水生動物C1qDC蛋白研究進展
2021-09-22 07:51:46高園園孔祥會
水產科學 2021年5期
關鍵詞:功能
高園園,朱 雷,孔祥會
( 河南師范大學 水產學院,河南 新鄉 453007 )
C1q是構成補體系統C1的重要成分,具有與配體結合的“球形”蛋白結構域,在結構上與腫瘤壞死因子十分類似。當C1q與免疫球蛋白IgM或IgG的Fc段結合后,C1q結構發生變化,進而導致C1r和C1s的相繼活化,并啟動補體經典激活途徑[1-2]。C1q還具有清除凋亡細胞、識別病原和調節細胞生長等功能[3-6]。研究人員發現,許多非補體蛋白分子C末端也含有C1q“球形”結構域,并將這些蛋白統稱為C1q結構域蛋白(C1qDC蛋白)[7-8],這些數量龐大的蛋白分子共同組成了一個嶄新的蛋白家族——C1qDC蛋白家族。通過分析C1qDC蛋白在不同種類物種中的分布特點,研究人員認為C1qDC基因起始于原口類動物,并逐漸輻射至脊椎動物[9]。隨著現代分子生物學技術的不斷進步和發展,越來越多的C1qDC蛋白分子在水生無脊椎和低等脊椎動物中被發現和鑒定。研究發現,水生動物C1qDC蛋白在識別病原、抵御病原入侵等先天性免疫活動中發揮重要作用。因此,筆者綜述了水生動物C1qDC蛋白近年來的最新研究進展,以期為水生動物C1qDC蛋白的相關研究提供理論依據。
1 C1qDC蛋白在水生動物中的鑒定及分布特點
C1qDC蛋白在貝類中廣泛分布。通過全基因組和轉錄組序列數據庫分析和挖掘,研究人員在長牡蠣(Crassostreagigas)中鑒定到337個具有完整編碼區的C1qDC基因序列,這些C1qDC基因通常是成簇出現,且CDS區域較為完整,這一現象被認為和基因擴張有關。長牡蠣C1qDC基因組織表達具有顯著特異性,在鰓和消化腺中的表達水平最高[10]。Zhang等[11-12]對生長在河口環境下的長牡蠣基因組數據的研究顯示,C1qDC基因在長牡蠣消化腺中高豐度表達,并推測高豐度表達的C1qDC基因與抵御外界病原體感染有關。C1qDC基因的擴張現象在地中海紫貽貝(Mytilusgalloprovincialis)中亦有發現,地中海紫貽貝中的168個C1qDC基因序列在血細胞、消化腺和鰓中表達水平較高[13]。基于海灣扇貝(Argopectenirradians)EST數據庫,Kong等[14]使用RACE技術鑒定到C1qDC基因全長序列,并發現其在肝胰腺中的表達豐度最高。Zhao等[15]利用RACE技術在三角帆蚌(Hyriopsiscumingii)中克隆鑒定到4個C1qDC基因全長序列,發現其在血細胞、肝胰腺、鰓、外套膜等免疫相關組織中廣泛分布。Zhang等[16]自菲律賓蛤仔(Ruditapesphilippinarum)血細胞cDNA文庫中克隆鑒定到4個C1qDC基因全長序列。鄭哲[17]在馬氏珠母貝(Pinctadafucatamartensii)中克隆鑒定到4個C1qDC基因,發現其在外套膜中表達量較高。
隨著研究的深入,C1qDC基因在魚類中也被陸續鑒定和發現。劉閣等[18]在七鰓鰻(Lethenteroncamtschaticum)肝臟cDNA文庫中克隆獲得19個結構各異的C1qDC基因序列,并將其分成了3個亞家族,為進一步深入研究C1qDC基因的結構和功能奠定了基礎。通過對斑馬魚(Daniorerio)EST、cDNA和基因組數據庫的綜合檢索,Mei等[19]共篩選出52個獨立的C1qDC基因序列,并且該類基因編碼的氨基酸序列結構呈現顯著多樣性。基于EST數據庫,研究人員也從半滑舌鰨(Cynoglossussemilaevis)基因組中克隆鑒定到C1qDC基因,并對其編碼的蛋白功能進行了一系列研究[20]。Nakamura等[21]在蘭氏褐海鯽(Neoditremaransonnetii)血漿中分離鑒定到一種與巖藻糖結合的C1qDC蛋白,并且該C1qDC蛋白序列具有一些保守結構位點。
