克里雅河尾閭綠洲胡楊種群年齡結構對地下水埋深的響應
2021-09-16 14:37:26史浩伯師慶東周小龍萬彥博
西北植物學報 2021年8期
史浩伯,師慶東,3*,戴 岳,3,周小龍,3,萬彥博,彭 磊
(1 新疆大學 資源與環境科學學院,烏魯木齊 830046;2 綠洲生態教育部重點實驗室,烏魯木齊 830046;3 新疆大學干旱生態環境研究所,烏魯木齊 830046)
植物種群的年齡特征是數量特征中的重要指標之一[1],對研究種群的未來發展動態與群落的更新演替均能起到良好的分析預測作用[2-4],在探討某一生境內的植物在生長發育過程中所呈現出的特征時,往往可以根據現階段的年齡結構給出其在現狀方面的良好解釋[5],有助于深化在種群尺度上對植株生長特性的認知,了解植被格局的形成機制,這對于受不良環境條件脅迫以及處在瀕危階段的植物保護工作有著重要的指導意義[6]。同時,在植物格局的形成過程中并不僅受生存環境的影響,生物學特性也是植物群落在構建過程中不可忽視的因素[7],同樣可能影響種群的結構動態和繁殖過程,而競爭關系作為自然界同種或異種個體之間爭奪共同資源的常見現象,在植物功能性狀的形成、多樣性格局的演變以及共存機制方面均具有一定的驅動意義[8-9]。
胡楊(Populuseuphratica)是大陸性干旱氣候條件下的常見樹種,廣泛分布于中國西北以及中亞等干旱地區,耐寒耐澇,對惡劣環境的抗性極強,是荒漠區特有的森林資源與關鍵種,在遏制荒漠化進程與風沙防治方面具有重要作用[10]。目前針對胡楊的研究以塔里木河流域與黑河流域居多,在宏觀尺度上的研究主要集中于胡楊的生活史特征、空間分布格局與動態等方面[11-13],同時在生理特征與遺傳分化等微觀尺度上也取得了豐碩的成果[14-15],多元化的研究方向與研究成果的疊加足以說明胡楊對于荒漠生態系統的重要性,因此在對干旱生境演變與荒漠植物種群演替的研究過程中,胡楊往往具有較強的指示作用。
