中條山油松人工林下物種對群落物種豐富度分布格局的貢獻
2021-09-16 14:37:26裴順祥杜滿義辛學兵
西北植物學報 2021年8期
裴順祥,法 蕾,杜滿義,郭 嘉,辛學兵
(中國林業科學研究院華北林業實驗中心,北京九龍山暖溫帶森林國家長期科研基地,北京 102300)
物種豐富度的分布格局是生物多樣性保護的關鍵問題之一[1]。物種豐富度格局的形成不僅依賴于群落的構建過程,同樣也依賴于群落中的物種組成[2-3]。群落中根據物種多度大小可以將物種劃分為常見種、稀有種和介于常見種與稀有種之間的物種,在大多數群落中,稀有種比常見種的數量更多,且由于稀有和常見物種的頻率分布不均勻而引發群落物種分布格局[4],因此研究物種對群落物種豐富度分布格局的貢獻是探索物種分布格局的重要手段之一[5]。
關于物種對群落物種豐富度分布格局的貢獻已開展了大量的研究,并取得了一些關鍵性結論,但仍存在不足,主要表現在以下2個方面。一是稀有種與常見種對群落物種豐富度分布格局貢獻度的大小存在爭議。群落的中性過程和個體水平上的隨機作用導致群落中存在大量的稀有種[6],進而使群落產生“稀有種優勢”[7-8],因此稀有種對整個群落的豐富度格局產生較大的影響[9-10],但也有學者認為常見種的貢獻要遠大于稀有種[11-13],即常見種“維持”了群落物種分布格局[14],而稀有種對整個群落功能的重要性可能被夸大[15],因為在實際群落中,常見種的生殖成功率要明顯高于稀有種[16],且中性理論認為群落中所有物種競爭對稱,即不存在“稀有種優勢”。研究尺度也影響稀有種和常見種的貢獻,在大尺度范圍內,稀有種的貢獻較大,而常見種在小尺度上影響物種豐富度,在大尺度上影響物種均勻度[17]。稀有種由于個體少,分布范圍受限,與常見種相比更易受到環境變化和競爭排斥的干擾[18-19],其通常被認為是特有的、局地的和受威脅的,這使得稀有種比常見種面臨更高的滅絕風險[20],但并不能根據稀有種的稀有性(頻度、豐富度和生物量低、高滅絕風險)的特征,判定其對生態系統功能、過程中的貢獻低于常見種[21]。此外,Mazaris等[22]認為常見種對物種分布格局的貢獻大是取樣偏差造成的,并非常見種與稀有種二者的生物學差異所致。二是以某一類物種組成的子群落對整體群落物種分布格局的貢獻研究較多,單一物種的貢獻研究較少,而單一物種的貢獻可能對于群落物種多樣性的保護更為重要。因此深入解析不同生態系統物種對群落物種豐富度分布格局的貢獻,對于區域物種生物多樣性保護和提升均具有重要的意義。
