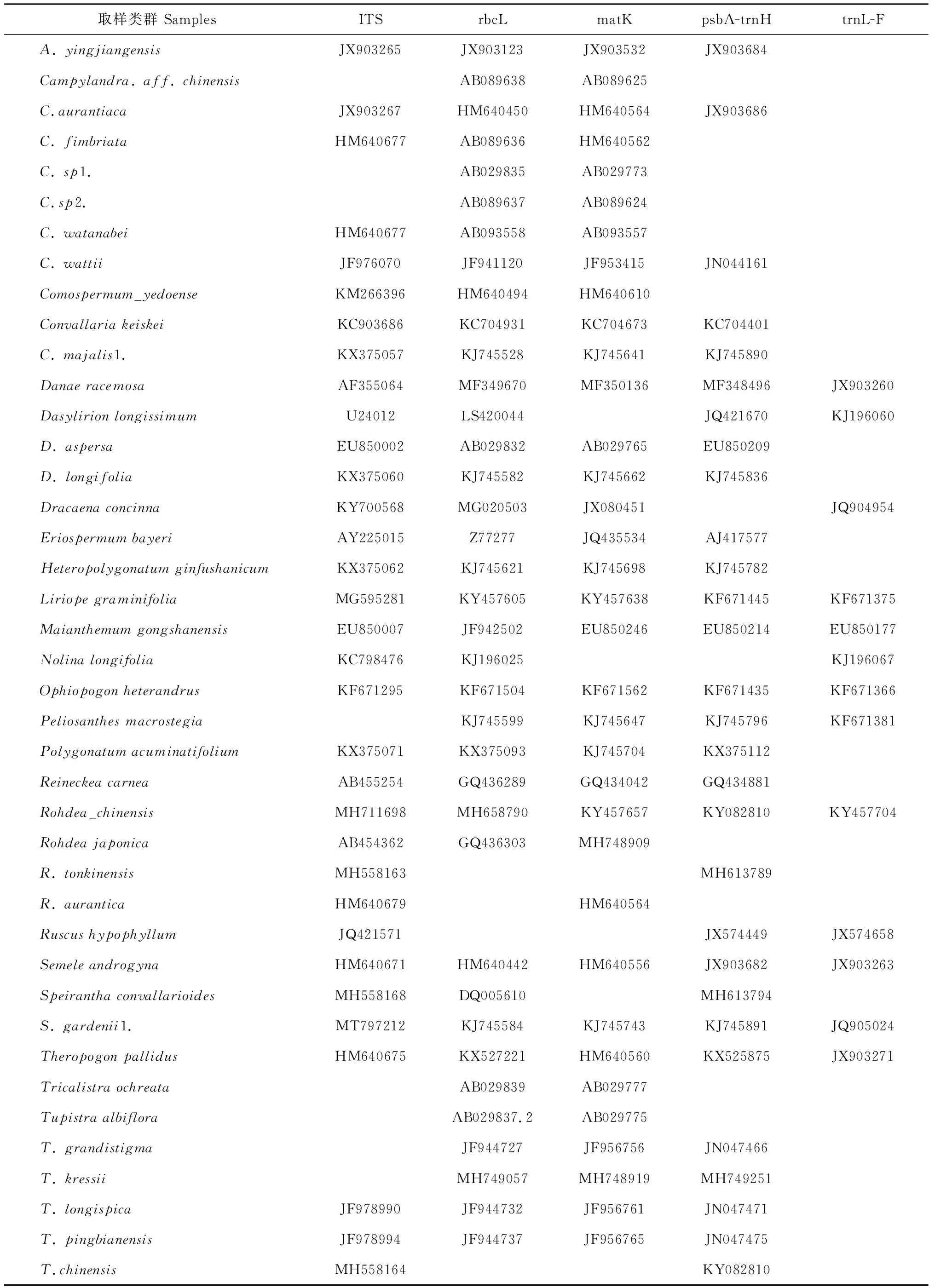
續表3 Continued Table 3
2 結果與分析
2.1 細胞學實驗觀察結果
本研究中鈴蘭族5個種植物的染色體核型公式及特征見表4,染色體形態和核型圖見圖1。結果表明這5個種中,除蜘蛛抱蛋染色體數目為2n=36外,其余4種均為2n=38,均為二倍體。這5個種中染色體均由m,sm和st的染色體構成,不對稱性核型均為Stebbins’-2C型,核型不對稱系數As.K%變化范圍為65%~69%,并沒有明顯差別。但在染色體核型公式上有一定差別。其結果分別表述如下。

表4 5種鈴蘭族植物的染色體核型公式及特征

A.蜘蛛抱蛋;B. 海南蜘蛛抱蛋;C. 四川蜘蛛抱蛋;D. 鳳凰蜘蛛抱蛋;E. 萬年青. 標尺=5 μm圖1 5種鈴蘭族植物中期染色體形態圖A.Aspidistra elatior; B. A. hainanensis; C. A. sichuanesis; D. Aspidistra fenghuangensis; E. R. japonica. Bar=5 μmFig.1 Morphology and karyotype of somatic chromosomes of 5 species from the tribe Convallarieae
2.1.1 蜘蛛抱蛋AspidistraelatiorBlume蜘蛛抱蛋的染色體數目為2n=36,染色體基數為x=18,二倍體(圖1, A)。在第1對(6.75~6.25 μm)與第2-9對(5.40~2.56 μm)之間在長度上有一個明顯的差別(0.85 μm),第9對與第10-18對(1.66~0.80 μm)也存在一個明顯的差別(0.90 μm)。核型公式為2n=36=16m+14sm+6st,最長染色體長度為6.75 μm,最短染色體長度為0.80 μm,最長染色體與最短染色體的比值為8.4,臂比小于2的染色體所占比例為61%。Stebbins’不對稱核型為2C。核型不對稱系數As.K%為67%。其核型表現為蜘蛛抱蛋型核型(三型,x=18)。蜘蛛抱蛋染色體數目有2n=36和2n=38兩種,本種不同個體或不同居群間核型不一樣,黃錦嶺報道該種染色體數目為2n=38,其核型公式為2n=38=22m+4sm(2sat)+12st,第9對染色體短臂具隨體[12]。雖然本研究的染色體數目與王任翔報道的一致,但在核型公式和隨體方面卻有所不同(2n=36=16m+4sm+12st(2sat)+4t,第10對染色體短臂具隨體),具有2對端部著絲點的染色體[19]。本研究中并沒有發現隨體染色體的存在,也沒有臂比值大于7的端部著絲點染色體(t)存在。可見,蜘蛛抱蛋在不同居群之間染色體核型和數目均有變異,是一個正在分化的類群。
2.1.2 海南蜘蛛抱蛋AspidistrahainanensisChun et F. C. How海南蜘蛛抱蛋染色體數目為2n=38,染色體基數為x=19,二倍體(圖1,B)。核型公式為2n=38=20m+12sm+8st,在第1對(7.20~6.98 μm)與第2~6對(5.81~3.96 μm)之間在長度上有一個明顯的差別(1.17 μm),第6對與第7~19對(2.89~1.10 μm)也存在一個明顯的差別(1.07 μm)。最長染色體長度為7.20 μm,最短染色體長度為1.10 μm,最長染色體與最短染色體的比值為6.54,臂比小于2的染色體所占比例為66%。Stebbins’不對稱核型為2C。長柱核型不對稱系數As.K%為69%。其核型表現為開口箭型核型(三型,x=19)。海南蜘蛛抱蛋染色體數目有2n=36和2n=38兩種報道,其中2n=38的占比為68%,該種與王任翔[19]報道的2n =36=20m(2sat)+14st+2t不一致,且我們也沒有發現t型染色體。