克里雅河是中國新疆南部的第四大水系,其尾閭達里雅布依深入塔里木盆地腹地300 km,是極具科學研究價值的“原始綠洲”[16],當地種群結構簡單,自我更新與繁殖能力均受到了干旱條件的制約[17],而胡楊作為當地的主體樹種,在維持綠洲生態系統穩定,減緩沙漠腹地荒漠化對綠洲侵占進程方面具有決定性意義。本研究為闡明地下水埋深制約條件下沙漠腹地胡楊種群的生長狀況及演替動態,通過小尺度試驗擬解決兩個科學問題:(1)當前環境條件下的胡楊種群呈現出怎樣的更新特征?(2)不同地下水埋深條件下的胡楊種群年齡結構有何差異?通過上述分析旨在說明地下水因子對胡楊年齡結構與更新特征的影響,為宏觀尺度上的綠洲胡楊林保護及生態修復等方面提供參考依據。
1 研究區概況
達里雅布依位于世界第二大沙漠塔克拉瑪干沙漠腹地,是中國塔里木盆地南緣第二大河流克里雅河的尾閭綠洲,該綠洲因其與世隔絕、交通不便而受到干擾因素較少,基本保持了自然條件下的狀態,具有相對“原始”特征。達里雅布依地理位置為38°16′~38°37′ N,81°05′~81°46′ E,海拔為1 100~1 300 m,總面積約342 km2。該區域為暖溫帶干旱沙漠氣候,年降水量不超過20 mm,潛在蒸發量達2 480 mm,年均溫12.1 ℃,晝夜溫差大,氣候極端干旱。綠洲內部具有網狀河相沉積特征,干枯河道分布于綠洲內部,土壤以沙土為主,天然植物群落則主要由胡楊、檉柳(Tamarixchinensis)、駱駝刺(Alhagisparsifolia)、蘆葦(Phragmitescommunis)等物種組成[18]。
2 研究方法
2.1 野外試驗
課題組于2018年結合生境異質性在綠洲內布設了19口地下水觀測井,并于2019、2020年7月前往達里雅布依綠洲進行野外實地踏查,通過典型樣地法選取胡楊種群分布集中且林分保持良好的地段,以所選擇的觀測井為中心布設11個20 m×20 m的植被樣方,樣地設置如圖1所示。在運用手持GPS對樣方進行定位后開展胡楊種群逐木測量工作,利用卷尺、游標卡尺等工具對胡楊的胸徑、株高、冠幅等基本數據進行測量,并統計種群的個體數量,同時基于地下水觀測井內的HOBO水位自動記錄儀獲取地下水埋深數據。
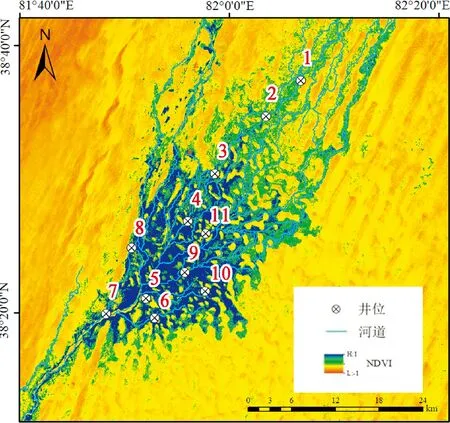
1~11.代表樣地位置圖1 研究區樣地位置示意圖1~11 represent the location of the selected sample plotsFig.1 Sketch map of study area
