牛蒡子苷對三陰性乳腺癌MDA-MB-453細胞增殖和凋亡的影響及作用機制
2021-10-07 13:22:50李霖楓陳重華李天平
中國藥理學與毒理學雜志 2021年8期
關(guān)鍵詞:乳腺癌
李霖楓,陳重華,李天平
(四川大學1.華西藥學院,2.臨床醫(yī)學院,3.華西醫(yī)院,四川 成都 610041)
乳腺癌是全球女性最常見的侵襲性腫瘤,嚴重威脅女性身心健康。《全球癌癥統(tǒng)計報告》[1]顯示,2020年全球新增乳腺癌患者約226萬,發(fā)病率為11.7%,死亡率為30%(68/226)。乳腺癌具有高度分子異質(zhì)性,其中三陰性乳腺癌以雌激素受體、孕激素受體和人表皮生長因子受體2陰性表達為特征,占所有乳腺癌的15%~20%,多發(fā)于絕經(jīng)前女性,癌腫體積大,易發(fā)生轉(zhuǎn)移,惡性程度高,預后相對最差[2-4]。由于缺乏有效治療靶點且對內(nèi)分泌治療不敏感,化療仍是目前針對三陰性乳腺癌唯一有效的全身治療措施,常以蒽環(huán)類、紫杉類和鉑類藥物為首選方案[5-6]。然而,化療藥物存在明顯的不良反應,如心臟毒性、骨髓抑制和神經(jīng)損傷等,造成很多患者治療依從性差,生活質(zhì)量下降;部分患者在治療過程中出現(xiàn)耐藥現(xiàn)象,導致化療失敗[4,7]。據(jù)統(tǒng)計,Ⅱ~Ⅳ期三陰性乳腺癌患者5年生存率明顯低于其他類型乳腺癌[5]。因此,積極尋找有效、低毒的治療藥物是藥物科學家面臨的重要任務。
藥用植物在藥物發(fā)現(xiàn)領(lǐng)域占有重要地位,是新化學實體開發(fā)的重要資源。臨床上約有1/3藥物來源于天然產(chǎn)物或其衍生物,如抗癌藥紫杉醇和抗瘧藥青蒿素。牛蒡子苷(arctiin,ARC)是中藥牛蒡子的主要活性成分之一,具有抗炎、抗病毒、降血糖、抗腫瘤等藥理作用,可用于防治糖尿病腎病[8-9]。研究表明,ARC能有效抑制多種腫瘤細胞生長,如前列腺癌、骨髓瘤、乳腺癌和結(jié)腸癌等[10-12],提示ARC可能是一種潛在的抗腫瘤藥物。Lee等[13]發(fā)現(xiàn),ARC是一種信號轉(zhuǎn)導與轉(zhuǎn)錄激活因子3(signal transducer and activator of transcription 3,STAT3)抑制劑,可誘導多發(fā)性骨髓瘤細胞G2/M期阻滯和凋亡。李孝慶等[14]報道,ARC能降低Bcl-2表達,誘導前列腺癌PC3細胞發(fā)生非凋亡性死亡。為闡釋ARC對三陰性乳腺癌的治療作用,本研究探討ARC對三陰性乳腺癌MDA-MB-453細胞增殖和凋亡的影響及其作用機制。
1 材料與方法
1.1 細胞系
人三陰性乳腺癌MDA-MB-453細胞(中國醫(yī)學科學院基礎(chǔ)醫(yī)學研究所細胞資源中心,資源編號:3111C0001CCC000016):接種于含有10%胎牛血清、1%青-鏈霉素的RPMI 1640培養(yǎng)基中,置于5% CO2,37℃恒溫及飽和濕度細胞培養(yǎng)箱中培養(yǎng)。細胞貼壁生長,穩(wěn)定傳代后,取對數(shù)生長期的細胞進行實驗。
1.2 藥物、試劑和儀器
ARC(成都瑞芬思生物科技有限公司,批號:N-004-100 mg,純度>98%):溶于細胞培養(yǎng)級DMSO配制濃度為100 mmol·L-1儲備液(DMSO終濃度≤0.5%),避光保存于-80℃。實驗時取適量儲備液,用完全培養(yǎng)基配制濃度為1 mmol·L-1的ARC母液。