中條山地處秦嶺與太行山之間,其植物區系是熱帶亞熱帶區系向溫帶區系的過渡和交匯地[23],而油松(Pinustabuliformis)人工林是中條山森林生態系統的重要組成部分,趨于針葉化的、成片高密度人工林導致土壤酸化,林下灌、草難以發育形成喬-灌-草垂直空間結構的森林群落,進而導致人工林下物種多樣性低下、群落穩定性和生產力差、病蟲害頻發、抵御氣候災害能力弱[24-25],而物種多樣性的恢復和保護是生態系統恢復和重建的基礎。因此,開展油松人工林下物種多樣性格局研究具有非常重要的現實和理論意義。本研究以中條山油松人工林群落為研究對象,研究林下不同大小的子群落對群落物種豐富度分布格局的貢獻,并確定影響該區域群落物種豐富度分布格局的關鍵種,為區域物種多樣性保護提供理論依據。
1 材料和方法
1.1 數據來源及質量
在對中條山油松人工林全面踏查及盡可能涵蓋該地區所有油松人工林類型的基礎上,設置12個樣地,每個樣地設置3個20 m × 20 m的用于喬木調查的樣方(其中有3個樣方受地形限制為15 m × 15 m),共計36個喬木樣方,在每個喬木樣方內設置16個5 m × 5 m和1 m × 1 m的小樣方調查灌木、草本植物。分別調查了555個灌木和草本樣方,記錄每個樣方內物種種類、數量、蓋度、高度等指標,同時記錄樣地的經緯度、海拔、坡向、坡位、林冠郁閉度等環境特征(表1)。為了確定抽樣調查是否可以充分反映群落物種多樣性,本研究采用物種累積曲線來判斷抽樣量是否充分(圖1),由圖1可知,曲線趨于平緩,表示中條山油松人工林下的物種并不會隨樣本量的增加而顯著增多,表明抽樣充分[26]。
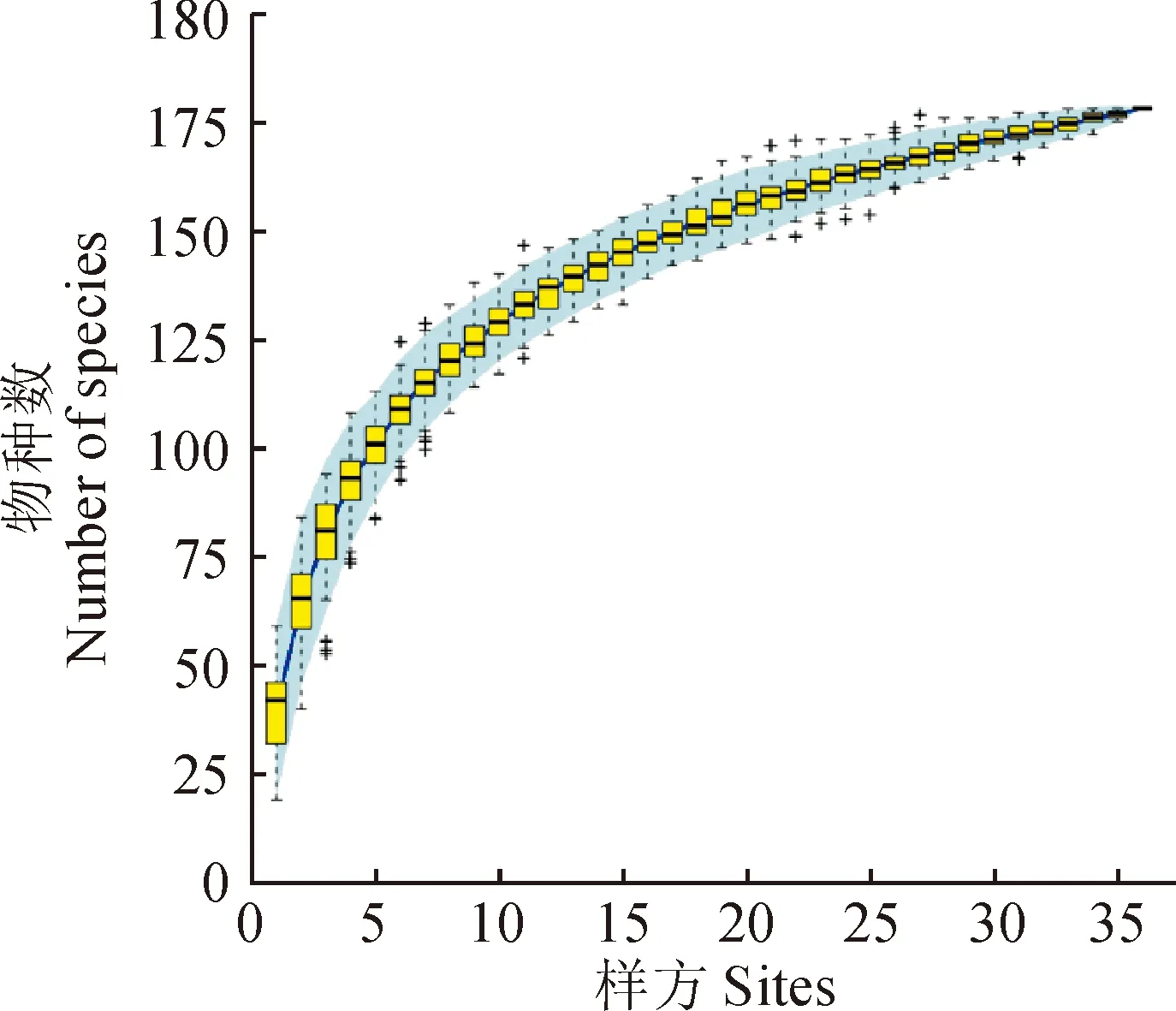
圖1 灌木、草本、藤本物種累積曲線Fig.1 Species accumulation curves of shrubs, herbs and lianas