2.2 數據處理與分析
2.2.1 地下水埋深梯度劃分采用K-均值聚類方法劃分地下水埋深梯度,將其梯度分為三個等級(圖2):0~3 m的地下水埋深為第一梯度等級(A);3~6 m的地下水埋深為第二梯度等級(B);6~9 m的地下水埋深為第三梯度等級(C)。
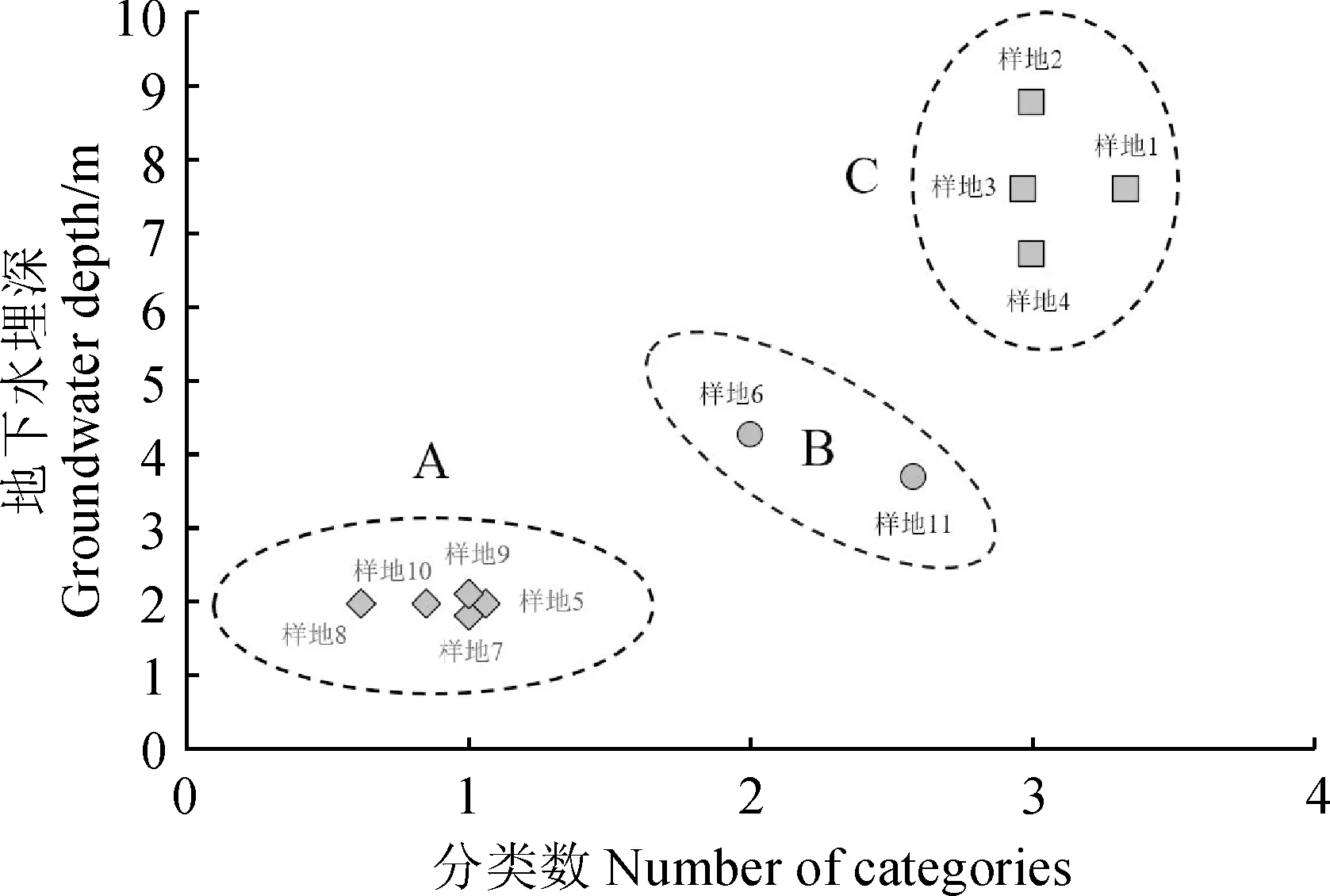
圖2 研究區綠洲地下水埋深梯度聚類結果Fig.2 Gradient clustering results of groundwater depth
2.2.2 齡級劃分結合胡楊的生活史特點可將其劃分為12個齡級[19],分別是Ⅰ齡級: 0 cm<胸徑≤4 cm;Ⅱ齡級: 4 cm<胸徑≤8 cm;Ⅲ齡級: 8 cm<胸徑≤12 cm;Ⅳ齡級: 12 cm<胸徑≤16 cm;Ⅴ齡級: 16 cm<胸徑≤20 cm;Ⅵ齡級: 20 cm<胸徑≤24 cm;Ⅶ齡級: 24 cm<胸徑≤28 cm;Ⅷ齡級: 28 cm<胸徑≤32 cm;Ⅸ齡級: 32 cm<胸徑≤36 cm;Ⅹ齡級: 36 cm<胸徑≤40 cm;Ⅺ齡級: 40 cm<胸徑≤44 cm;Ⅻ齡級: 胸徑>44 cm。同時結合胡楊生活史特點,可將胡楊的年齡等級進行匯總,進一步分為幼齡林(Ⅰ~Ⅲ)、中齡林(Ⅳ~Ⅶ)、成熟林(Ⅷ~Ⅺ)和老齡林(Ⅻ)4個等級,以便降低年齡結構劃分的復雜程度。
2.2.3 年齡結構動態變化分析胡楊種群的年齡結構是一種靜態的生態學特征,應通過量化手段將其呈現出時空尺度上的直觀動態變化,文中選用相鄰齡級個體數量動態變化指數(Vn)與種群整體年齡結構動態系數(Vpi)進行種群發育動態分析[20],方法如下:
式中,Sn與Sn+1分別表示第n與第n+1齡級種群的個體數量,Vn存在值域[-1,1],當Vn取值為正、負、零時分別代表該種群在兩個相鄰齡級的個體數量呈增長、衰退、穩定的結構動態關系。Vpi是由不同年齡級間的動態變化指數對個體數Sn加權分配得來的,乘以權重后因為并不存在最大年齡級,所以將k這一最大齡級排除,最終Vpi的結果也存在正、負與零3種不同類型的值,且意義與Vn一致。
2.2.4 年齡結構動態預測運用一次移動平均法能對以年齡級為生長周期單位的胡楊種群數量動態進行有效預測[21],方法如下:
