寧夏春小麥品種寧春27號成株期抗條銹性遺傳分析
2021-09-23 13:32:34白海波呂學蓮馬斯霜王敬東沈文娟李樹華
種子 2021年8期
關鍵詞:模型
白海波, 惠 建, 呂學蓮, 馬斯霜, 王敬東, 沈文娟, 李樹華
(1.寧夏農林科學院農業生物技術研究中心, 銀川 750002; 2.寧夏回族自治區原種場, 銀川 750200)
小麥是世界上最重要的糧食作物之一,全球35%~40%人口以其為主食[1],我國每年小麥種植面積在2 400萬hm2左右[2]。 由小麥條銹菌專化型(Pucciniastriiformisf. sp.tritici)引起的小麥條銹病是世界性流行的真菌病害,嚴重危害我國廣大麥區[3-4]。小麥條銹病多發于低溫和潮濕環境,從小麥生長出一葉到成熟的過程中都可能遭到侵染[5]。研究表明,利用、選育和種植抗病品種是防治小麥條銹病最經濟有效的方法和措施[6]。

表1 各品種對供試條銹菌系的抗條銹性表現
我國學者在研究小麥抗條銹病方面做了許多工作,鑒定和研究生產上推廣的品種和小麥農家品種。李邦發[7]對西科麥2028抗條銹病進行了遺傳分析;尹軍良等[8]對冬小麥品種天867進行了抗條銹性評價及抗條銹病基因遺傳分析;侯璐[9]在苗期對4份春小麥種質資源進行了條銹病抗性評價和抗性遺傳分析;張調喜等[10]和姚強等[11]分別對青海小麥品種青春38號和青春39號進行了抗條銹遺傳分析;黃苗苗等[12]對甘肅冬小麥品種蘭天23號苗期抗條銹性進行遺傳分析;曹世勤等[13]在苗期和成株期對26個春小麥品種(系)進行了抗性基因分析和評價。寧夏育成的春小麥品種適應性好,在甘肅、青海、新疆等西北地區都有種植,但是育成的抗條銹病品種較少,給新品種的推廣帶來了不利的影響。項目組發現,春小麥旱地品種寧春27號高抗條銹病,本研究主要探究寧春27號的成株期抗條銹病基因Fenix和條銹病抗性遺傳特點,為其在小麥抗病育種中的合理利用提供參考。
1 材料與方法
1.1 試驗材料
以春小麥品種“寧春4號×寧春27號”構建的RILs群體(128個家系)及其親本為遺傳研究材料,RILs材料由本項目組創制并保存。寧春4號[14]是寧夏永寧育繁所選育,于1981年審定推廣。寧春27號[15]是寧夏固原市農科所選育,于1998年審定推廣。
1.2 試驗方法
苗期試驗在甘肅省農業科學院植物保護研究所蘭州溫室進行,成株期試驗分別于寧夏農林科學院農作物所試驗基地和甘肅省農業科學院植物保護研究所甘谷試驗站進行。供試材料按順序編號進行播種。成株期抗病性評價按當地小麥播種適期依次種植在小種圃內。每個材料種1行,行距0.2 m,行長1.0 m,順序排列。苗期試驗于小麥生長1葉1心期采用抖孢子粉法接種。成株期于絕大多數材料旗葉完全展開時采用噴灑孢子懸浮液法進行接種。接種后18~20 d,待感病對照品種銘賢169充分發病后調查病情,RILs群體及親本的條銹病情指標(反應型/嚴重度/普遍率)按照行業標準進行記載[16],調查2次,間隔7 d,以調查中發病最嚴重的數據作為終期病情進行最終統計。
1.3 統計分析
數據采用Microsoft Office 2016軟件進行統計分析。采用植物數量性狀主基因+多基因混合遺傳模型分析軟件[17]分析測定數據。利用極大似然和期望最大化算法估計分布參數,適宜遺傳模型選取AIC值較小且適合性檢驗較優的模型。最適遺傳模型確定后通過最小二乘法計算模型的一階遺傳參數,并計算主基因和多基因的方差和遺傳率。