RPMI 1640培養(yǎng)基和0.25%胰蛋白酶(批號:SH30809.01和SH30042.01),美國Hyclone公司;胎牛血清(批號:04-001-1ACS),以色列BI公司;CCK-8試劑盒(批號:HY-K0301),美國MCE公司;細胞周期和細胞凋亡檢測試劑盒(批號:KGA512和KGA108),江蘇凱基生物技術(shù)股份有限公司;GreenNuc?活細胞胱天蛋白酶3活性檢測試劑盒和BCA蛋白濃度測定試劑盒(批號:C1168S和P0012),上海碧云天生物技術(shù)有限公司;二抗:辣根酶標記山羊抗兔IgG和辣根酶標記山羊抗小鼠IgG(批號:ZB-2306和ZB-2305),北京中杉金橋生物技術(shù)有限公司;一抗:兔抗人周期蛋白依賴性激酶2(cyclindependent kinases,CDK)單抗、兔抗人CDK4單抗、兔抗人細胞周期蛋白D1單抗、小鼠抗人細胞周期蛋白E1單抗、兔抗人P27 KIP1單抗、兔抗人β肌動蛋白單抗(批號:2546T,12790T,2978T,4129T,3686T和4970T),美國CST公司。
倒置相差顯微鏡和OBSERVER D1/AX10 cam HRC倒置熒光顯微鏡,德國Zeiss公司;CO2培養(yǎng)箱,德國Binder公司;EON酶標儀,美國BioTek公司;Cytoflex流式細胞儀,美國Beckman公司;Mini P-4小型垂直電泳系統(tǒng)和Mini TBC小型濕式轉(zhuǎn)印電泳槽,北京凱元信瑞儀器有限公司;ChemiDoc MP化學發(fā)光成像儀,美國Bio-Rad公司。
1.3 CCK8法檢測細胞存活
取200 μL細胞懸液接種至96孔板(每孔3×104細胞),貼壁生長24 h。用不同濃度ARC(100,200,300,400和500 μmol·L-1)處理24,48和72 h,細胞對照組不予藥物干預。加入100 μL CCK8工作液,37℃避光孵育1~2 h,測定450 nm處的吸光度值(A450nm),計算各組細胞存活率。每組設5復孔,實驗重復3次。細胞存活率(%)=(藥物組A450nm-空白組A450nm)/(細胞對照組A450nm-空白組A450nm)×100%。后續(xù)實驗均以48 h為藥物干預時間。
1.4 平板克隆實驗檢測細胞克隆形成率
取2 mL細胞懸液接種至6孔板(每孔2000細胞),貼壁生長24 h。加入ARC 25,50,100和200 μmol·L-1處理48 h,細胞對照組不予藥物干預。撤藥后用新鮮培養(yǎng)基繼續(xù)培養(yǎng),當孔板內(nèi)出現(xiàn)肉眼可見集落時,終止培養(yǎng),用4%多聚甲醛固定,吉姆薩染色液染色15~30 min,計數(shù)克隆,計算各組克隆形成率。克隆形成率(%)=克隆數(shù)/細胞接種量×100%。實驗重復3次。
1.5 流式細胞儀檢測細胞周期分布
參照細胞周期檢測試劑盒說明書進行。取2 mL細胞懸液接種至6孔板(每孔4×105細胞),貼壁生長24 h。加入適量無血清培養(yǎng)基,饑餓處理8 h。藥物干預(同1.3)48 h。收集細胞于2 mL離心管中,350×g離心5 min,用預冷的PBS清洗1~2次,保留細胞沉淀。用300 μL預冷的PBS重懸細胞,緩慢滴加至700 μL預冷的無水乙醇中,混勻后于4℃靜置過夜。洗去固定液,加入300 μL PI/RNA酶A染色工作液,室溫避光孵育30~60 min。上機檢測。利用Modfit LT軟件分析細胞周期分布。實驗重復3次。
1.6 熒光顯微鏡觀察胱天蛋白酶3活性
參照GreenNuc?活細胞胱天蛋白酶3活性檢測試劑盒說明書。取200 μL細胞懸液接種至96孔板(每孔3×104細胞),貼壁生長24 h。藥物干預(同1.3)48 h。PBS洗滌1~2次。取適量1 mmol·L-1胱天蛋白酶3熒光耦聯(lián)底物,用完全培養(yǎng)基稀釋至5 μmol·L-1,每孔加入100 μL,室溫孵育15~30 min,每組設3復孔,用熒光顯微鏡定性觀察綠色熒光并拍照。綠色熒光越多,表明酶活性越強。
