養豬廢水中磺胺嘧啶對濕地底泥中氮轉化微生物及過程影響
2021-09-24 12:00:58李紅芳張苗苗盧少勇吳金水
環境科學研究 2021年9期
卿 葉, 李紅芳, 張苗苗, 盧少勇, 吳金水, 劉 鋒*
1.中國科學院亞熱帶農業生態研究所, 亞熱帶農業生態過程重點實驗室, 長沙農業環境觀測研究站, 湖南 長沙 410125 2.中國科學院大學, 北京 100049 3.中國環境科學研究院, 湖泊水污染治理與生態修復技術國家工程實驗室, 環境基準與風險評估國家重點實驗室, 北京 100012
隨著畜禽養殖業的迅猛發展,獸用抗生素的廣泛應用和殘留已成為自然環境中抗生素污染的重要來源之一. 其中,磺胺類抗生素因抗菌廣譜、價格低廉等特點,是使用量最大的獸用抗生素之一[1],在畜禽糞污、養殖廢水以及養殖場周圍的地表水和地下水中被頻繁檢測到,且檢出濃度在ng/L~μg/L級別[2-4],最高可達3 630 μg/L[5]. 人工濕地作為一種有效的污水生態處理技術[6],對養殖廢水中NH4+-N、TN的去除率分別為98.8%~99.8%和87.8%~93.8%[7-8],但濕地系統水體氮素的去除是一個微生物參與的過程,任何影響到微生物活性的因素都可能影響濕地的脫氮性能. 而養殖廢水中的抗生素本身就是一種在低濃度下就能抑制或殺死細菌的藥物,因此養殖廢水中的磺胺類抗生素在濕地系統中可能會對濕地的脫氮過程及氮轉化功能微生物產生作用,進而影響濕地對養殖廢水中氮素的轉化和去除.
微生物的硝化、反硝化作用已被證實是人工濕地氮去除的主要機制,可占總去除氮量的60%~90%[9]. 已有研究表明,磺胺類藥物會刺激土壤的氨化與硝化作用,且土壤銨態氮激活率最高可達271%[10],但在100~500 mg/kg濃度范圍內,SD(sulfadiazine,磺胺嘧啶)能顯著抑制池塘底泥氨氮氧化[11]. 另外,SD還會抑制池塘底泥NO3--N(硝態氮)還原,且nirK、nrfA、nosZ基因的相對豐度均低于對照組,但抑制作用隨時間推移而減弱,在30 d后,50和 1 000 μg/L的SD抑制率分別為26%和21%[12]. Chen等[13]也認為,磺胺甲惡唑會對河流沉積物NO3--N還原速率產生顯著影響,且在58 mg/L的高濃度時抑制率為39%,但此時磺胺甲惡唑會促進NO2--N(亞硝態氮)還原,其NO2--N產生率為對照組的7.5倍. 由此可見,不同磺胺類抗生素種類和濃度對氮轉化均存在不同程度的抑制現象,且對氮轉化過程的不同階段或者不同的氮轉化微生物影響有所不同,但目前國內外研究多集中于抗生素對氮素濃度的動態影響,鮮見用氮轉化功能基因和微生物群落的動態變化來研究抗生素對濕地脫氮過程及機制的影響[14-15].
基于人工濕地在畜禽養殖廢水處理的廣泛應用,為探討抗生素的存在對養殖廢水中濕地系統氮轉化的過程及微生物的影響,該研究以典型抗生素SD為研究對象開展室內模擬試驗,通過對氮轉化過程中ρ(NH4+-N)、ρ(NO2--N)、ρ(NO3--N)和ρ(TN)以及底泥中氮轉化功能基因豐度的變化特征,探討養豬廢水中SD對濕地底泥中氮轉化微生物及過程影響,以期為合理設計畜禽養殖廢水高效處理人工濕地提供科學依據.