2 結果與分析
2.1 親本苗期和成株期條銹病抗性統計分析
溫室苗期和田間成株期分小種接種試驗結果(表1)表明,親本寧春4號在苗期和成株期對條銹菌小種和混合菌種均表現為感病,反應型(IT)表現為3~4級,而親本寧春27號在苗期對CYR 32、中4和混合菌表現為感病(IT:3),成株期穩定表現為抗病(IT:0~IT:2)。數據分析顯示,兩親本成株期條銹病抗性比較穩定,寧春4號的嚴重度為40%~80%,普遍率為60%~100%;而寧春27號的嚴重度為0~5%,普遍率為0~20%。結果表明,寧春27號的抗條銹性屬于成株期抗性類型。
2.2 RILs群體成株期條銹病抗性統計分析
RILs群體分別在銀川和甘谷兩個環境中反應型、嚴重度和普遍率的頻數分布見圖1、圖2和圖3。可知RILs群體在兩個環境中各株系間不同條銹病成株抗性評價指標反應型、嚴重度和普遍率的最大值和最小值之間差異明顯,表現出雙向超親分離,且均呈現連續性分布。此外,RILs 群體材料的反應型、嚴重度和普遍率在甘谷都要高于銀川,說明其發病更充分。
2.3 寧春27號的成株期條銹病抗性最適遺傳模型
采用植物數量性狀主基因+多基因混合遺傳分析法,對寧春27號的成株期條銹病抗性基因進行最佳遺傳模型分析(表2),共得到20種遺傳模型,進行適合性檢驗,獲得U12、U22和U32、Wn2和Dn統計量。先篩選出最小AIC值或者接近最小AIC值的模型,再篩選顯著檢驗個數最少的模型確定為最適遺傳模型。因為RILs 群體材料的反應型、嚴重度和普遍率在甘谷均高于銀川,因此以甘谷調查的RILs 群體條銹病指標數據為準進行最優遺傳模型的計算。
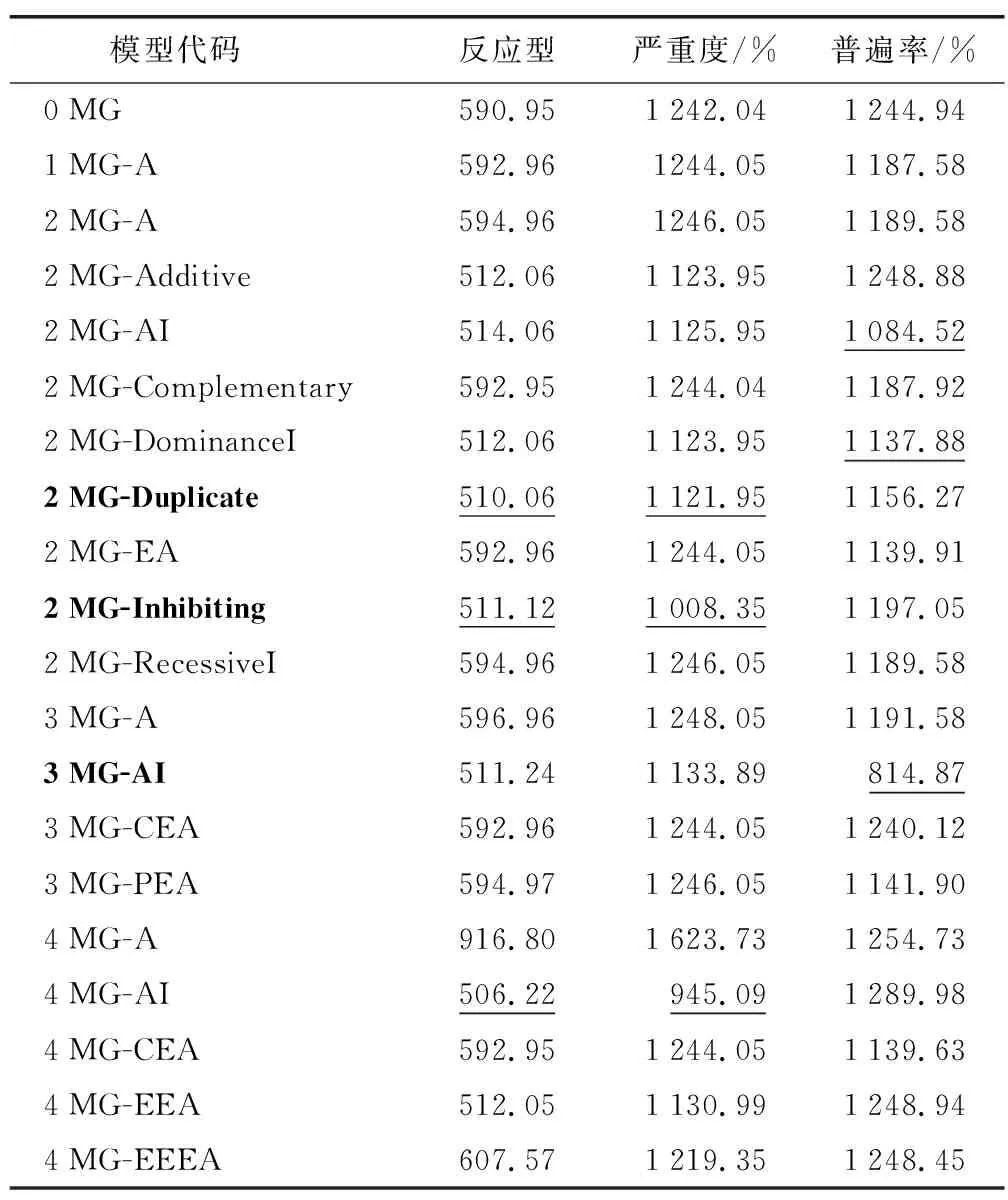
表2 RILs群體抗條銹病指標在各遺傳模型下的AIC值
反應型備選模型中,模型4 MG-AI、2 MG-Duplicate 和2 MG-Inhibiting的AIC值較小,分別為506.22、510.06和511.12,適合性檢驗表明2 MG-Duplicate模型適合性更好,因此,春小麥品種寧春27號對條銹病的反應型符合2對具有重疊作用的加性效應主基因遺傳模型。嚴重度的模型以4MG-AI最小,雖然與模型2 MG-Inhibiting的AIC值相差不大,但適合性檢驗表明,2 MG-Inhibiting模型適合性更好,即為最適遺傳模型。普遍率的模型以3 MG-AI的AIC最小,可作為最適遺傳模型,即3對主基因控制的加性-加性×加性上位性遺傳模型。
2.4 寧春27號條銹病抗性參數


表3 RILs群體在最優遺傳模型下的遺傳參數值
2 MG-Inhibiting模型,即2對具有抑制作用的主基因遺傳模型,2對主基因間加性×加性互作為負向效應(iab<0),主基因遺傳率為83.53%。以普遍率數據分析為3 MG-AI模型,即3對主基因控制的加性-加性×加性上位性遺傳模型,主基因表現為正向加性效應(da>0,db>0,dc>0),其他互作均為負向,主基因的遺傳率為99.90%。
3 討論與結論