1.7 流式細胞儀檢測細胞凋亡率
參照細胞凋亡檢測試劑盒說明書進行。取2 mL細胞懸液接種至6孔板(每孔4×105細胞),貼壁生長24 h。藥物干預(同1.3)48 h。收集細胞于2 mL離心管中,350×g離心5 min,用預冷PBS清洗1~2次,保留細胞沉淀。加300 μL結(jié)合緩沖液,吹打混勻,再加 5 μL Annexin Ⅴ-FITC 和 5 μL PI,室溫避光孵育5~15 min。上機檢測。利用FlowJo V10軟件分析細胞凋亡率。實驗重復3次。
1.8 Western印跡法檢測CDK2、CDK4、細胞周期蛋白D1、細胞周期蛋E1和P27Kip1蛋白表達
收集ARC(100,300和500 μmol·L-1)組和細胞對照組細胞,分別加適量裂解液,冰上裂解30 min,14 000×g離心15 min,收集上清,采用BCA法測定蛋白質(zhì)濃度。調(diào)整各組蛋白質(zhì)濃度,加適量5×上樣緩沖液,95℃變性5 min。SDS-PAGE垂直凝膠電泳(80V→120 V),200 mA冰浴轉(zhuǎn)膜1.5 h,5%脫脂奶粉封閉1 h,一抗(1∶1000)4℃孵育過夜,TBST洗膜3次,二抗(1∶1000)室溫孵育1 h,再次洗膜3次,ECL發(fā)光液顯影。凝膠成像系統(tǒng)掃描條帶,Image J軟件進行積分吸光度值(integrated absorbance,IA)分析,計算各組目標蛋白IA與內(nèi)參蛋白β肌動蛋白IA的比值,表示目標蛋白相對表達水平。實驗重復3次。
1.9 統(tǒng)計學分析
2 結(jié)果
2.1 ARC對MDA-MB-453細胞存活率的影響
CCK8法結(jié)果(圖1)顯示,與細胞對照組相比,ARC 100,200,300,400 和 500 μmol·L-1可 使MDA-MB-453細胞存活率顯著降低(P<0.01),但同一時間點各濃度組間并無明顯差異。然而,在同一濃度下,48 h組細胞存活率明顯低于24 h組(P<0.05,P<0.01),72 h組細胞存活率明顯低于48 h組(P<0.05,P<0.01)。
2.2 ARC對MDA-MB-453細胞克隆形成的影響
平板克隆實驗結(jié)果(表1)顯示,與細胞對照組相比,ARC 25 μmol·L-1組MDA-MB-453細胞克隆形成率明顯降低(P<0.05),50,100和200 μmol·L-1組克隆形成率顯著降低(P<0.01),提示ARC能有效抑制該腫瘤細胞的體外增殖。
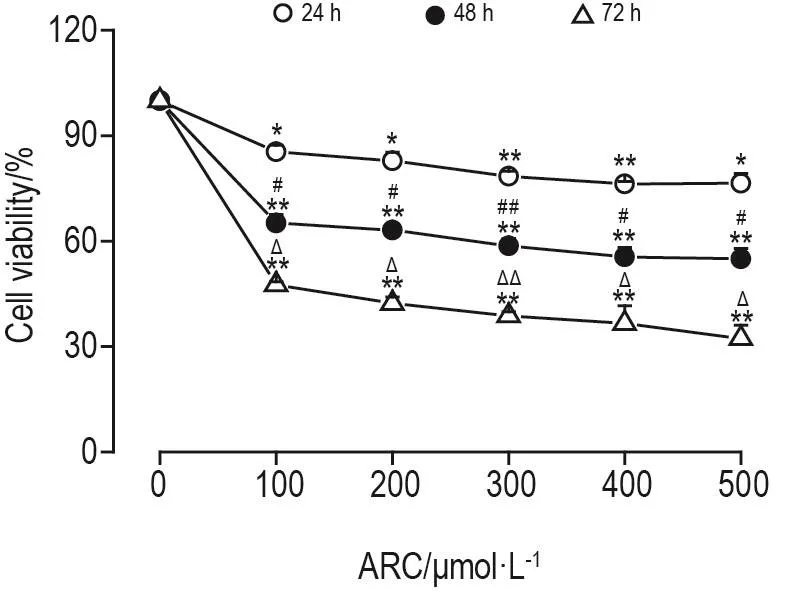
Fig.1 Effect of arctiin(ARC)on cell viability of MDAMB-453 cells.MDA-MB-453 cells were incubated with ARC 100,200,300,400 and 500 μmol·L-1for 24,48 or 72 h.Cell viability was detected by CCK8 assay.±s,n=3.*P<0.05,**P<0.01,compared with cell control(0 μmol· L-1)group;#P<0.05,##P<0.01,compared with corresponding 24 h group;ΔP<0.05,ΔΔP<0.01,compared with corresponding 48 h group.