1 材料與方法
1.1 室內培養試驗設計
試驗設置于中國科學院長沙農業環境觀測研究站黑暗恒溫〔(25±2)℃〕培養室,由5 L塑料桶盛裝2 kg 底泥與3 L養殖廢水混合構建供試濕地系統,其中,底泥采集于定位試驗養殖廢水綠狐尾藻人工濕地處理系統,經馴化后備用;養殖廢水采集于附近養豬場,暴曬1 d后加入SD. 廢水中ρ(TN)、ρ(NH4+-N)、ρ(NO2--N)、ρ(NO3--N)分別為(155.31±3.41)(106.87±8.54)(13.38±0.13)(15.31±3.16) mg/L,ρ(SD)設置為低濃度(10 μg/L)、中濃度(100 μg/L)、高濃度(1 000 μg/L)三組,依次記為SD10、SD100、SD1000,同時設置不加SD空白對照組,記為CK,每個處理組設3個重復. 根據已有研究[16],培養周期設計為28 d,分別在第0、7、14、21、28天采集水樣并當天測定不同形態氮素濃度,同時采集底泥保存于-80 ℃ 冰箱用于DNA提取.
1.2 分析方法
1.2.1水樣分析
定期采集水樣后,用0.22 μm的水系濾頭過濾,然后采用流動注射儀(AA3,SEAL,德國)直接測定ρ(NH4+-N)、ρ(NO2--N)和ρ(NO3--N). 經堿性過硫酸鉀消解后,根據GB 11894—1989《水質 總氮的測定 堿性過硫酸鉀消解紫外分光光度法》,采用流動注射儀測定ρ(TN).
1.2.2底泥微生物氨氧化和反硝化功能基因豐度分析
稱取0.25 g冷凍干燥的底泥樣品,按照Power Soil DNA Isolationkit試劑盒(MoBio,美國)操作說明,提取底泥總DNA. 用Nano-Drop核酸蛋白儀(NanoDropOne,Thermo,美國)和1%的瓊脂電泳測定DNA質量. 將通過質量檢測的DNA樣品于-20 ℃冰箱保存備用.
實時熒光定量PCR(qPCR)反應體系按照SYBR? Premix Ex TaqTM(TakaRa,日本)說明書進行配置. qPCR反應循環條件:95 ℃預變性30 s;隨后95 ℃變性15 s,57 ℃退火20 s(AOA-amoA、AOB-amoA和narG的退火溫度均為55 ℃),72 ℃延伸20 s,反應進行40個循環. 使用已知濃度的質粒載體進行梯度稀釋,并制備標準曲線. 每個樣品3個重復,陰性對照用無菌水. 為消除不同批次擴增的誤差,將每個基因的標準曲線、陰性對照和樣品同時在384孔板上進行擴增,所用儀器為熒光定量qPCR儀(ABI7900,Applied Biosystems,美國),擴增效率從90%到110%且R2大于0.99時默認數據合格. 氮轉化功能基因引物序列信息見表1.

表1 氮轉化功能基因引物序列信息
1.2.3數據分析處理
采用Excel 2013和SPSS 21.0軟件統計分析,Origin 2021軟件作圖. 數據在重復試驗中取平均值和標準誤. 選取單因素方差(One-way ANOVA)分析不同處理組間和組內差異,通過Pearson相關系數分析不同氮素濃度與氮轉化功能基因豐度的相關性.
2 結果與討論
2.1 水體中各氮素濃度及其占比變化
水體中ρ(TN)、ρ(NH4+-N)、ρ(NO2--N)和ρ(NO3--N)的變化見圖1. 在28 d培養期內,ρ(TN)隨時間逐漸下降,去除率為75.4%~80.5%,且添加SD處理組對水體TN去除無顯著影響. 相比ρ(TN),ρ(NH4+-N)隨時間下降更快,但SD1000處理組減緩了ρ(NH4+-N)降速,特別在第7天,4個處理組水體ρ(NH4+-N)的差異最大,SD10、SD100和SD1000處理組NH4+-N轉化率分別為72.8%、59.1%和32.3%,與CK組比,SD100和SD1000處理組NH4+-N轉化抑制率分別達14.0%和53.0%. 這與張敏等[11]在研究SD(100~500 mg/kg)對水產養殖池塘底泥脫氮過程時的結果一致,即SD會顯著抑制NH4+-N的轉化,濃度越高抑制作用越顯著,且SD添加初期抑制作用最顯著.