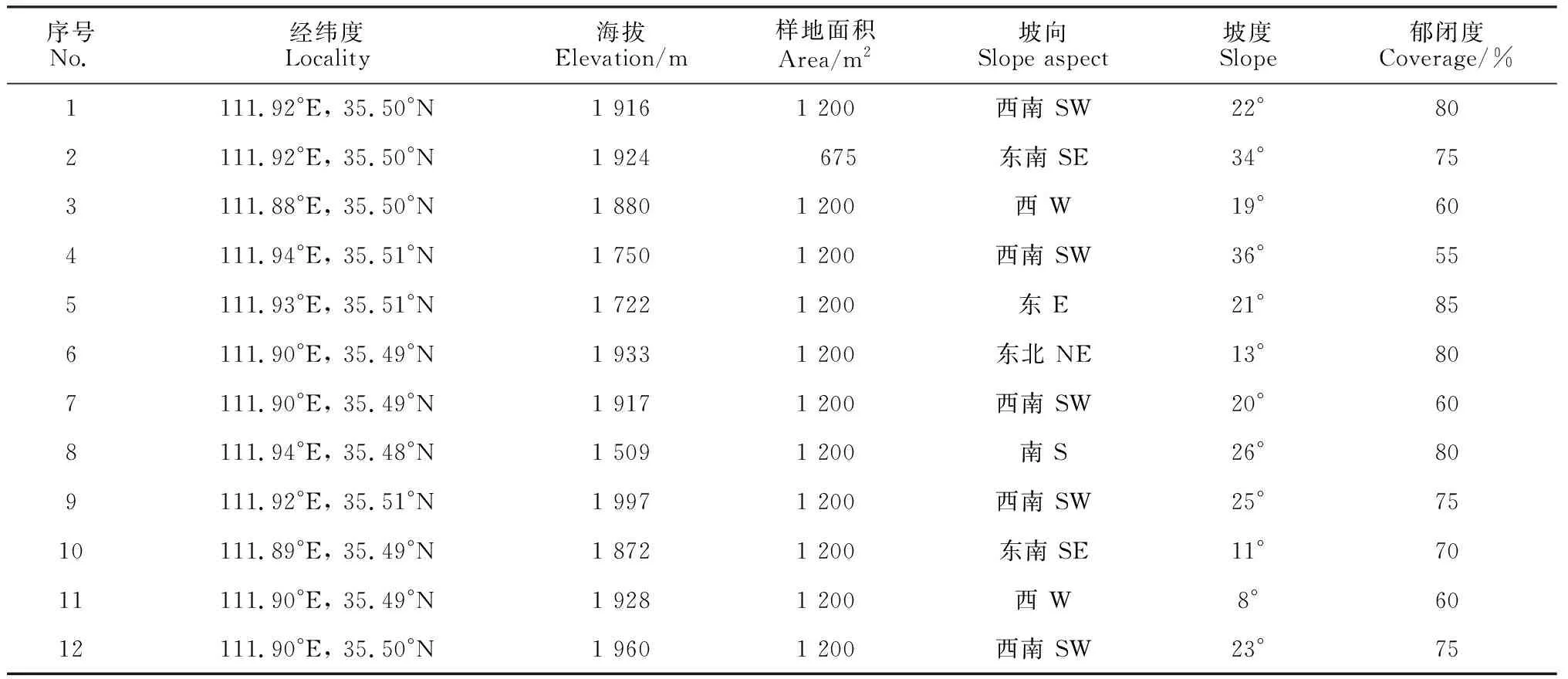
表1 油松人工林群落樣地概況
1.2 數據分析方法
本研究選取物種優勢度指數(Y)計算群落優勢種,其計算公式如下:
式中,N表示群落物種個體總數,ni表示第i種的個體總數,fi表示第i種的頻度。當Y>0.02時,該物種為群落優勢種;選取物種豐富度(R)表示多樣性;采用稀釋曲線(rarefaction curve)法分析各樣方物種多樣性的差異[27];將頻度小于等于2的物種為稀有種,頻度大于9的物種為常見種[17]。
根據物種在群落中的頻度定義物種稀有度和常見度,按頻度由低到高或由高到低的序列對物種依次排序,然后根據序列逐一添加(或去除)物種,通過比較添加(或去除)前后引起的群落物種豐富度變化的大小來衡量稀有/常見種對群落物種豐富度格局的貢獻[2-3]。
為了進一步分析最稀有/常見物種組成的子群落對整個群落物種豐富度格局的貢獻,將占群落總體物種25%的最稀有/常見的物種組成子群落,分析子群落、整個群落、除25%最稀有/常見物種子群落間的相關關系[3]。
將物種依據頻度由低到高排序,以稀有—常見為橫坐標,常見—稀有為縱坐標,橫坐標為“開始”種(starting species),縱坐標為“結束”種(finishing species)[3],178個物種共構建15 931個子群落,然后每個子群落與整個群落做相關分析,然后計算添加(或去除)物種后,子群落與整個群落相關系數變化量,依據變化量大小排序,得到影響群落豐富度格局最大的5個物種。然后將所有物種按頻度由高到低排序,以貢獻最大的種為例,基于Jaccard指數,分析貢獻度最大的種與比其頻度更高的物種間的關聯度[28]。
物種數據整理采用Excel 2016(Microsoft,USA)完成,除Jaccard指數及種間關聯分析及作圖分別采用R 4.0.3軟件中“spaa”程序包中的“sp.pair”和“plotlowertri”命令完成外,其他分析及作圖分別采用R 4.0.3軟件中“vegan”程序包中的“specaccum”、“ggplot”、“rarecurve”、“cor”命令完成。