2.1.3 四川蜘蛛抱蛋AspidistrasichuanensisK. Y. Lang et Z. Y. Zhu四川蜘蛛抱蛋的染色體數目是2n=38,染色體基數為x=19,二倍體(圖1,C)。核型公式為2n=24m+6sm+8st,在第1對(7.48~6.95 μm)與第2-6對(6.09~3.95 μm)之間在長度上有一個明顯的差別(0.86 μm),第6對與第7~19對(2.92~0.86 μm)也存在一個明顯的差別(1.03 μm)。最長染色體長度為7.48 μm,最短染色體長度為0.86 μm,最長染色體與最短染色體的比值為8.7,臂比小于2的染色體所占比例為71%。Stebbins’不對稱核型為2C。核型不對稱系數As.K%為65%。其核型表現為長柱開口箭型核型(三型,x=19)。洪德元所報道的2n=22m+2sm+14st(2sat)核型中發現了0~5條數目不等的B染色體,但是本報道未發現B染色體。本報道與王任翔所報道2n=38=24m+4sm+10st(2sat),冉志華所報道的2n=38=24m+2sm+12st(2sat)及黃錦嶺所報道的2n=38=20m+6sm+12st(2ast)頗為相似[12, 18-19],但本研究中并沒發現有隨體存在。
2.1.4 鳳凰蜘蛛抱蛋AspidistrafenghuangensisK. Y. Lang鳳凰蜘蛛抱蛋為武陵山區特有種,該種染色體數目是2n=38,染色體基數為x=19,二倍體(圖1,D)。核型公式為2n=18m+14sm+6st,沒有觀察到隨體,在第1對(7.86~6.95 μm)與2-6對(5.96~3.71 μm)之間在長度上有一個明顯的差別(0.99 μm),第6對與第7-19對(3.06~0.88 μm)也存在一個明顯的差別(0.65 μm)。最長染色體長度為7.86 μm,最短染色體長度為0.89 μm,最長染色體與最短染色體的比值為8.83,臂比小于2的染色體所占比例為55%。Stebbins’不對稱核型為2C。核型不對稱系數As.K%為65%。其核型表現為長柱開口箭型(三型,x=19)。與鄧濤[42]所報道的2n=38=22m+6sm+10st核型公式有所差別,但核型不對稱性一致。
2.1.5 萬年青Rohdeajaponica(Thunb.) Roth本研究中染色體數目為2n=38,染色體基數x=19,二倍體(圖1,E)。核型公式為2n=12m+24sm+2st,沒有觀察到隨體,在所有染色體中不存在一個明顯的差別,呈從大到小逐漸變化的趨勢。最長染色體長度為4.99 μm,最短染色體長度為0.88 μm,最長染色體與最短染色體的比值為5.67,臂比值小于2的染色體數所占比例為53%。Stebbins’不對稱核型為2C。核型不對稱系數As.K%為67%。其核型表現為鈴蘭型(單型,x=19)。本結果與尚宗燕等[43]和Yamashita等[29]報道的染色體數目一致,但本研究結果的不對稱性核型為更不對稱的2C型,與它們報道的2B型不同。但在本研究中萬年青屬的最長與最短染色體之比(5.67)是5個種中最小的,同時臂比值小于2的染色體所占的比率也是最低的,僅為53%,總體來說萬年青屬的對稱性比蜘蛛抱蛋屬更為對稱。萬年青屬與開口箭屬、鈴蘭屬和吉祥草屬的核型頗為相似,均為鈴蘭型核型。