圖1 ρ(TN)、ρ(NH4+-N)、ρ(NO2--N)和ρ(NO3--N)隨培養時間的變化Fig.1 Changes in the concentration of TN, NH4+-N, NO2--N and NO3--N over incubation time
添加SD與否,對水體ρ(NO2--N)變化無顯著影響,各處理組ρ(NO2--N)均在第14天時迅速下降,到21 d接近0 mg/L,但CK組在28 d略有回升. 添加SD對水體ρ(NO3--N)變化影響差異較大,在培養前期(0~7 d),CK和SD10處理組ρ(NO3--N)很快增至52.5 mg/L,而SD100和SD1000處理組則分別在第14 天和第21天增至52.5 mg/L左右. 這說明SD抑制了ρ(NO3--N)升高,且濃度越高抑制越明顯,其中SD100和SD1000處理組抑制率在第7天達最大值,分別為75.5%和99.5%. Wang等[12]在研究SD對池塘底泥脫氮效果影響時也發現,SD會抑制NO3--N的還原而導致ρ(NO3--N)升高,其中50和 1 000 μg/L的SD對NO3--N抑制率分別為26%和21%.
水體中不同氮素濃度占TN的比例如圖2所示. 在試驗第21天后,各試驗處理組間各氮素組分及占比無顯著性差異,這表明隨時間推移,SD對試驗后期氮轉化的影響逐漸消失. 王燕等[23]通過室內好氣培養法研究SD對水稻土氮礦化影響時也發現,當SD濃度低于2 mg/kg時,隨SD濃度升高,其抑制作用增大,氮凈礦化在前21 d快速降低,在第21~25天降速減慢. 