2 結果與分析
2.1 中條山油松人工林下物種多樣性
樣地物種調查數據顯示,中條山油松人工林下共有178種植物,其中灌木47種、草本112種,藤本19種,分屬56科124屬,優勢種為披針薹草(Carexlancifolia)、茜草(Rubiacordifolia)、赤瓟(Thladianthadubia)、金線草(Rubiamembranacea)、南蛇藤(Celastrusorbiculatus)。頻度-物種數分布圖顯示(圖2),該地區物種頻度分布格局呈明顯右偏。具體表現為,頻度小于等于2的稀有種54種,頻度大于9的常見種59種,分別占全部物種的30.34%和33.15%。頻度排名前10的物種分別為披針薹草、粗齒鐵線蓮(Clematisargentilucida)、金線草、披堿草(Elymusdahuricus)、接骨木(Sambucuswilliamsii)、鞘柄菝葜(Smilaxstans)、茜草、土莊繡線菊(Spiraeapubescens)、南蛇藤、紅椋子(Swidahemsleyi),草本多于灌木。
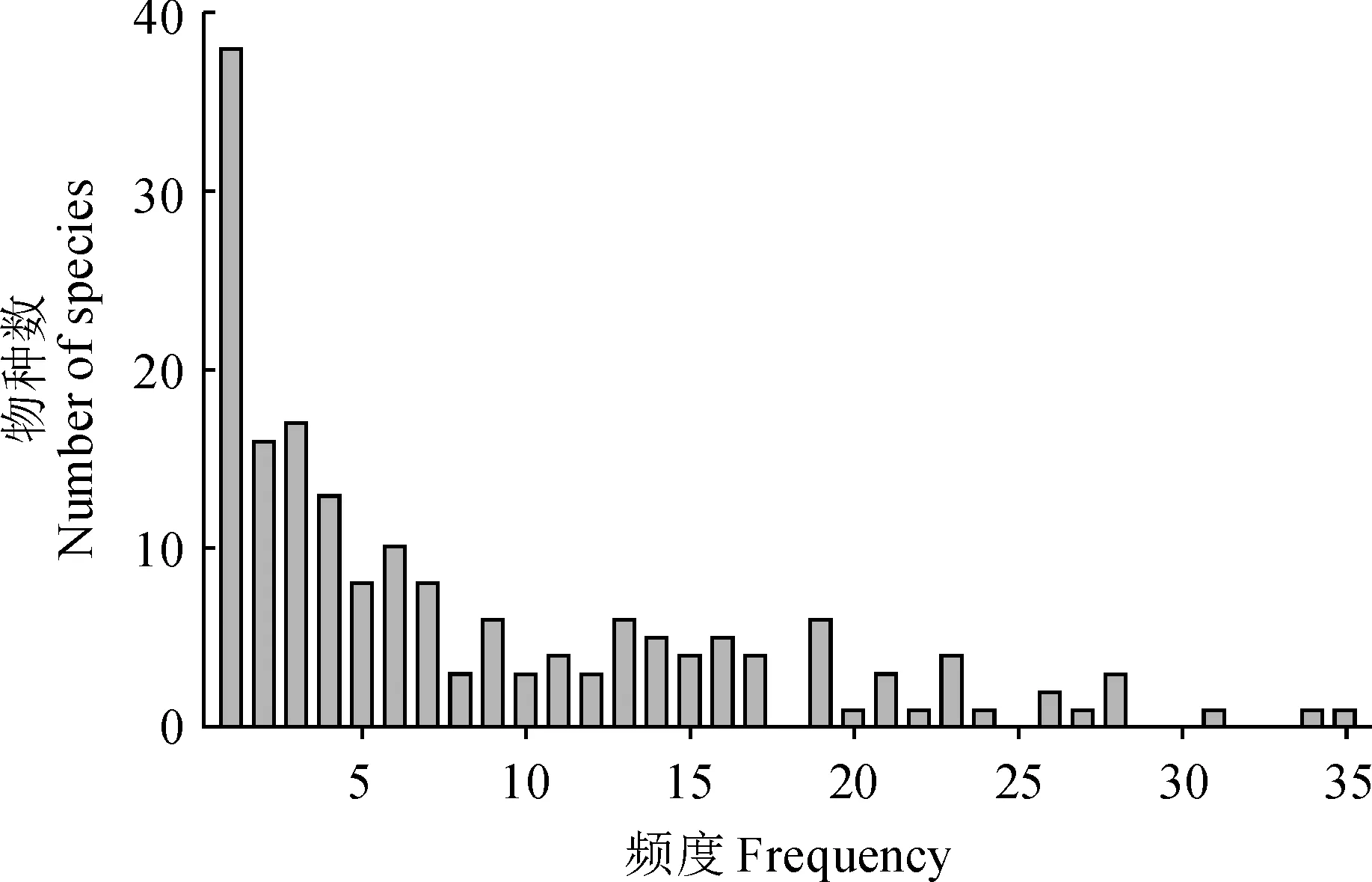
圖2 中條山油松人工林下物種頻度分布Fig.2 Species-frequency diagram for under P. tabuliformis plantations in Zhongtiao Mountain
基于中條山36個樣方林下物種多度數據,繪制稀釋曲線圖(圖3)。由圖3可知,含有最多物種的是樣方22(55個物種),而含有最少物種的是樣方7(19個物種)。為了減小取樣規模對物種數量的影響,將樣本取樣規模標準化,即基于220個個體取樣規模的基礎上比較不同樣方的物種豐富度。標準化后發現5號樣方的物種數最多(45.0個物種),而物種最少的樣方仍為7號樣方(18.9個物種),接近樣方真實物種數據(55,19)。此外,隨著取樣個體數量的增加,5號樣方的物種數增加最快,而21號樣方增加最慢。
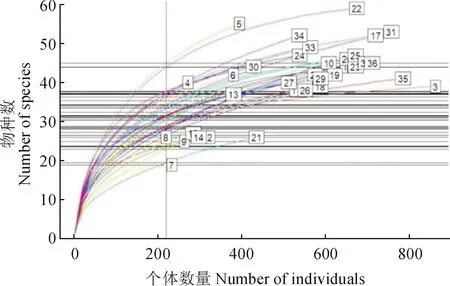