2.2 染色體收集結果
本研究共收集鈴蘭族中7屬(包含Campylandra屬)共81個種,253條染色體報道記錄(表5)。這些報道主要集中在蜘蛛抱蛋屬(170余種)內,其中187條染色體報道記錄來自于蜘蛛抱蛋屬,代表了48種,其染色體數目有兩種(2n=36或2n=38), 染色體基數為x=18或19,絕大多數種為二倍體,少數為四倍體和六倍體;長柱開口箭屬(29種)的記錄有41條,代表21種,染色體數目均為2n=38,基數為x=19,均為二倍體;Campylandra屬(16種)的報道有11條記錄,代表了6個種,鈴蘭屬7條記錄代表全部2個物種,單種屬萬年青3條記錄、吉祥草屬2條記錄、白穗花屬1條記錄,這5個屬的染色體數目均為2n=38, 基數為19,均為二倍體。
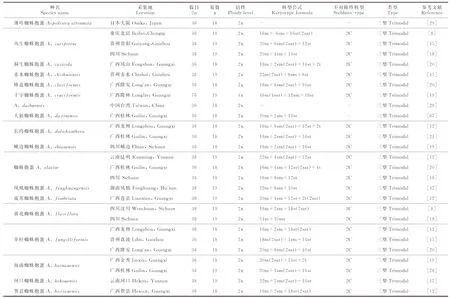
表5 鈴蘭族已報道種的染色體信息



2.3 系統發育分析與染色體進化分析
我們把染色體數目和核型類型映射到鈴蘭族系統發育樹上來探討它們的染色體進化關系(圖2)。分子發育樹的結果表明鈴蘭族分為三支,第Ⅰ支為鈴蘭-白穗花分支,是鈴蘭族首先分化出來一支,它們都以x=19為基數;第Ⅱ支為萬年表-吉祥草分支,染色體基數為x=19;第Ⅲ支為蜘蛛抱蛋-長柱開口箭分支,染色體基數為x=18和19。因此我們推測x=19鈴蘭族的原始基數,x=18是由x=19通過減少1對小染色體得到的。而在核型不對稱性方面基部鈴蘭-白穗花分支大多具有比較對稱的2B型核型,而處于進化位置上的蜘蛛抱蛋-長柱開口箭大多具有更不對稱的2C或3C型,因此我們推測本屬植物在核型不對稱性方面是從對稱向不對稱的方向進化的。另外,結合從長度變異和染色體基數來看,基于Yamashita和Tamura的核型分類法,鈴蘭-白穗花分支具有鈴蘭型核型位于基部,其進化方向是從鈴蘭屬型向長柱開口箭屬型,再到蜘蛛抱蛋屬型的演化。

單型. 指染色體間在長度上沒有明顯的差異,呈逐漸過渡的趨勢,染色體基數x=19的鈴蘭型;三型. 指由3組不同長度的染色體組成,并結合中期染色體的基數和長度變異模式將鈴蘭族分為:染色體基數為x = 19時為長柱開口箭型,染色體基數為x = 18時為蜘蛛抱蛋型圖2 基于葉綠體片段rbcL、matK、psbA-trnH 和trnL-F和核基因ITS鈴蘭族分子分子系統樹Unimodal refers to the Convallaria type which shows Unimodal trend of gradual transition without obvious difference in length between chromosomes, with chromosome number x=19; Trimodal refers to the three groups of chromosomes with different lengths, combined