這可能是隨培養時間增加,一方面底泥中SD逐漸降解而濃度降低,或SD作為微生物活動氮源一部分被分解而濃度降低,導致對微生物氮轉化的抑制作用降低[24];另一方面可能是由于在培養期底泥氮轉化相關微生物對SD產生抗性,適應性增強,使得在培養后期SD對氮轉化微生物抑制作用減弱[25].
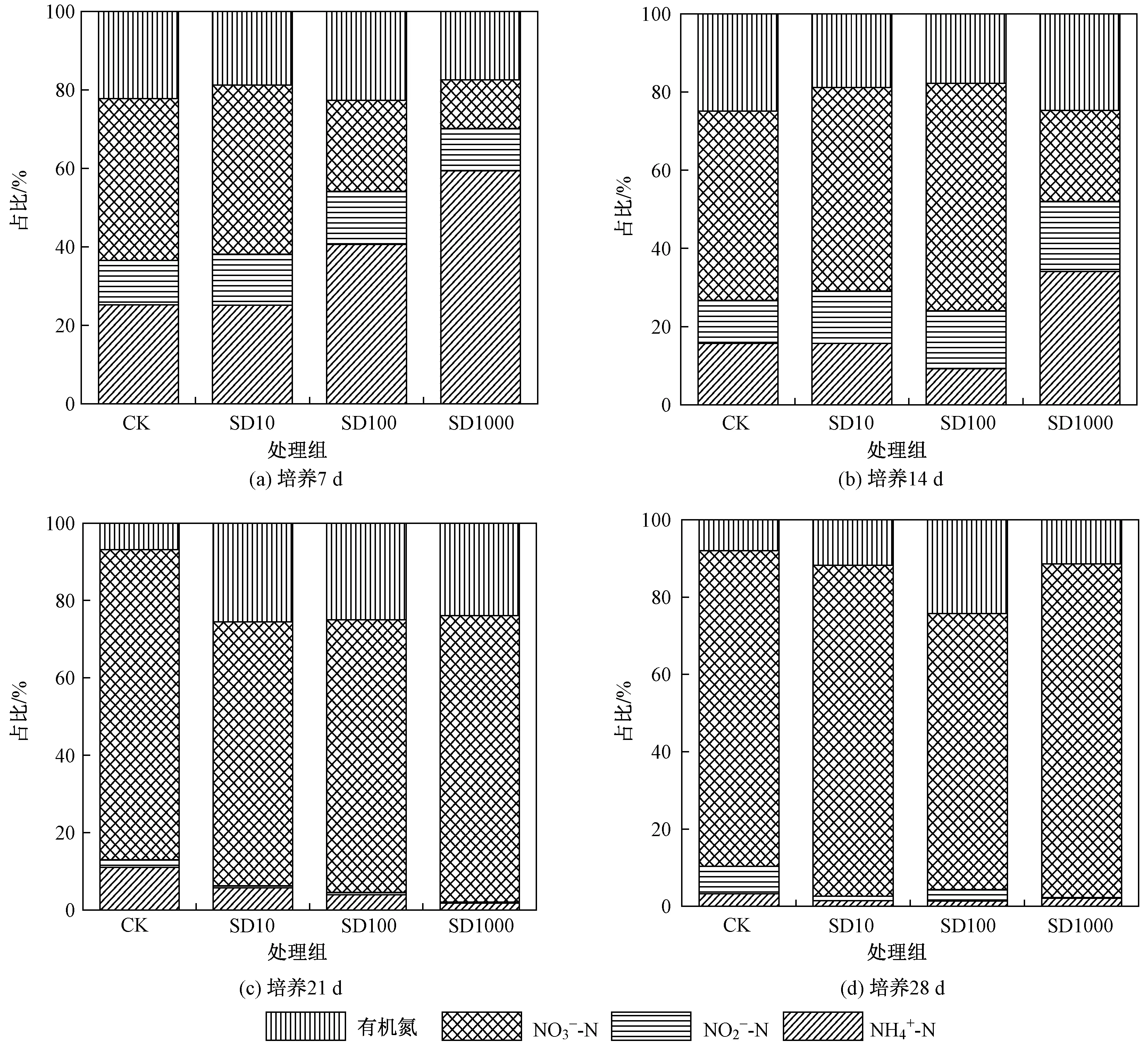
圖2 4個處理組中不同形態氮占TN比例的變化Fig.2 Changes in the proportion of different nitrogen forms to TN in four treatments
2.2 SD對底泥微生物中amoA基因豐度的影響
氨氧化是氮素轉化的關鍵過程,由含氨單加氧酶基因(amoA)的AOA和AOB共同驅動,是硝化作用的限速步驟. 通過qPCR來測定其特異性amoA基因豐度的變化,能有效反映其反應體系中氨氧化微生物AOA和AOB豐度的變化[26].
如圖3所示,底泥中AOA和AOB的豐度分別為5.51×107~3.34×108和1.31×106~1.26×107copies/g. AOA的豐度比AOB高出1~2個數量級,這與Limpiyakorn等[27]研究結果一致,由于古細菌細胞壁和膜的差異,AOA在環境中耐受性較強,分布范圍廣泛,說明AOA可能在氨氧化過程中起主導作用[28].