目前,C1qDC基因已經在多種水生動物中得到鑒定,主要包括長牡蠣[10,22-24]、地中海紫貽貝[13,25]、海灣扇貝[14,26]、櫛孔扇貝(Chlamysfarreri)[27]、三角帆蚌(Hyriopsiscumingii)[15]、七鰓鰻[28]、許氏平鲉(Sebastesschlegelii)[29]、文昌魚(Branchiostomafloridae)[30]、斑馬魚[31]、半滑舌鰨[20]和蘭氏褐海鯽[21]等。C1qDC基因在水生無脊椎和脊椎動物中廣泛存在,并在雙殼類生物長牡蠣和地中海紫貽貝中發生較大規模的基因擴張,其數量遠多于魚類和人類中的C1qDC基因的數量,出現這一現象的原因可能與無脊椎動物缺乏獲得性免疫、抵御外界不良環境和病原的能力較弱有關。
2 C1qDC基因擴張特點
2011年,Gerdol等[13]對地中海紫貽貝進行Sanger測序后,發現并鑒定到168個不同的C1qDC基因序列,隨著新一代測序技術的出現,地中海紫貽貝中C1qDC基因數量增加至232個[32]。2015年,Gerdol等[10]通過深入挖掘牡蠣基因組數據,發現雙殼類動物長牡蠣中C1qDC基因發生了基因擴張。隨著研究的不斷深入,研究人員發現C1qDC基因家族的擴張不局限于地中海紫貽貝中,在長牡蠣和美洲牡蠣(C.virginica)中也均有發生[10,33]。基于此,研究人員推測雙殼類中C1qDC基因發生了大規模的基因擴張事件。
通過對地中海紫貽貝和牡蠣等基因組和轉錄組數據的研究,發現C1qDC基因在擴張過程中存在以下特點:(1)C1qDC基因序列豐度高、多樣性高,C1qDC蛋白結構域呈現高變異性;(2)C1qDC基因在擴張過程中存在碎片化,如馬氏珠母貝有234個C1qDC預測基因序列,但是生物信息學軟件HMMER分析顯示,在其基因組中有多達720個C1qDC基因位點[34];(3)與其他軟體類動物相比(腹足類和頭足類物種),雙殼類動物的C1qDC轉錄本數目眾多,這表明C1qDC基因家族的擴張可能僅限于雙殼類動物。進一步研究發現,C1qDC基因的大規模擴張只發生在雙殼綱的畸形亞綱和異齒亞綱中,而原鰓亞綱和古異齒亞綱中幾乎不存在[10,13]。
3 水生動物C1qDC蛋白的結構及保守性
水生動物中C1qDC蛋白分子結構與人類補體C1復合物的C1qA、B和C肽鏈結構類似但也存在一些不同之處(表1)。根據C1qDC蛋白分子的結構域構成特點[9],發現水生動物C1qDC蛋白主要以C1q-like和球形頭狀C1q (ghC1q)兩種類型存在。

表1 補體C1q分子與C1qDC蛋白結構與功能比較Tab.1 Comparison of structure and function between complement C1q and C1qDC protein