式中,n代表期望預測的時間長度,Mt代表n個生長周期后t齡級種群的數量大小,Xk是當前k齡級種群的大小,本研究分別對未來2個和5個齡級時間后的胡楊種群年齡結構進行預測。
2.2.5 生長空間競爭指數模型已有研究表明,干旱地區的胡楊冠幅與胸徑的生長狀況與地下水埋深有顯著關系[22-24],因此如何建立以水資源作為限制條件的競爭關系模型極為重要。本研究基于馬履一提出的生長空間競爭指數模型[25]進行推導,以此適應荒漠生態系統胡楊的生長特性。已知生長空間競爭指數(growth space competition index,GSCI)公式如下:
GSCI=CASi/BAi
由于胡楊生長受環境條件的限制導致樹冠形狀呈多樣化,為更準確地說明其徑向生長情況,故選用投影面積(SA)代替原始公式中的樹冠表面積(CAS),具體公式為:
SA=CAS= π· [(a+b) / 4]2
式中,CASi代表競爭木的樹冠表面積,BAi代表競爭木的胸高斷面積,a為東西方向的冠幅,b為南北方向的冠幅。GSCI值越小,反映單位面積需要的樹冠面積越小,其競爭能力越強,同時作為樹冠覆蓋程度的無量綱度量,是一種相比于生態過程更注重競爭結果的度量方法。
3 結果與分析
3.1 綠洲胡楊種群年齡結構與動態分析
達里雅布依綠洲內的胡楊整體年齡結構呈“不規則形”(圖3)。根據本研究擬定的齡級結構劃分標準,中齡林、成熟林在現有植株數量中占比最少,分別為0.19與0.14;其次是幼齡林數量,占比0.25;Ⅻ齡級以上的老齡林數量最多,占比0.40。總體來看,在綠洲胡楊種群的年齡結構中老齡化現象明顯,在中齡林與成熟林兩個年齡組分中存在斷層現象,但幼齡林組分尚具規模,其補員作用能夠在一定程度上緩解種群在更新方面的壓力。
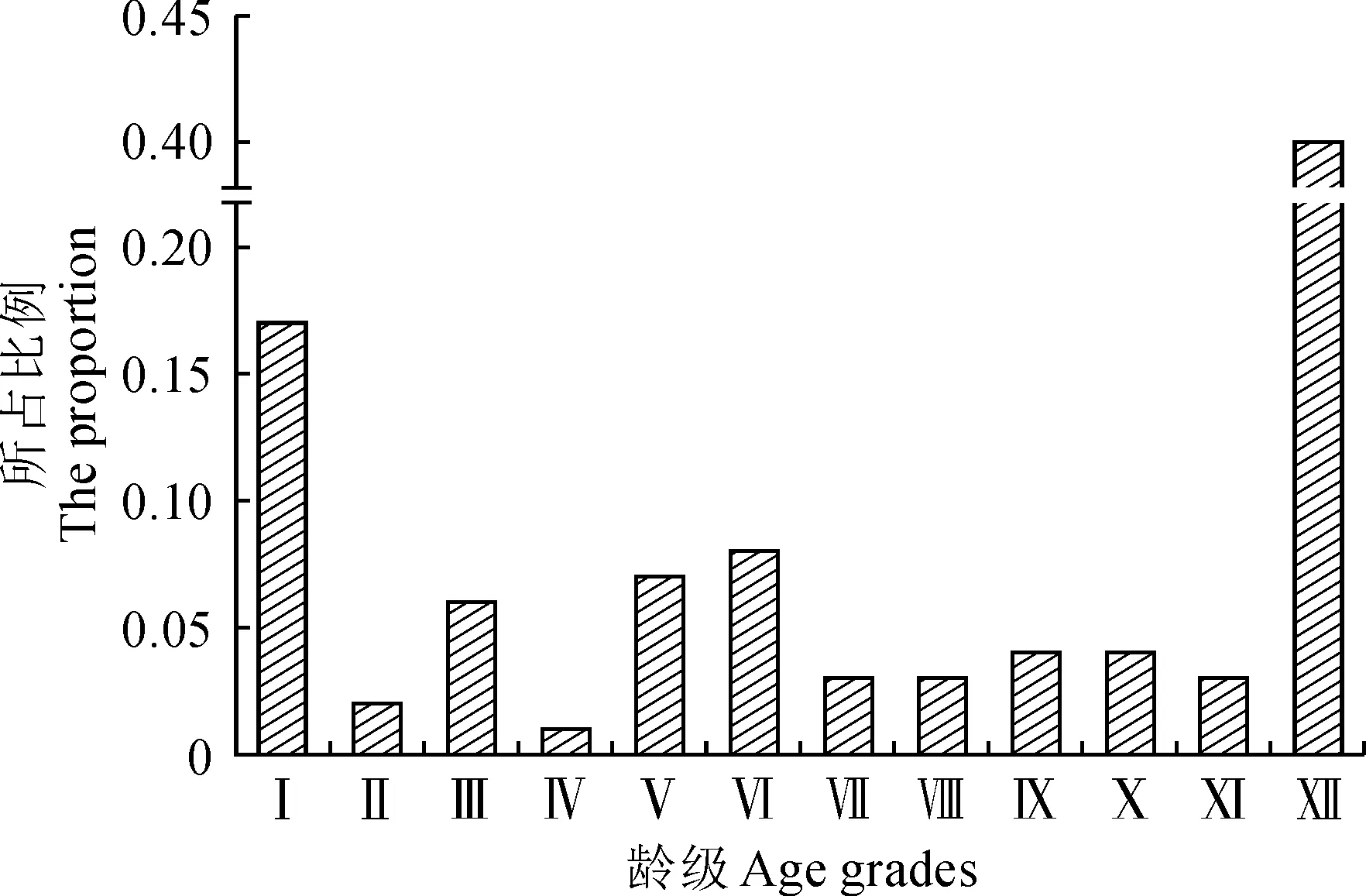
Ⅰ~Ⅻ.齡級;DBH. 胸徑; Ⅰ. 0 cm
將種群動態量化后所得出的相鄰年齡級之間的動態變化指數(表1)表現出較強的隨機性:當前綠洲內的Ⅰ齡級的胡楊幼苗數量具備更新基礎,與Ⅱ齡級之間呈增長的結構動態關系,與之相似,Ⅲ和Ⅳ、Ⅵ和Ⅶ、Ⅹ和Ⅺ三對相鄰齡級之間同樣存在增長關系;而在Ⅱ和Ⅲ、Ⅳ和Ⅴ、Ⅴ和Ⅵ、Ⅷ和Ⅸ、Ⅺ和Ⅻ五對齡級之間的相鄰年齡結構動態呈現出衰退趨勢;Ⅶ和Ⅷ、Ⅸ和Ⅹ兩對相鄰齡級的種群動態則呈穩定關系。綠洲胡楊整體種群的年齡結構動態指數Vpi=29.55%,其數值趨近于零,說明種群整體現階段較為穩定,雖具有一定的增長趨勢但并不明顯。