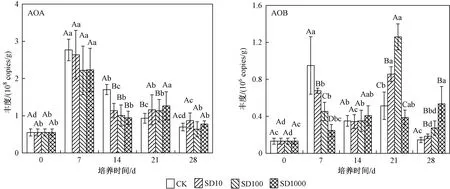
注:不同大寫字母代表同一時間不同處理間差異顯著(P<0.05),不同小寫字母代表同一處理不同時間差異顯著(P<0.05). 下同.圖3 4個處理組的AOA和AOB豐度隨培養時間的變化Fig.3 Changes in the abundance of AOA and AOB in four treatments over incubation time
在第7天,各處理組的AOA豐度達到最高,然后逐漸降低. 相比于CK組,SD添加在14 d時AOA豐度先降低,后略有升高,培養末期(第28天),恢復至與CK組無顯著性差異,SD僅在試驗前期降低了AOA豐度. 這與已有的研究結果相一致,如張敏等[11]對淡水池塘沉積物中AOA和AOB的豐度和群落結構研究時發現,100 mg/L SD能顯著抑制AOA豐度,但在第21天后,各組間AOA豐度無顯著性差異;Liu等[29]也發現,在培養試驗前期,5 mg/L SD能顯著抑制AOA豐度,但在第28天時AOA豐度較CK組無顯著性差異,說明SD對AOA的抑制作用可能會隨著培養時間的增加而減弱.
與AOA相比,添加SD的處理組AOB豐度在第7天抑制效果達到顯著水平,降幅為28.7%~74.0%,AOB豐度隨抗生素濃度升高而顯著降低,Schauss等[30]在研究SD對土壤中AOA的動力學和功能關聯時發現,10和 100 mg/kg SD對AOA的抑制常數均為30 mg/kg,對AOB的抑制常數分別為0.01和10 mg/kg,而抑制常數越大,抑制作用越小,這說明AOB對土壤中SD的抑制更為敏感. 第14天,4個處理組之間的差異不顯著;但到第21天,SD10和SD100兩個處理組的AOB豐度顯著高于CK和SD1000處理組;第28天,SD1000處理組的AOB豐度最高,且隨SD濃度增加AOB豐度也呈現相應的增加趨勢. 這與Wang等[31]研究結果一致,即在50和100 μg/L SD處理下硝化污泥中氨氧化活性受強烈抑制,但在48 h后恢復,AOB能夠克服SD帶來的不利影響致使其豐度上升,且SD濃度越高,AOB豐度增加越多. Yu等[32]也發現,隨時間增加不利因素對AOB的抑制轉變為刺激,從而在培養后期使得AOB數量增加,其原因可能是AOB在SD降解和長期適應中具有了抗藥性[31],可見SD對AOB的抑制作用可能會隨著培養時間的增加而變成刺激作用.
2.3 SD對底泥微生物中narG、nirS、nirK和nosZ基因豐度的影響
微生物反硝化作用是將硝酸根還原成NO2-、NO、N2O或N2的過程,由多種厭氧或兼性厭氧微生物參與的系列酶催化反應組成,包括硝酸還原酶(Nar)、亞硝酸還原酶(Nir)、一氧化氮還原酶(Nor)和氧化亞氮還原酶(Nos)等,相應編碼基因分別為narG、nirK或nirS、norB、nosZ[26].
試驗期間,底泥中4個反硝化功能基因(narG、nirS、nirK和nosZ基因)的豐度分別為1.09×109~2.51×109、8.10×108~5.51×109、2.30×108~2.52×109、5.10×107~2.91×108copies/g(見圖4).narG、nirS、nirK和nosZ基因豐度隨時間變化規律與AOA和AOB類似,都在第7天達到最高值,與CK組相比,添加SD都降低了反硝化基因豐度,但只有SD1000處理組達到顯著性差異. 第14天,4個處理組間的narG和nirK基因豐度差異不顯著;但與CK和SD10處理組比較,SD100和SD1000處理組顯著降低了nirS和nosZ基因豐度. 第21天,添加SD處理總體上增加了反硝化基因豐度,但均未達到顯著水平. 在第28天時,反硝化功能基因豐度均無顯著性差異.