方框內數字(1~36)代表樣方編號圖3 36個樣方林下物種多度稀釋曲線圖The number in the box (1-36) represent the plots numberFig.3 Refraction curves of 36 sample plots
2.2 稀有種與常見種對群落物種豐富度的貢獻
稀有種與常見種對群落物種多樣性的貢獻分析結果(圖4)顯示,稀有種與常見種加入后群落的物種豐富度逐漸增大,其中,常見種加入后,群落豐富度指數呈對數增加,而稀有種加入后,群落豐富度指數呈指數增加,說明常見種對群落豐富度分布格局的貢獻大于稀有種。
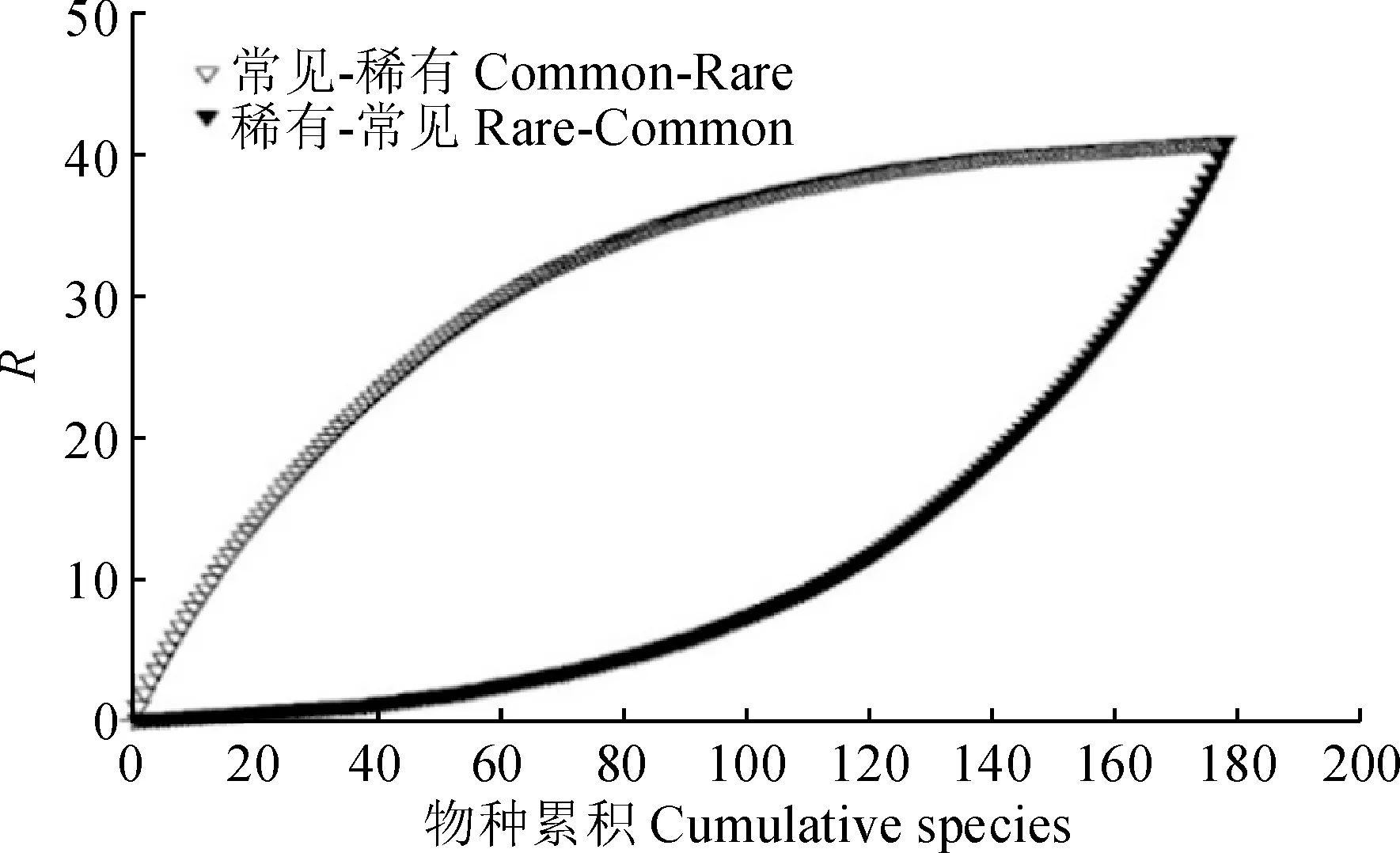
圖4 稀有種與常見種對物種豐富度的貢獻Fig.4 Contribution of rare and common species to species richness
占群落總體物種25%的最稀有/常見的物種(最稀有共54種,最常見共49種)、整個群落(178種)、除25%最稀有/常見物種(分別剩124種、129種)子群落間的相關關系分析結果如表2。由表2可知,最常見的物種解釋了整個群落物種豐富度格局的88.4%(P<0.01),而最稀有物種僅解釋了整個物種群落豐富度格局的24.5%(P>0.05)。去除最稀有后,最常見物種可以解釋剩余物種的90.8%(P<0.01),而去除最常見物種后,最稀有物種僅能解釋剩余物種的48.6%(P<0.01)。進一步說明常見種對物種群落豐富度分布格局的貢獻大于稀有種。
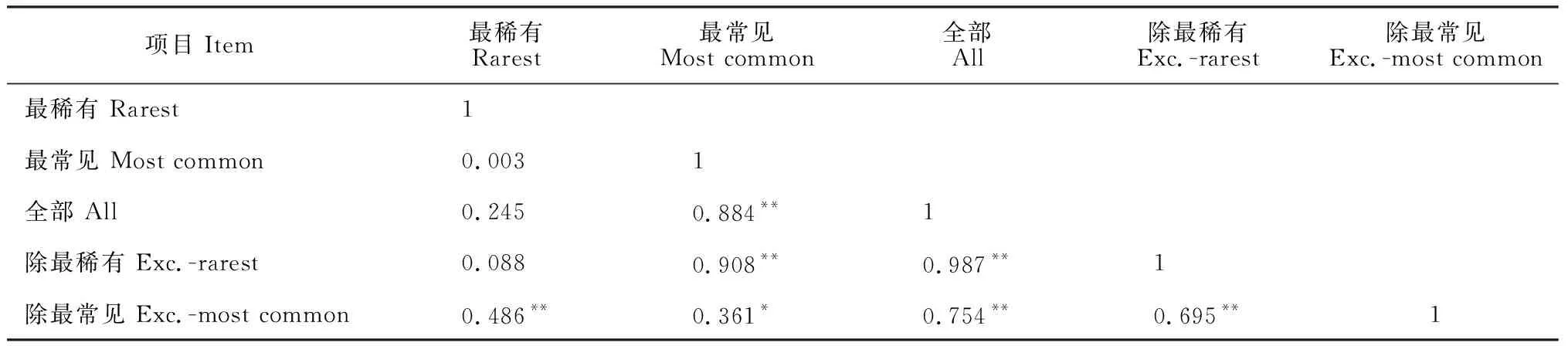
表2 物種豐富度格局間的相關性
2.3 子群落及單一植物種對群落物種豐富度的貢獻
根據子群落與整個群落物種豐富度格局相關分析結果(圖5)可知,當子群落中的“結束”種相對常見而“開始”種相對稀有時,相關性較高,因為整個群落的大部分物種被子群落選取。與直角三角形斜邊平行,子群落的物種豐富度是一個常數,可以看到不同比例的常見物種對稀有物種的影響,即當子群落中常見種越多時,子群落與整個群落的豐富度分布格局相關性越高。
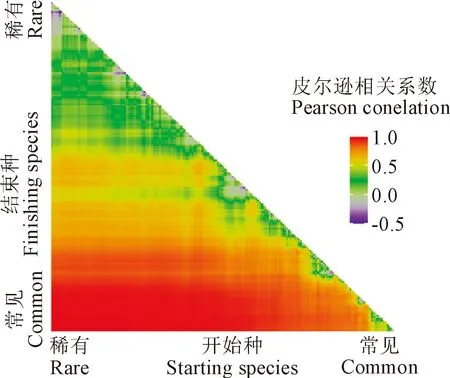