3.2 年齡結構與生長空間競爭指數變化關系
胡楊個體的生長空間競爭指數與齡級的線性擬合關系呈現一定的規律性(圖4),擬合方程為y=66.79±4.28x-2.08±0.6,R2=0.69。這一結果說明胡楊種群的年齡結構與個體競爭能力之間存在冪函數關系,表明依據胸徑特征劃分出的齡級能夠在一定程度上說明物種的個體競爭能力,即隨著胡楊的齡級的上升,個體的競爭能力會逐步加強,但加強的幅度持續變低。
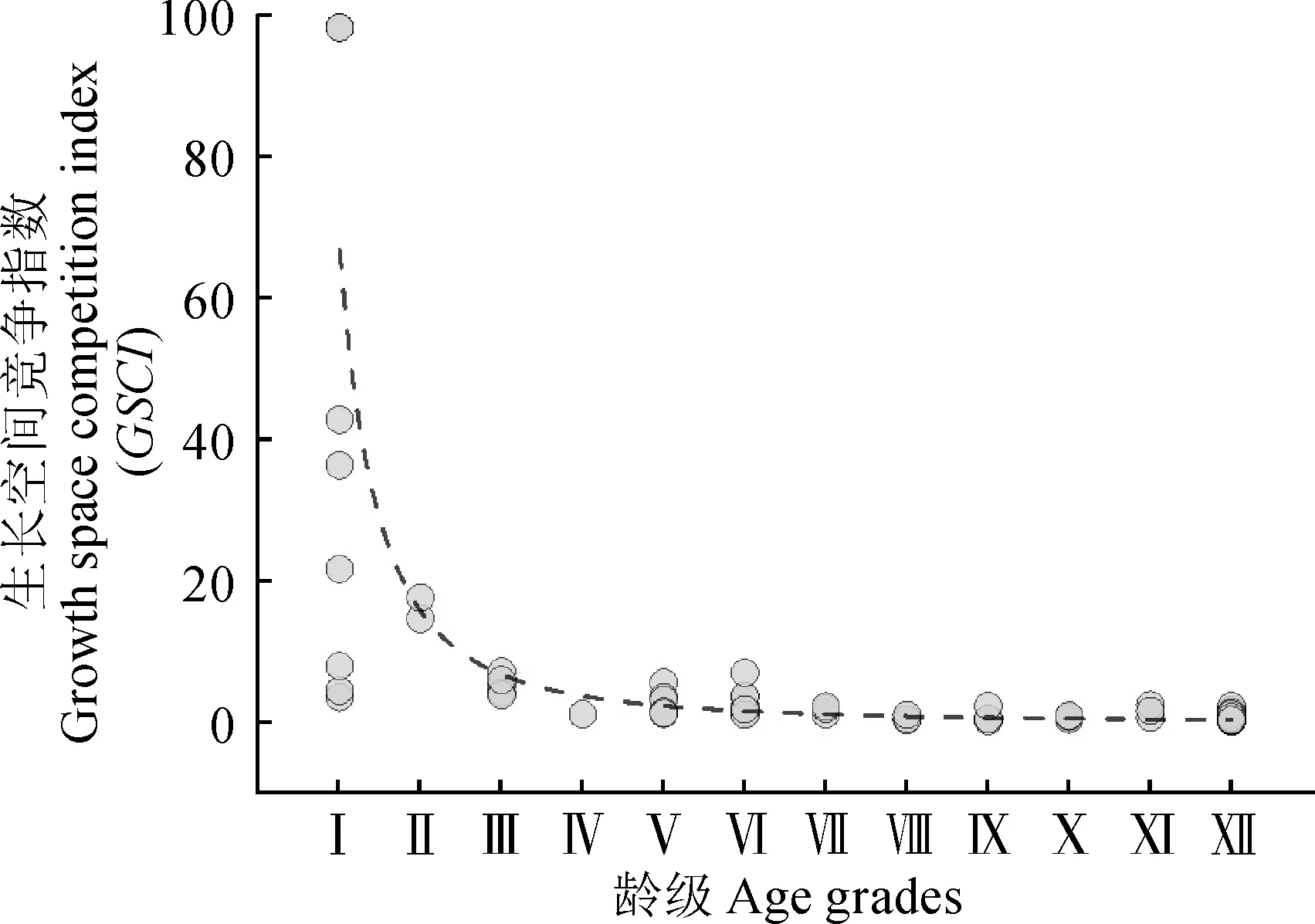
圖4 研究區綠洲胡楊齡級與生長空間競爭指數變化關系Fig.4 Relationship between P. euphratica age grade and growth space competition index
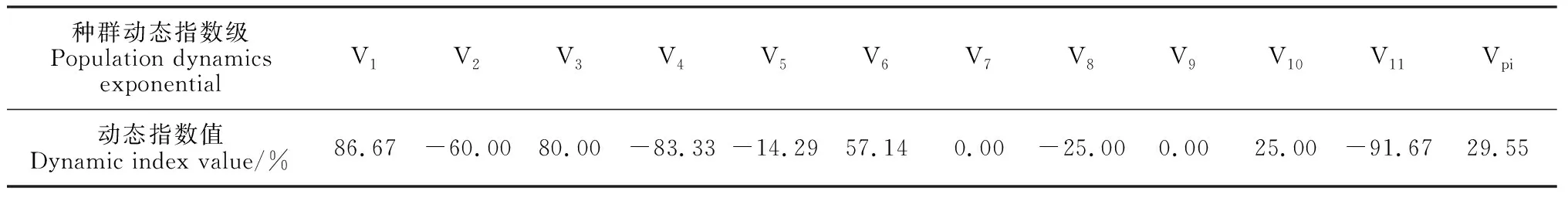
表1 胡楊種群齡級結構動態指數
3.3 地下水埋深梯度下的種群年齡結構變化
胡楊種群年齡結構與個體競爭能力存在較好的線性關系,因此可以通過不同地下水埋深梯度下的具體生長空間競爭指數來判斷生境對種群生長的維持情況。如圖5所示,在地下水埋深處于0~3 m的第一梯度(A)時,胡楊種群的生長空間競爭指數的變異系數平均值為1.09,該梯度下胡楊個體競爭能力的離散程度與其他兩個梯度相比處于中等水平,各樣地種群具體的年齡結構可歸為兩種類型,一類是僅存在胸徑大于44 cm的老齡林群落,另外一類則是年齡結構復雜,不同齡級個體均有出現的多齡級群落;在地下水埋深處于3~6 m之間的第二梯度(B)時,競爭指數的變異系數平均值為2.176,離散程度較高,雖各自樣地均僅包括2個齡級,但總體的誤差值高達2.167,說明了這一地下水埋深梯度下胡楊個體的競爭能力存在較高的可波動范圍;在地下水埋深處于6~9 m的第三梯度(C)時,競爭指數的平均變異系數為0.634,且離散程度極低,原因在于該梯度下各樣地的胡楊種群均無幼齡林出現,成熟林與老齡林居多,占個體總數的84.85%,且現存個體齡級之間次序緊密,在年齡結構方面呈現出典型的老齡化特征。