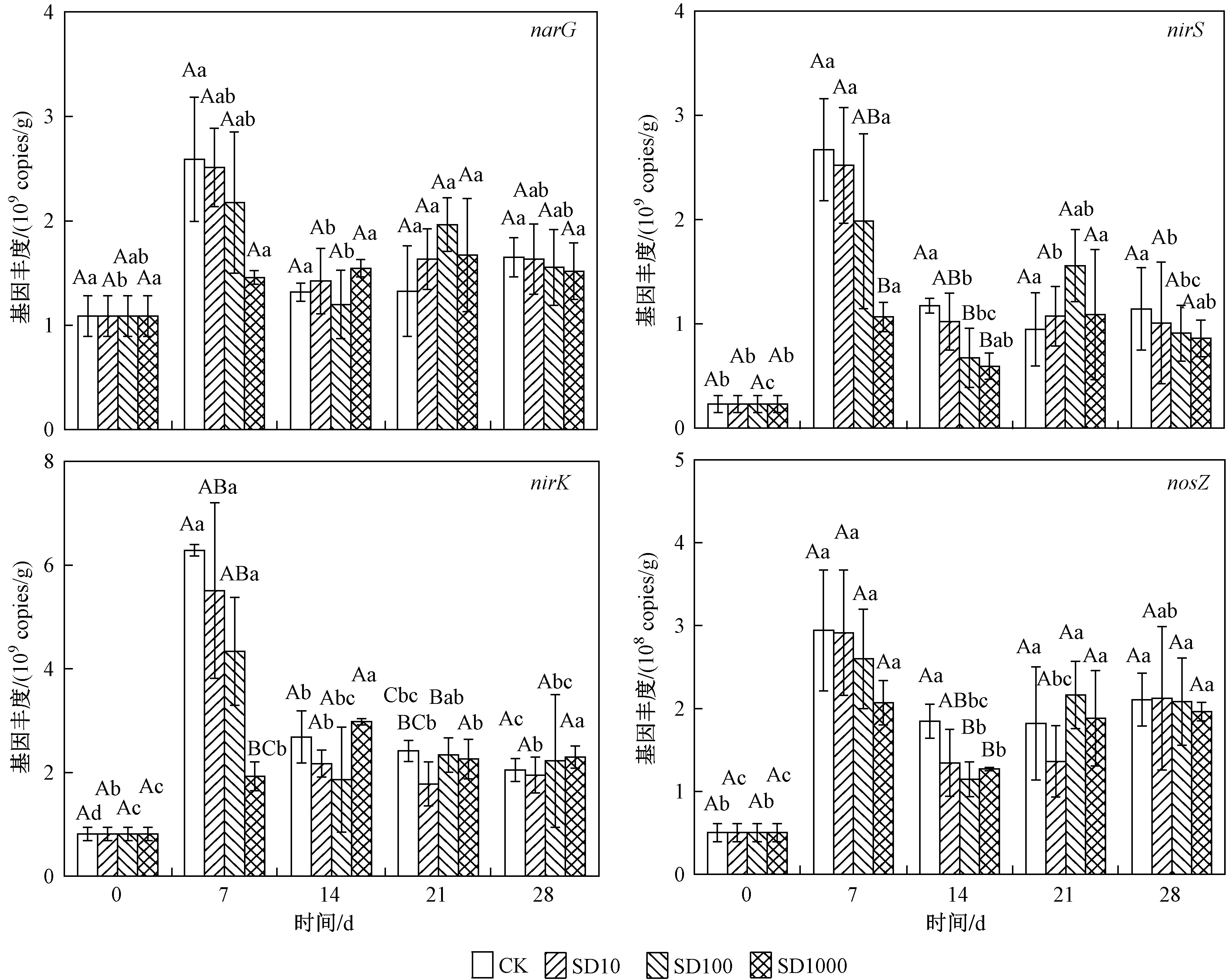
圖4 4個處理組的narG、nirS、nirK和nosZ基因豐度隨培養時間的變化Fig.4 Changes in the abundance of narG, nirS, nirK and nosZ genes in four treatments over incubation time
在培養前期SD添加濃度高于100 μg/L時,narG、nirS和nirK的基因豐度降低,且在SD添加濃度為 1 000 μg/L時,narG、nirS和nirK基因豐度顯著低于其他SD添加組和CK組. Zou等[33]研究地下水中以假單胞菌為主的反硝化菌對各類抗生素敏感性時發現,反硝化細菌對磺胺甲惡唑敏感性的臨界值約為100 μg/L,當磺胺甲惡唑高于100 μg/L時反硝化細菌均被顯著抑制,這表明反硝化基因narG、nirS和nirK對磺胺類抗生素的敏感性臨界值可能具有一致性,值得進一步研究. 在第14天,SD1000處理組的nosZ基因豐度顯著低于CK組,表明高濃度SD抑制了nosZ基因豐度. Wang等[12]研究發現,在池塘底泥中50 μg/L和 1 000 μg/L SD處理組有效降低了nosZ基因豐度;Yin等[34]研究也發現,長江沉積物中磺胺類抗生素對nosZ基因影響最大,具有顯著抑制作用,其原因可能是SD通過抑制葉酸合成抑制大多數革蘭氏陽性和許多革蘭氏陰性細菌的生長[35],從而抑制反硝化菌中nosZ基因豐度增加.