C1q-like蛋白羧基端編碼一個“球形”C1q結構域[9],氨基端存在Gly-Pro-X(其中X可以是任何其他氨基酸)的氨基酸重復序列,可形成膠原樣區。與C1q-like蛋白不同的是,ghC1q蛋白氨基端缺乏膠原樣區,沒有特定的基序。根據氨基末端是否編碼信號肽序列,ghC1q蛋白又可分為頭狀分泌型C1q蛋白(sghC1q)和頭狀胞內型C1q蛋白(cghC1q)。sghC1q型蛋白氨基末端編碼一段信號肽序列,所編碼的蛋白具有胞外分泌能力,cghC1q型蛋白不含信號肽序列,不具有分泌到胞外的能力[9]。據此可知,C1qDC蛋白家族成員呈現出多樣化的特點,這一結果與關鍵氨基酸的變化、膠原樣區的有無以及與其他結構域的組成有關。
進一步分析水生動物C1qDC蛋白的“球形”C1q結構域發現其具有明顯的保守性和高度的序列變異性[13]。對斑馬魚所有C1qDC序列進行多序列比對發現,這些蛋白序列雖然某些位置的氨基酸位點出現較大變異,但其C1q結構域的β折疊處氨基酸位點具有明顯的保守性[19]。長牡蠣的多個C1qDC蛋白在C1q結構域處的氨基酸序列僅有1/4呈現出一定保守性,其余區域顯示出高度變異性[10]。同樣,地中海紫貽貝中C1qDC蛋白序列C1q結構域也呈現高度的序列變異性,僅有少量氨基酸殘基顯示保守性[13]。基于當前的研究結果,筆者將已經報道的水生動物和人類的C1qDC蛋白序列的數目與結構域構成特點進行了歸納總結(表2),可以發現C1qDC蛋白在不同物種中數目差異較大,結構域構成呈現多樣化。
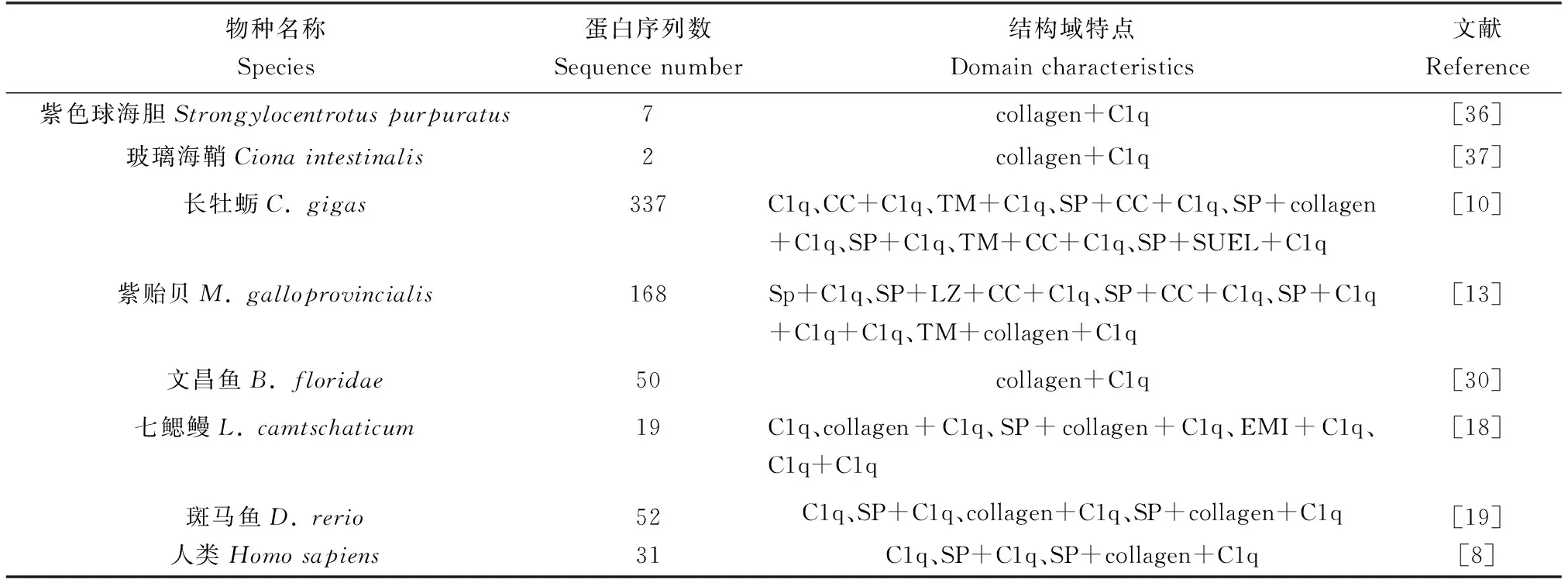
表2 不同物種C1qDC蛋白序列數目與結構特征Tab.2 Number and structural characteristics of C1qDC protein sequences in different species
4 水生動物C1qDC蛋白分類
4.1 貝類C1qDC蛋白分類
貝類C1qDC蛋白存在C1q-like和ghC1q 兩種類型。Gerdol等[10]研究了C1qDC蛋白結構域組成,發現長牡蠣C1qDC蛋白主要存在C1q-like和ghC1q類型,但也發現C1q-like和ghC1q蛋白結構域組成具有多樣性,有些具有膠原樣區和C1q結構域,有些具有與膠原樣區功能類似的卷曲螺旋結構和C1q結構域,有些具有信號肽和C1q結構域,有些僅具有C1q結構域,有些具有SUEL凝集素結構和C1q結構域。SUEL凝集素結構可與D-半乳糖苷/L-鼠李糖結合[35]。Gerdol等[13]根據相同的分類方法對地中海紫貽貝168個C1qDC蛋白序列進行分類,發現地中海紫貽貝中僅1個C1qDC蛋白序列以C1q-like類型存在;其他均以ghC1q類型蛋白存在。另外,在三角帆蚌[38]、櫛孔扇貝[27]等貝類中,C1qDC蛋白均以ghC1q類型存在。
4.2 魚類C1qDC蛋白分類
魚類中C1qDC蛋白也有C1q-like和ghC1q兩種類型。Mei等[19]在斑馬魚基因組數據庫中鑒定到52條C1qDC蛋白序列,根據其N端是否含有膠原樣區和信號肽結構,將斑馬魚C1qDC蛋白主要分為:(1)僅具有C1q結構域以及具有信號肽和C1q結構域的ghC1q類型蛋白;(2)具有膠原樣區和C1q結構域以及具有信號肽、膠原樣區和C1q結構域的C1q-like類型蛋白。劉閣等[18]根據七鰓鰻C1qDC蛋白結構域構成特點將其分為:(1)具有信號肽、膠原樣區和C1q結構域以及具有膠原樣區和C1q結構域的C1q-like類型蛋白;(2)僅具有C1q結構域的ghC1q類型蛋白。Huang等[30]在文昌魚中獲得50個C1qDC蛋白序列,其中42個蛋白為C1q-like類型,由膠原樣區和C1q結構域組成。另外,在半滑舌鰨[20]、許氏平鲉[29]、牙鲆(Paralichthysolivaceus)[39]、蘭氏褐海鯽[21]等魚類中,C1qDC蛋白均以ghC1q類型存在。