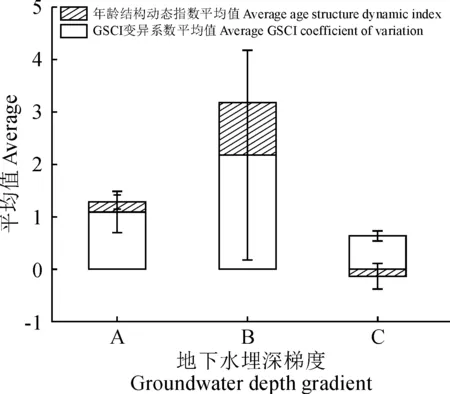
圖5 研究區綠洲不同地下水埋深梯度下胡楊年齡結構動態與個體競爭指數變異特征Fig.5 Age structure dynamics and individual competition index variation characteristics under different groundwater depth gradients
而在胡楊種群生長動態變化特征方面,不同地下水埋深梯度下的種群年齡結構動態也呈現出一定的差異(圖5)。當所處環境的地下水埋深位于第一梯度(A)時,動態指數的平均值為0.194,標準誤差為0.135,不同區域的種群生長動態存在穩定與增長兩種不同的水平;而在地下水埋深處于第二梯度(B)時,種群的年齡動態呈現極為明顯的增長特征,且不存在誤差值;在地下水埋深處于第三梯度(C)時,年齡結構動態指數平均值為-0.134,該梯度下的絕大部分區域均呈現出衰退的種群年齡結構特征,而僅存的具有增長趨勢的區域其增長動態也并不明顯,但正是這種正負關系的存在導致了該梯度范圍內出現了誤差值較大的現象。
3.4 時間序列上的種群動態預測
綠洲胡楊種群的年齡結構動態受所處地下水梯度的差別影響而有所不同(圖6)。在地下水埋深0~3 m的第一梯度(圖6,A)中,隨著時間的推移老齡林與幼齡林均有所降低,中齡林逐漸成為林地中的重要組分,該區域的種群年齡結構逐漸變得穩定,但直至5個齡級時間后依然存在增長的趨勢;在地下水埋深3~6 m的第二梯度(圖6,B)中,2個齡級時間后其結構依然為增長型,5個齡級時間后的年齡結構逐漸趨于穩定,且以幼齡林與中齡林為主,但該環境在初始條件下的個體數目較少,對時間序列上預測的波動幅度產生了一定的影響;在地下水埋深6~9 m的第三梯度(圖6,C)中,干旱的環境條件下幼齡林的不足導致胡楊年齡結構呈持續性的衰退趨勢,加強對幼苗的撫育工作是維持種群持續發展的主要手段。
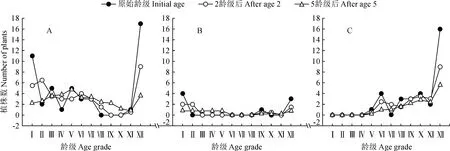