試驗后期反硝化功能基因narG、nirK、nirS和nosZ的豐度均無顯著性差異,這說明SD對微生物抑制作用在后期減弱甚至消失. Liu等[36]發現磺胺甲惡唑在土壤中第7天的降解率達90%,且在試驗前期土壤微生物群落功能多樣性降低,在試驗后期微生物影群落多樣性相應恢復,其原因可能是:①SD降解導致其濃度降低,自身消除了藥效,從而使得抑制強度降低;②隨硝化與反硝化微生物與SD接觸時間增加,微生物體內抗生素抗性基因豐度增加,導致對SD產生了抗藥性. Shan等[37]的研究證實,磺胺類抗生素抗性基因(sul1和sul3)豐度與SD濃度(大于600 μg/L)呈正相關,且磺胺類抗生素抗性基因可能對磺胺脅迫下的脫氮生化途徑形成保護機制.
2.4 水體不同形態氮組分與底泥微生物氨氧化和反硝化功能基因的關聯性
Pearson相關性分析結果(見表2)顯示,AOA豐度與ρ(TN)、ρ(NH4+-N)和ρ(NO2--N)均呈顯著正相關(P<0.05),而AOB豐度與ρ(NO3--N)呈極顯著正相關(P<0.01). 這說明相比于AOB,AOA可能與水體氮轉化過程及氮素形態變化關系更為密切,這與已有的研究結果[30]類似,可能是因為AOA對環境的耐受性更強,分布范圍更廣. 該研究中AOA豐度比AOB高出1~2個數量級,說明AOA可能在氨氧化過程中起主導作用[27]. 反硝化功能基因nirK與ρ(TN)和ρ(NO2--N)均呈極顯著正相關(P<0.01),且nirK、nirS、AOA和AOB四者間均呈極顯著正相關. Liu等[16]研究結果顯示,nirK基因比nirS基因對環境因子的響應更敏感,說明nirK基因在反硝化過程中起著更為重要的作用[38]. 此外,AOA及AOB主導的氨氧化過程,是氮轉化過程的限速步驟,說明可能由于抗生素對AOA和AOB產生影響,進而間接影響反硝化過程. 由此可見,抗生素可能通過直接或間接影響硝化反硝化基因豐度,進而影響養殖廢水中氮的轉化和去除[39],但具體的作用機制還需要進一步研究.