圖5 子群落與整個群落物種豐富度格局的相關關系Fig.5 Correlation between sub-assemblages with the full assemblage species richness pattern
根據子群落與整個物種豐富度格局的相關關系結果,計算添加(或去除)某一物種后子群落與整個群落的相關系數變化量(圖6)。由圖6可知,相關系數變化較大的值均分布于直角三角形斜邊上,特別是直角三角形斜邊的右下方,其值變化值最大,而直角三角形斜邊上的子群落一般由單一物種組成的群落,即一個種組成的群落添加一個物種后,群落的相關系數變化最大,且其呈垂直于橫坐標軸的帶狀分布,即只要有該種存在的子群落,添加(或去除)任意物種后其相關系數變化均最大。按由大到小提取相關系數變化排名前5的物種,即影響該群落物種豐富度分布格局最大的物種,分別為連翹(Forsythiasuspensa)、太平花(Philadelphuspekinensis)、鞘柄菝葜、多歧沙參(Adenophorawawreana)和金花忍冬(Lonicerachrysantha),其中灌木4種,草本1種。將群落物種頻度由高到低排序,以上物種頻度分別為16、23、28、2和15,其中常見種4種,稀有種1種。由此結果可以得出,并非越常見的物種對群落豐富度格局影響越大。
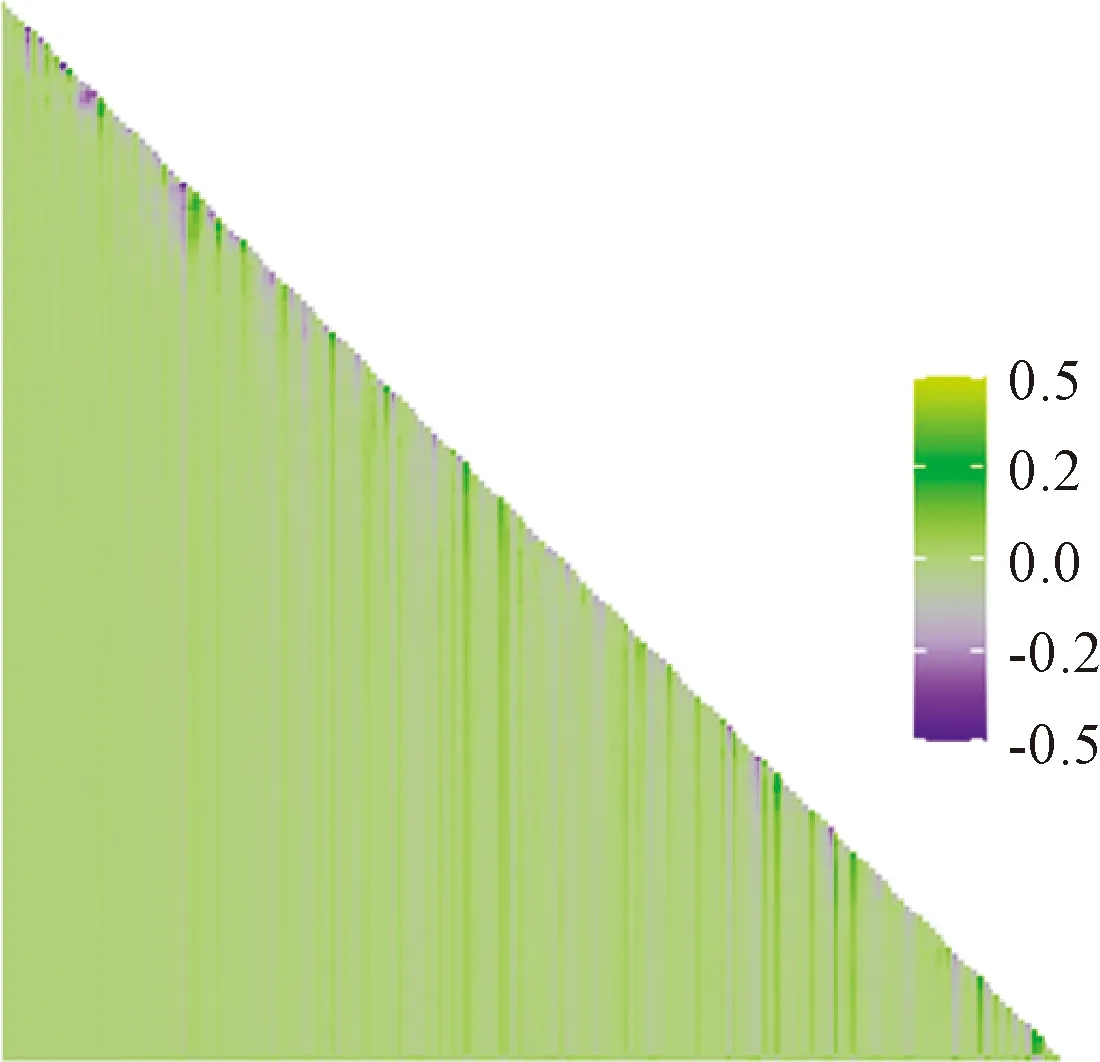
圖6 添加(或去除)某一物種后子群落與整個群落的相關系數變化量Fig.6 The change of correlation coefficient between the sub- assemblage and the full assemblage after the addition (or removal) of a species
以連翹為例,基于Jaccard指數,分析連翹與比其頻度更高的31個物種間的關聯度(圖7)。由圖7可知,相比較于其他物種,連翹與其他物種之間的關聯度最低,即連翹與其他常見種很少出現在同一樣方中。
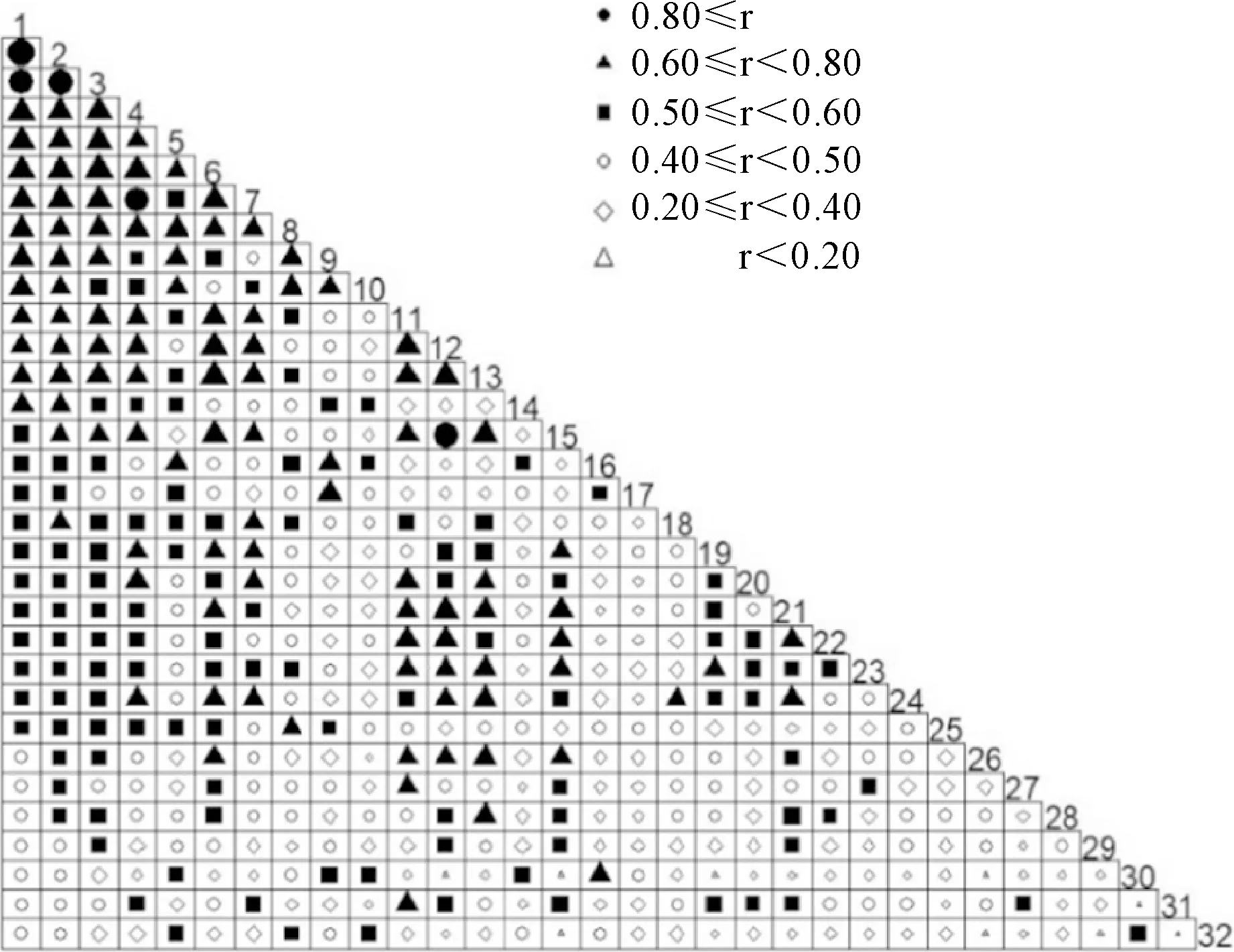