A.0~3 m;B.3~6 m;C.6~9m圖6 研究區綠洲不同地下水埋深梯度下胡楊種群動態預測結果Fig.6 Prediction results of population dynamics under different groundwater depth gradients
4 討 論
4.1 水文動態對胡楊種群更新演替的影響
克里雅河在維吾爾語中有“漂移不定”的意思,流域的形成依賴于昆侖山遠古冰川與融雪補給,受季節性洪水的影響改道頻繁[26],其尾閭達里雅布依綠洲的網狀河相也因此不能保證各條河道均有穩定的地表水經過[27]。本研究中第一資源梯度中的樣地均處于綠洲東南部,即克里雅河主河道與綠洲網狀河道的交匯處,在理論上該區域的多數河道均應依靠季節性洪水存在穩定的地下水埋深補給。第二資源梯度下的樣地則主要分布在綠洲南部與中部,最南端的河道屬于獨立的水系支流,干涸程度較重,而中部地區的河道已與主干河道存在一定距離,季節性洪水對這兩處區域的地下水的補給強度遠不及第一梯度區域,但其之所以呈現出典型的增長型年齡結構群落,很可能是6~9月的克里雅河豐水期產生的河水漫溢作用影響了胡楊實生苗的更新,提高了幼齡林的個體數量[28-29]。而第三資源梯度的區域主要分布于綠洲北部,即使是洪水期對地下水的補給作用也相對乏力,幼齡林在低水位的情況下無法生長,受此影響導致胡楊林表現為衰退型群落。綜上所述,淺層地下水作為荒漠植被生長的重要資源,是促使其更新演替的動力源泉,對胡楊的齡級結構組成有重要影響。
4.2 年齡結構與種內關系的互作機制
本項研究中選取的生長空間競爭指數是基于干旱區胡楊生長屬性特征進行量化,在忽略外界干擾情況下最大程度反映不同個體維持自身基本屬性所需的地下水埋深閾值。由圖4和圖5可知,伴隨著齡級的增長,植株個體的競爭能力也隨之上升,進而使同一區域內的種內競爭現象愈發激烈。而地下水作為荒漠區植被生存所依賴的關鍵因子,對維持物種的種內競爭關系具有重要作用。在地下水埋深處于0~3 m的情況下能夠同時維持單一老齡林的強烈競爭作用與多齡級條件下復雜的競爭機制;在地下水埋深處于3~6 m的情況下,能夠維持齡級較少的增長型群落,但這種對競爭關系的維持作用存在一定的波動閾值,不能同時滿足多齡級的存在;而在地下水埋深處于6~9 m的情況下,種群年齡結構已處于衰退狀態,激烈的競爭關系則會進一步排斥低競爭優勢的幼齡林,相關研究表明,隨著齡級增長至老齡林,林木已擺脫被壓狀態,因此低齡級林木成為了競爭關系中的主要被排斥對象[30],在無低齡級與充足地下水補給的情況下,這種衰退情況會持續加劇,直至區域性的種群完全喪失。