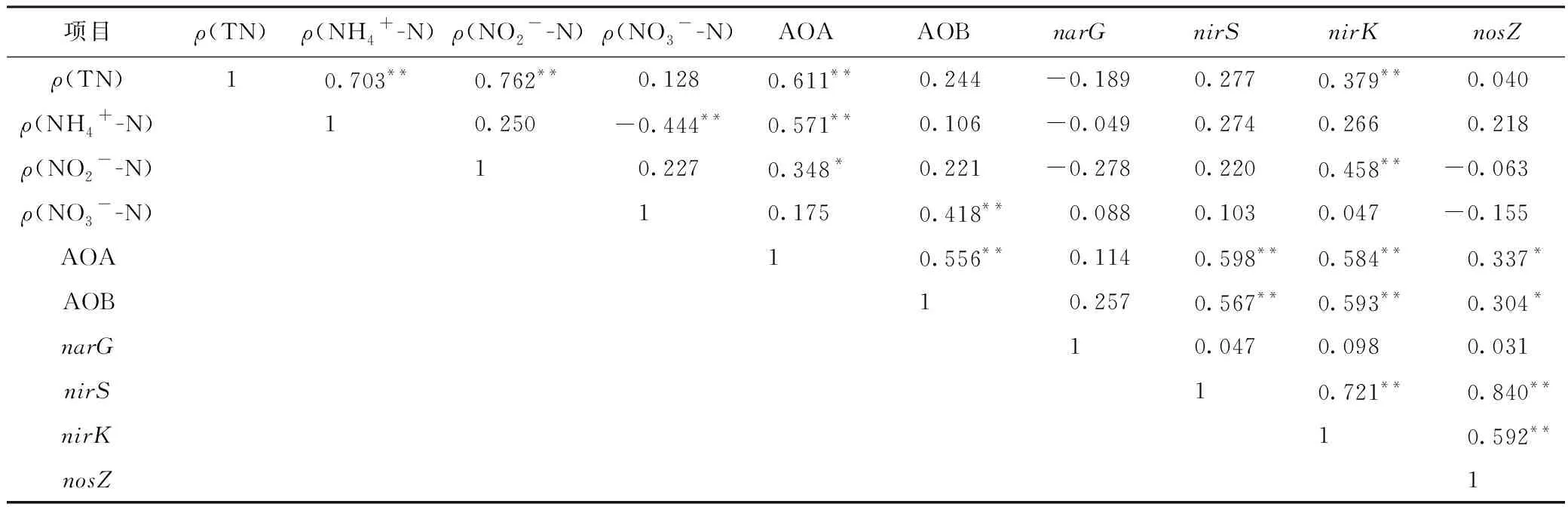
表2 不同氮素濃度與氮轉化功能基因豐度的Pearson相關性
3 結論
a) 室內模擬試驗結果表明,與CK組比較,添加SD處理組對濕地養殖廢水中TN的最終效果無顯著影響,TN去除率為75.4%~80.5%,且到第28天,各處理組中各形態氮素占比無顯著差異;SD對各形態氮的轉化過程產生抑制作用,在第7天抑制效果最強,對水體NH4+-N轉化的抑制率最高達53.0%,對水體NO3--N轉化的抑制率最高達99.5%,且隨著SD濃度的增加抑制作用越強.
b) SD在試驗早期(前14 d)抑制了濕地底泥氨氧化菌和反硝化菌中相關功能基因豐度增加. AOA在氨氧化過程中占主導地位,AOA豐度較AOB高出1~2個數量級,AOB對SD更為敏感,產生抑制作用早于AOA;SD對narG、nirS和nirK、nosZ均產生抑制現象,在第7天和第14天抑制效果最強,與SD濃度呈正相關. 但隨培養時間增加,抗生素抑制作用會減弱,除AOB豐度增加外,各處理組間底泥微生物其他氮轉化功能基因豐度均無顯著性差異.
c) 通過相關性分析表明,AOA與ρ(NH4+-N)呈極顯著正相關(P<0.01),AOB與ρ(NO3--N)呈極顯著正相關(P<0.01),nirK與ρ(NO2--N)呈極顯著正相關(P<0.01),narG、nirS和nosZ與水體中的氮素濃度不顯著相關.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