with the base number and length variation pattern of metaphase chromosomes, can be divided into two groups: Tupistra type when the chromosome number is x=19, and Aspidistra type when the chromosome number is x=18Fig.2 The phylogenetic tree based on 4 chloroplast gene(rbcL, matK, psbA-trnH and trnL-F) and nuclear gene ITS of Convallarieae and allied tribe
3 討 論
3.1 蜘蛛抱蛋屬染色體核型特征
蜘蛛抱蛋屬是鈴蘭族中擁有物種數目最多,形態變異最復雜多樣,生長的地理環境也最為廣泛的一個屬[13]。結合前人報道的187條記錄,加上本研究對蜘蛛抱蛋、海南蜘蛛抱蛋、四川蜘蛛抱蛋、鳳凰蜘蛛抱蛋的實驗結果,共191條記錄代表48個物種。本屬植物近年來發表了百余新種,目前為止已經包括170余種[16, 44-50],因此還有大量的物種急需開展細胞學研究。從現有的染色體數據來看,蜘蛛抱蛋屬的染色體進化表現非常復雜,既有多倍化和非整倍化的演化,同時還兼具二倍體水平上的結構變異以及隨體,B染色體的有無等特征。在蜘蛛抱蛋屬中二倍體水平上的結構變異占主導地位,多倍化和非整倍性變異僅在少數種中發現。本屬植物的染色體通常由3種類型的染色體組成:第1對為大型的中部著絲粒染色體(m),第2至第9或10對為中等大小的亞中部或亞端部著絲粒染色體(sm或st),其余為小型的中部或亞中部著絲粒染色體。李光照[2]把本屬植物的核型分為兩類:一類為2n=36=18(L+M)+18S(2sat),即9對大中型染色體和9對小型染色體;另一類為2n=38=16(L+M)+22S(2sat)。本研究中的蜘蛛抱蛋為第一種類型,由9對大中型染色體和9對小染色體組成,但四川蜘蛛抱蛋、海南蜘蛛抱蛋、鳳凰蜘蛛抱蛋與李光照[2]發現的規律有所不同,僅有6對大中型染色體,其余皆為小型染色體,而且在本研究中4個種都沒有發現隨體染色體和B染色體。這表明染色體在分裂時可能受各種因素影響或在減數分裂時染色體互換和重組過程中發生了不同染色體之間的染色質重組,從而導致染色體的長度組成比例發生變化。不同物種之間,甚至在同一物種的不同居群間或同一居群不同個體間也會發生染色體結構上的變異,而且這種變異通常與外部形態變異相關。如小花蛛蛛抱蛋、四川蛛蛛抱蛋中就有不同居群間核型多態性與表型變異存在密切相關性[2]。
從現有的統計結果來看,染色體2n=36的類群比2n =38的類群通常都多1 對中型染色體,少2對小型染色體,這可能是由于2對小的染色體融合成為1對中型染色體。染色體融合與斷裂事件在植物進化的過程中起到了一定的作用,這在許多類群中也有發現,如陳耀華等[51]發現在石蒜屬植物中有染色體的融合的情況發生。周勇等在研究3種禾本科植物時發現水稻的12條染色體經過2次染色體融合事件最終形成了高粱的10條染色體,隨后又經歷了染色體融合事件,最終形成了玉米的10條染色體[52]。當然在蛛蛛抱蛋屬中2n=36和2n=38之間的變異機制有待進一步研究。