4.3 時間序列上的胡楊種群動態
如果以地下水埋深三種不同的梯度條件作為限制因子研究種群動態,能夠發現不同區域的種群在面臨環境條件差異時,表現出的動態特征也有所不同。地下水埋深第一梯度(A)下的胡楊種群在演替過程中表現出的增長特征極為明顯,在5個齡級時間后其年齡結構依舊完整,表現出較強的活力;而地下水埋深第二梯度(B)很可能受到了試驗過程中樣地數量的影響,導致初始條件下的胡楊個體數量較少,但在時間序列預測上依然表現出了明確的動態特征,即在5個齡級時間后趨于穩定。受地下水埋深的影響下,該種群的演替特征應處于高水位埋深與低水位埋深的過渡階段,即雖然無法滿足年齡結構的完整,但依舊具有一定的更新能力;地下水埋深的第三梯度(C)缺失了幼齡林這一整體組分,同時老齡林所占比例極高,導致在沒有補員作用下的胡楊林呈現出明顯的衰退趨勢。上述分析中所用的時間預測分析手段具有較高的準確性,適用于多種限制條件下的預測[31],同時其弊端也很明顯,預測的結果僅基于現狀進行研究而往往忽視了生物的擴散與再定殖過程[32],因此在現實的發展動態中很可能受到生物因素的影響,但在胡楊林的保護與可持續發展方面,則可以根據預測的動態結果制定針對性的幼苗撫育、生態修復等工作,這對于促進胡楊林的天然更新,維持種群的穩定性均具有重要意義。
5 結 論
(1)達里雅布依綠洲胡楊老齡林數量居多,其次是幼齡林。整體年齡結構動態存在微弱的增長趨勢,但不同生境之間的差異較大,多數區域的齡級更新依舊乏力。
(2)胡楊個體的競爭能力與徑級存在冪函數關系,受控于對地下水埋深差異導致維持的種內競爭機制存在一定的差異,即隨著地下水埋深的增加,所能維持的競爭關系愈發簡單,而個體競爭能力較低的幼齡林比例也隨之降低。
(3)隨著時間的推移,0~6 m的地下水埋深依舊能夠保障胡楊種群正常的更新演替,而大于6 m后的地下水埋深則會使胡楊種群喪失幼齡林這一組分,呈現出明顯的衰退趨勢。