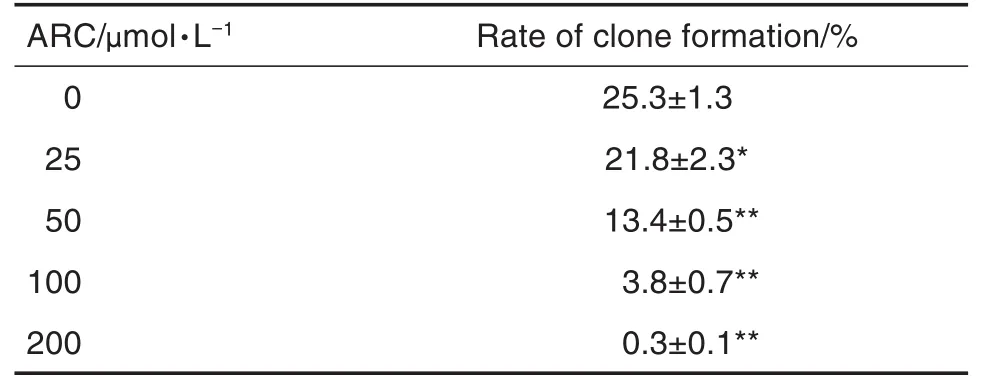
Tab.1 Effect of ARC on clone formation of MDA-MB-453 cells
2.3 ARC對MDA-MB-453細胞周期的影響
流式細胞檢測結(jié)果(圖2)顯示,與細胞對照組相比,MDA-MB-453細胞經(jīng)ARC 100,200,300,400和500 μmol·L-1處理48 h后,G0/G1期細胞百分比顯著增加(P<0.01),S期細胞百分比顯著降低(P<0.01),G2/M期細胞百分比無明顯變化,提示ARC可干預細胞周期進程。
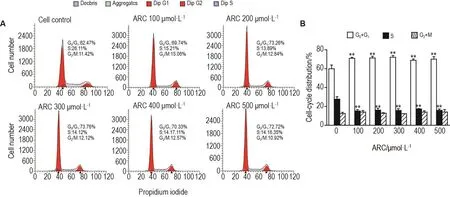
Fig.2 Effect of ARC on cell cycle of MDA-MB-453 cells by flow cytometry.MDA-MB-453 cells were incubated with ARC 100,200,300,400 and 500 μmol·L-1for 48 h.B was quantitative result of A.±s,n=3.*P<0.05,**P<0.01,compared with cell control(ARC 0 μmol·L-1)group.