3.2 鈴蘭族染色體核型進化模式
近年來基于分子系統發育結果來探討染色體進化機制已經在很多類群中廣泛應用[53-58]。中國學者大多認為蜘蛛抱蛋屬為二型性核型,分為大中型染色體和小型染色體兩種[2, 8, 12-17, 19, 26],Yamashita發現第1對大的中部著絲粒的染色體在蜘蛛抱蛋屬中是非常顯著的特征,因此把第1對大的染色體單獨分出來,因此從長度上來說定義為三型更為合理,并結合分子系統發育分析結果,把鈴蘭族植物的核型分為3種類型:x=18的三型染色體僅在蜘蛛抱蛋屬中發現,因此命名為蜘蛛抱蛋核型;x=19的三型染色體命名為長柱開口箭核型;x=19的一型染色體為鈴蘭型。本研究基于4個葉綠體基因和ITS核基因的聯合分析,重建鈴蘭族代表類群的系統發育關系,并結合染色體數目和核型分析發現:白穗花屬和鈴蘭屬形成一支位于整個族的基部,它們均具有x=19的基數,染色體在長度上呈逐漸變小的趨勢,也就是鈴蘭型;族中第二個分支由吉祥草屬、開口箭屬和萬年青屬組成,它們核型特征與第一分支相似,也是鈴蘭型;第三支由蜘蛛抱蛋屬、長柱開口箭屬和Tricalistra組成,其中蜘蛛抱蛋屬獨立構成一支,而Tricalistra嵌在長柱開口箭屬中。在第三個分支中蜘蛛抱蛋屬的核型既有長柱開口箭型也有蜘蛛抱蛋型,結合分子系統發育結果來看,鈴蘭族中處于基部的類群以及與之親緣關系最近的開口箭屬的染色體基數均為x=19,因此推測在蜘蛛抱蛋屬中染色體的原始基數應為x=19,而x=18是后來演生出來的基數。換而言之,鈴蘭族中染色體核型進化的趨勢是從鈴蘭型演化為長柱開口箭型,再到蜘蛛抱蛋型,本研究結果與Yamashita等[29]基于rbcL和trnK兩個葉綠體片段分析的結果一致,但本研究中選取更多的序列聯合分析,得到更高的支持率,3個分支的演化關系更為清楚,但要解決蜘蛛抱蛋屬和長柱開口箭屬下種間的關系還需要更多分子數據的進一步研究。
劉安莉等[16]認為染色體基數x=18的種類大多數種的花被闊鐘狀或鐘狀,根狀莖小于10 mm以下,花梗較長,花被片反卷,花被內基部無明顯突起,雄蕊著生位置明顯高于柱頭或與柱頭等高,柱頭較小,表面結構及紋飾簡單,果實表面光滑等特征是原始的類型;染色體基數為x=19的類群中,有諸多演生性狀,如花的形態和結構較復雜,花被壇狀,雄蕊著生位置低于柱頭,柱頭較大,表面形狀及紋飾特化成蘑菇狀等特征。通常認為植物的外部形態特征的變異是由基因和環境變異共同決定的,受光照、溫度、水分、海拔、營養物質等環境因子影響,具有很強的表型可塑性[59-62]。因此推測某一個特征是否為原始性狀,不能僅從結構簡單或復雜來判斷。現代分子技術手段為推測物種演化方向提供了最為可靠的證據,因此利用分子系統發育手段綜合考慮它與近緣類群在外部形態、染色體核型特征的區別與聯系,從而對性狀和核型的演化方向作出科學合理的判斷。
3.3 鈴蘭族染色體核型不對稱性進化模式
中國學者對于蜘蛛抱蛋屬的核型不對稱性也進行了大量的探討,他們普遍認為蜘蛛抱蛋屬植物花的形態特征與核型進化相關,花形態結構越簡單,核型不對稱程度越強,形態結構越復雜特化,核型不對稱程度就越弱,因此推測在蜘蛛抱蛋屬內核型不對稱性是由不對稱性向對稱性的方向演化[2, 16]。在鈴蘭族中Stebbins’不對稱性核型為2B、2C,少數為3C,僅在長梗開口箭中發現有1A。結合分子系統發育結果來看,位于鈴蘭族最基部的鈴蘭屬為2B型,其次吉祥草屬、萬年青屬也為2B型,其他種類基本為2C型,因此可以推測鈴蘭族的染色體核型不對稱性總體表現為由對稱向不對稱性增強的趨勢。