數字1到32物種頻度遞減,32表示連翹圖7 種間關聯分析Numbers 1 to 32 indicate decreasing frequency of species, and 32 means F. suspensaFig.7 Interspecific association analysis
3 討 論
稀有種對群落物種豐富度分布格局的貢獻存在非對等性。稀有種雖然對整個群落功能的重要性可能不很顯著[15],但其豐富度對整個群落豐富度的貢獻很大[9]。例如,本研究發現稀有種雖然對群落物種豐富度分布格局的貢獻小于常見種,但稀有種還是占據了整個群落物種豐富度的30.34%,略低于常見種的33.15%,而群落中維持大量稀有種的主要原因是局域群落之間的遷移與生態位分化,同時也是物種能穩定共存的基礎[29]。
去除稀有種后,剩余群落物種仍能解釋整個群落物種分布信息的98.7%。Schalkwyk等[12]認為群落總體物種豐富度分布格局不反映稀有種,因此在底棲動物生物學評估過程中往往將稀有種按照一定的方式舍去,認為稀有種只能單純反映一些隨機的影響,會對結果產生干擾,而非提供有價值的信息[30],但去除稀有種也可能會損失許多有價值的信息。例如,本研究發現稀有種多歧沙參可能會對群落的物種豐富度格局產生較大的影響。所以在研究中并不能簡單舍去,稀有種與常見種相比其對環境脅迫的耐受性要小[31],如果舍去將無法反映群落是否遭受干擾脅迫,進而對群落穩定評價結果產生不確定性。Mouillot等[32]認為在局域和整體水平,稀有種維持生態系統大多數比較顯著的功能特性,特別是比較脆弱的生態功能。同時,群落中稀有種相對于常見種對環境或氣候變化的響應速度較慢,所以其喪失速率和遷移速率較高[29],進一步說明在生態系統物種多樣性保護方面不能忽略稀有種的重要作用。
群落優勢種均為常見種,而包含常見種越多的子群落與整個群落豐富度分布格局的相關性越高,即優勢種或常見種是群落物種豐富度分布格局指示者[33],相比較于稀有種,常見種受環境擾動的影響更大,因此可以產生最大的變異性[13],但群落基礎物種多樣性的減少可能會同時導致微環境條件下分化的物種的遺傳多樣性降低[34],進而導致群落的遺傳多樣性下降,所以有學者提出應該優先保護常見種[33]。