小麥條銹菌小種多,且小種變異快,會導致新致病性小種的產生,極易使抗小麥條銹病抗源及抗病性品種丟失抗性,給小麥生產帶來巨大損失。因此今后需要進一步發掘抗病新品種,拓寬抗病遺傳資源,以確保小麥安全生產。繼條銹小種CYR 32、CYR 33流行之后,新的流行性小種是CYR 34[18]。目前,CYR 34即將傳播到我國小麥主產區,將會給小麥生產造成危害[19]。篩選和利用新的抗源及抗病性品種顯得尤為緊迫和重要。國內學者以引進材料、地方品種和育成品種在苗期和成株期抗條銹病基因遺傳分析等方面做了大量工作,研究表明,小麥條銹病抗性分別由1對、2對顯性或隱性基因控制的作用規律[20-24],周春宏等[20]研究表明,曲白春苗期對CYR 33的抗性由1對顯性基因控制,成株期對小麥條銹混合菌種的抗性由2對獨立顯性核基因控制。馬東方等[21]研究表明,小偃6號對條銹小種(CYR 30、CYR 32)的反應型由2對隱性基因控制,對Su 11-4的反應型由1對顯性基因控制。張瑩等[22]研究表明,小麥品系P 9897對CYR 29、CYR 31、CYR 32、CYR 33、PST-Su 5和PST-CH 42混合小種成株期表現高抗,其條銹病抗性是數量性狀,由2對(一顯一隱)基因獨立控制或起重疊作用控制。寧利園等[23]研究結果顯示,紅鎖條對CYR 31和CYR 32的抗病性由2對隱性獨立或重疊遺傳基因控制,對CYR 33的抗病性由1對隱性基因控制;白螞蚱對CYR 31、CYR 32、CYR 33的抗病性分別由2對顯性互補基因、1對顯性基因和2對隱性獨立或重疊基因控制。蘇萍萍等[24]研究表明,P 10078苗期對CYR 32、V 26/CM 42表現為感病,成株期對混合小種表現為高度抗病,其抗性由1對顯性主效基因控制。本研究表明,寧春27號苗期高感,成株期抗病,屬于成株期抗性,與張瑩研究的小麥品系P 9897相同,其抗病性是數量性狀,條銹病抗性指標反應型和嚴重度都是由2對主基因控制。上述研究表明,控制條銹病基因的遺傳特點不同,這種結果差異可能與試驗材料、接種條銹菌小種及鑒定指標等的不同有關。
寧春27號是寧夏固原市農科所選育的春小麥品種,具有高產、優質、抗旱、高抗條銹病、中抗白粉病、感銹及黃矮病等特點,在寧夏南部山區雨養地區大面積種植。本項目組對2011—2017年大田(寧夏銀川、固原)種植的寧春27號進行成株期條銹病調查,發現寧春27號是優良的抗病品種[25],研究寧春27號的條銹病抗性遺傳機制,為今后科學合理利用抗病資源奠定基礎。張薇等[26]對寧春27號進行了條銹病鑒定,但未見關于寧春27號的抗條銹病基因的遺傳規律和遺傳分析研究。因此對寧春27號抗條銹基因進行相關研究,對豐富寧夏培育抗病品種具有重要意義。
本研究對寧春4號與寧春27號及其構建的RILs群體后代進行田間條銹病抗性鑒定試驗。結果顯示,用反應型和嚴重度數據分析得出,寧春27號含有2對成株期抗條銹病性主基因,只是主基因表現的作用方式不一樣。用普遍率分析得出,寧春27號含有3對成株期抗條銹病性主基因。根據寧春27號的抗條銹病的遺傳特點,通過聚合抗病基因,獲得持久抗性的育種新材料,可以用構建的重組自交系群體進一步對抗病性狀進行分析驗證,對抗病基因進一步定位,以期獲得與抗病基因緊密連鎖的分子標記,為分子標記輔助選擇育種提供參考。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