3.4 鈴蘭族的染色體數目與核型演化趨勢與意義
結合本研究對鈴蘭族各屬的細胞學報道以及對本族染色體資料進行了統計分析,結果表明在鈴蘭族的染色體基數與系統關系方面,鈴蘭族的染色體數目變化和分子系統發育相對一致。鈴蘭族的染色體基數是x = 18、19,根據中期染色體的基本數目和形態將鈴蘭族分為單型和三型2種核型類型。單型主要包括吉祥草屬、萬年青屬、Campylandra、鈴蘭屬和白穗花屬,染色體基數均為x=19,而三型包括蜘蛛抱蛋屬、長柱開口箭屬和Tricalistra,染色體均為x=19,部分蜘蛛抱蛋為x=18。我們認為中期染色體的三型染色體核型是由單型染色體核型衍生而來。然而,需要通過更多分類群的系統發育分析以及更多類群更精確的核型分析來進一步確認。鈴蘭族絕大多數種是2n=36、38的二倍體,其核型具有穩定性,存在少數多倍化現象,鈴蘭族染色體核型的差異正是環境因素和結構變異共同作用的結果。蜘蛛抱蛋屬植物的染色體倍性變異多數為2x和4x兩種,泰國蜘蛛抱蛋為2n=114,為染色體基數x=19的六倍體。西林蜘蛛抱蛋和十字蜘蛛抱蛋兩種為四倍體種2n=4x=76,且為同源四倍體,植物多倍化可能是對惡劣生存環境適應的結果,在蜘蛛抱蛋屬中發現多倍體的類群在外部形態方面與干旱環境相適應有一定的相關性[13]。整倍體變異和非整倍體變異不是該屬植物演化的主要趨勢,蜘蛛抱蛋絕大部分類群是二倍體,這表明該屬植物的系統發育主要是在二倍體水平上進行演化。
王任翔等[19-20]認為蜘蛛抱蛋屬植物的演化與高等植物核型的一般進化趨勢向不對稱性增強的方向演化的規律正好相反,也與該屬的近緣屬長柱開口箭屬中的核型演化趨勢相反。但是根據蜘蛛抱蛋屬植物染色體不對稱系數可以看出,染色體數目不同的兩組間染色體不對稱指數的變化相對較小,而x= 18組的染色體不對稱指數明顯低于x=19組。我們的結果表明,x=19為族內染色體原始基數,x=18為派生基數,而x=18是從x=19演化而來。通過對染色體長度研究表明本族染色體低基數的x=18由x=19通過染色體融合或丟失進化而來,而核型則由比較對稱的單型向不對稱的三型演化。所以祖先類群的核型對稱性高于派生類群,蜘蛛抱蛋屬的核型可能是向對稱性增強的方向演化。因此,從目前已有的核型資料看,鈴蘭族植物符合Stebbins所提出的核型不對稱性增加的趨勢。長柱開口箭屬中絕大部分種的染色體屬于一型,染色體長短是遞增排列的較為對稱,傘柱長柱開口箭染色體中短染色體較明顯,但是中長染色體與長染色體的長度很接近,染色體長度不對稱因此屬于二型染色體,傘柱開口箭這個種是能夠代表長柱開口箭屬向蜘蛛抱蛋屬演化的一個證據,而從染色體的類型來看開口箭屬比蜘蛛抱蛋屬更為原始[10]。由研究結果看來蜘蛛抱蛋屬和長柱開口箭屬之間的核型演化關系和系統發育關系很明顯較親密,兩屬確定為姐妹類群。本研究揭示鈴蘭族由高基數向低基數減少以及單型向三型演化的趨勢,按照染色體長度鈴蘭族核型演化趨勢由對稱向不對稱演化,核型在屬間和屬內都有很大變化。鈴蘭族的核型的進化趨勢表明鈴蘭屬為擬原始特征,由此衍生出長柱開口箭屬,進而衍生出蜘蛛抱蛋屬。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