4.3 其他水生動物C1qDC蛋白分類
C1qDC蛋白不僅在多種魚類和貝類中被廣泛研究,而且在海膽、海鞘和水蛭(Hirudomedicinalis)等其他水生動物中也被研究和分類。研究發現,紫色球海膽中有7個C1qDC蛋白序列,且均以C1q-like類型存在[36],玻璃海鞘中發現的2個C1qDC蛋白序列也均為C1q-like類型蛋白[37]。此外,Tahtouh等[40]發現,水蛭中C1qDC蛋白同樣以C1q-like類型存在。
5 水生動物C1qDC蛋白的功能特性
5.1 C1q-like類型蛋白功能特性
C1q-like蛋白的免疫功能已在多種水生動物中得到研究。牡蠣C1q-like蛋白不僅具有抗菌活性,且可作為調理素增強牡蠣血細胞的吞噬作用,還能夠介導牡蠣血細胞的遷移[41]。水蛭C1q-like蛋白具有趨化活性,可參與早期受損小神經間質細胞的恢復,在水蛭中樞神經修復過程中起著重要作用[40]。脊索動物海鞘中所發現的C1q-like蛋白具有類似補體C1q分子的作用,推測其可以特異性識別和結合病原微生物表面的寡聚糖結構[37]。文昌魚中C1q-like蛋白可通過與脂多糖相互作用來抑制血小板聚集反應的發生[42]。七鰓鰻中分離出的C1q-like蛋白能夠和N-乙酰葡糖胺相結合,并且可以與絲氨酸蛋白酶-A形成復合體進而裂解七鰓鰻體內的補體C3蛋白[43],推測其可能執行類似于凝集素的功能,參與補體凝集素途徑。斑馬魚中C1q-like蛋白能夠特異性結合斑馬魚免疫球蛋白IgM及人免疫球蛋白IgG和IgM,還能抑制經典途徑引起的溶血反應[44],該結果也表明硬骨魚中可能存在補體經典途徑,而C1q-like蛋白在其中發揮啟動補體經典途徑的作用。通過以上研究可以看出,C1q-like蛋白可以識別細菌、增強細胞吞噬作用以及參與補體途徑,以此在水生動物先天性免疫中發揮重要作用。
C1q-like蛋白不僅具有重要免疫功能,還有一些其他功能。Gerdol等[10]認為長牡蠣中具有卷曲螺旋結構域的C1q-like蛋白序列,可作為膠原樣區結構域的一種替代,協助C1qDC蛋白形成多聚體,由此可見,該蛋白具有組織和穩定多組分C1qDC蛋白復合物的作用[45]。除此之外,C1q-like蛋白還在幼體發育過程中發揮重要調控作用。長牡蠣中C1q39基因在幼體發育的原腸胚到鼓膜幼蟲階段表達水平極高,而在成年組織中表達水平極低[10],說明C1q39蛋白在長牡蠣幼體發育過程中至關重要。
5.2 ghC1q類型蛋白功能特性
ghC1q蛋白分為sghC1q和cghC1q兩類。sghC1q蛋白在雙殼貝類和魚類中廣泛分布,并且在機體先天性免疫過程中發揮重要功能。三角帆蚌中sghC1q基因不僅參與了抗細菌免疫響應過程,而且敲除該基因后會導致免疫基因(TNF和WAP)表達水平降低,表明sghC1q蛋白在免疫信號通路中發揮關鍵作用[15,38]。海灣扇貝中鑒定到的sghC1q蛋白,對細菌和真菌具有明顯的凝集活性,可發揮類似凝集素的功能[14]。研究人員推測在免疫球蛋白進化之前,C1qDC蛋白可以通過凝集素途徑激活古老的補體系統[26]。