群落中物種頻度、優勢度的排序與物種對群落豐富度格局貢獻度的排序存在不一致性。本研究發現對群落物種豐富度分布格局貢獻最大的前5個物種無一是優勢種,且并不是分布范圍越廣的物種對群落物種豐富度分布格局貢獻越大,相反那些與頻度較高物種間關聯度低的物種對于群落物種的分布格局貢獻較大,但此方法并不能充分解釋稀有種多歧沙參貢獻較高的原因。補償死亡率假說又稱稀有種優勢(rare species advantage)或者常見中劣勢(common species disadvantage)認為稀有種具有比常見種更高的種群增長率,從而避免其在局域群落種滅絕[7-8],且Connell等[35]用群落補償趨勢(community compensatory trend,CCT)來描述物種增長率與密度的一種負關系,即群落通常并不會通過犧牲稀有種為代價換取常見種的持續增長,從而避免了稀有種在局域群落中滅亡;Mason等[36]認為環境篩的發生是由于物種是稀有種,其個體數太少而不能完全占有所有生境,但稀有種的產生,可能彌補了群落缺失的某些功能,使群落的功能性狀空間分布更加均勻。前人研究結論說明稀有種對群落豐富度格局存在較大貢獻。
本研究基于α多樣性分析了中條山油松人工林下物種對群落物種豐富度分布格局的貢獻,豐富了物種豐富度分布格局的研究,但還存在以下幾點不足。本研究發現稀有種多歧沙參對群落物種豐富度分布格局具有較高的貢獻,且種間關系、補償死亡率假說和環境篩作用并不能完全解釋稀有種貢獻度高的原因,所以今后應對此開展深入研究,以補充本研究的不足;研究生物群落構建的過程中,采用物種豐富度的研究方法無法量化物種在生態策略和生態功能等方面的差異,而且也缺少生物多樣性應包含的其他重要信息[37],而基于功能性狀及功能多樣性的研究方法已成為探索物種共存于生物多樣性維持機制的一個新的突破口[38-40];本研究基于數據分析結果發現稀有種對群落物種豐富度分布格局也存在較大的貢獻,但僅以種間關聯度低來解釋并不可靠。所以要得出更有意義的結論還應該綜合分析其他關鍵生物學信息。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