在長牡蠣[46]、櫛孔扇貝[27]及地中海紫貽貝[13]中,sghC1q蛋白可與多種細菌或病原類似物結合,執行模式識別受體功能。在魚類中,sghC1q蛋白同樣具有模式識別受體功能和凝集素活性。在虹鱒(Oncorhynchusmykiss)[47]、斑馬魚[31]和半滑舌鰨[20]中鑒定到的sghC1q基因可參與細菌刺激魚體后的免疫響應過程。在蘭氏褐海鯽血漿中分離得到的sghC1q蛋白可發揮凝集素活性,且在非還原條件下sghC1q蛋白可以形成多聚體[21]。許氏平鲉[29]中的sghC1q蛋白可作為模式識別受體,發揮結合LPS和PGN的作用,還可結合免疫球蛋白參與補體經典途徑,以及作為調理素增強巨噬細胞對副溶血性弧菌(Vibrioparahaemolyticus)的吞噬活性。sghC1q蛋白不僅具有重要的免疫功能,而且參與機體幼體發育過程。斑馬魚受精后0~48 h,sghC1q基因表達水平受到嚴格調控并呈現出不同的表達模式。其中sghC1q01和sghC1q04以及sghC1q14基因在受精后約24 h開始轉錄,這一時期是咽部形成過渡期,說明其與咽部發育相關[31]。
cghC1q蛋白主要存在于環節動物之前的物種中,最早可追溯到芽孢桿菌屬(Bacillus)細菌[18]。存在于低等生物中的cghC1q蛋白與哺乳動物的細胞周期相關蛋白高度相似,細胞周期相關蛋白能參與細胞增殖等細胞內的許多反應[48]。目前水生動物中關于cghC1q蛋白的報道較少,僅在長牡蠣[49]中鑒定得到cghC1q類型蛋白。該蛋白不僅可作為模式識別受體結合細菌,也可作為調理素來促進長牡蠣血細胞的吞噬作用[49],進而增強長牡蠣血細胞對大腸桿菌(Escherichiacoli)和燦爛弧菌(V.splendidus)的吞噬能力[50]。
根據已報道的水生生物中C1q-like和ghC1q蛋白的研究結果,可以發現兩種類型的C1qDC蛋白均具有識別結合病原、促進細胞吞噬作用以及可能參與補體激活途徑等免疫功能,并且在水生動物幼體發育過程中發揮重要調控作用。
6 總結與展望
補體系統是先天免疫系統的重要組成部分,其組成成分在先天性免疫反應中發揮至關重要的作用[51-56]。C1qDC蛋白作為水生動物補體系統的重要成分,數目繁多,功能多樣[57-60]。C1qDC蛋白不但參與水生動物抵御外界病原入侵的過程,而且具有模式識別和調節補體的功能。隨著測序技術不斷提高,越來越多的水生動物C1qDC蛋白序列被鑒定出來,為該蛋白家族成員的研究工作提供了豐富的數據支撐;既擴展了對該家族蛋白成員的認識,又為深入探索C1qDC蛋白家族蛋白成員的功能和補體系統激活等相關研究提供重要的理論依據。
目前,關于水生低等脊椎動物和無脊椎動物C1qDC蛋白的研究還處于初級階段,尚有許多科學問題值得進一步探索,主要體現于:(1)C1qDC蛋白家族序列在從低等脊椎動物到哺乳類中廣泛存在,但是對于該基因的起源和系統進化關系尚待深入研究;(2)水生動物C1qDC蛋白能與病原或免疫球蛋白結合,這種相互作用已被映射到“球形”C1q結構域的不同結合位點[61],然而,尚未發現促進這種相互作用的特定氨基酸基序;(3)C1qDC蛋白家族成員的功能及其調控的分子機制仍需深入研究。查明C1qDC蛋白家族成員的起源、功能及其調控的分子機制,對于提高水生動物免疫能力和疾病防控水平具有重要理論意義。
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