2.4 ARC對MDA-MB-453細胞內(nèi)胱天蛋白酶3活性的影響
胱天蛋白酶3熒光染色結(jié)果見圖3,細胞對照組處于正常生長狀態(tài),胞內(nèi)胱天蛋白酶3活性低,鏡下幾乎未見綠色熒光;ARC各濃度組鏡下也僅可見零星綠色熒光,提示MDA-MB-453細胞經(jīng)ARC刺激后胞內(nèi)胱天蛋白酶3活性未明顯提高。
2.5 ARC對MDA-MB-453細胞凋亡的影響
細胞對照組和 ARC 100,200,300,400 和500 μmol·L-1組細胞死亡率分別為(5.9±1.6)%,(15.1±1.3)%,(15.1±0.5)%,(12.1±1.1)%,(13.8±2.6)%和(15.0±2.1)%(均P<0.05)(圖4)。與細胞對照組相比,ARC各濃度組凋亡晚期與壞死細胞百分比顯著升高(P<0.01),但早凋細胞百分比無顯著差異,提示ARC可能未誘導該細胞凋亡。
2.6 ARC對MDA-MB-453細胞CDK2、CDK4、細胞周期蛋白D1、細胞周期蛋白E1和P27蛋白表達的影響
Western印跡結(jié)果(圖5)顯示,與細胞對照組相比,ARC 100,300和 500 μmol·L-1組 MDA-MB-453細胞內(nèi)CDK4和細胞周期蛋白D1蛋白水平明顯下降(P<0.01),CDK2和細胞周期蛋白E1無明顯變化;ARC 100和300 μmol·L-1組P27蛋白水平顯著增加(P<0.05)。
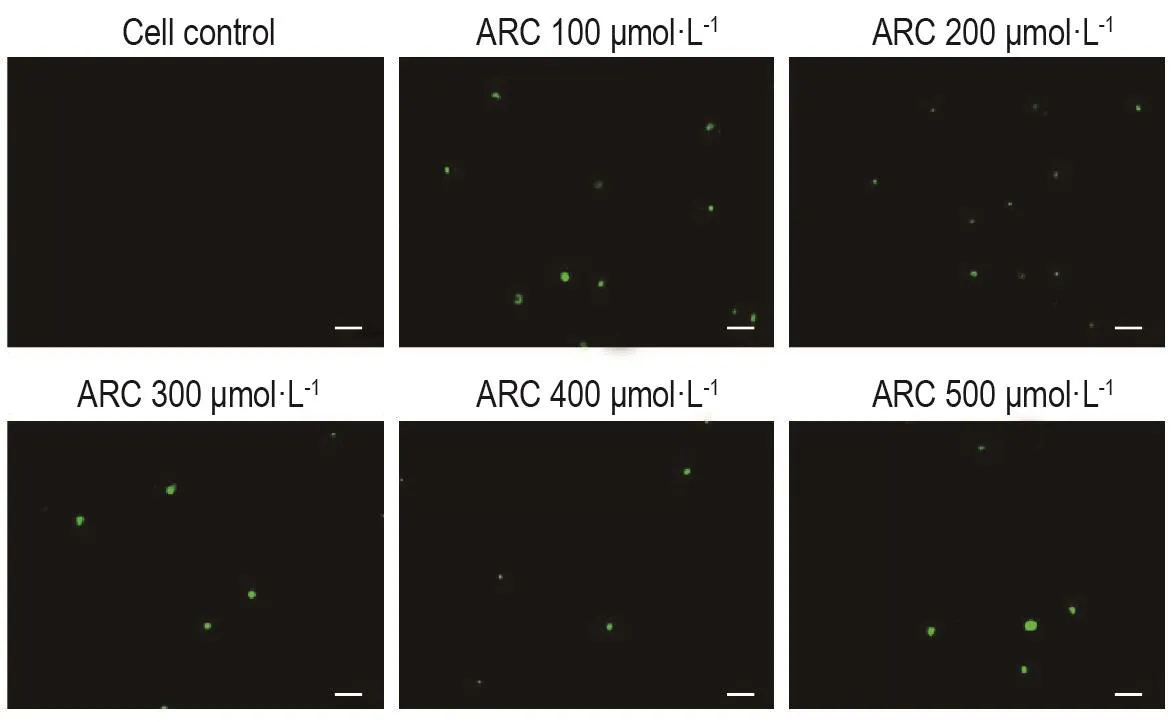
Fig.3 Effect of ARC on activity of caspase 3 in MDA-MB-453 cells.See Fig.2 for the cell treatment.The cells were fluorescent-stained and observed under a fluorescence microscope(GFP channel,×200).The more intense green fluorescence,the stronger the caspase 3 activity.
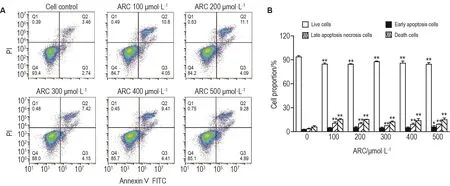
Fig.4 Effect of ARC on apoptosis of MDA-MB-453 cells by flow cytometry.See Fig.2 for the cell treatment.B was quantitative result of A.±s,n=3.*P<0.05,**P<0.01,compared with cell control(ARC 0 μmol·L-1)group.
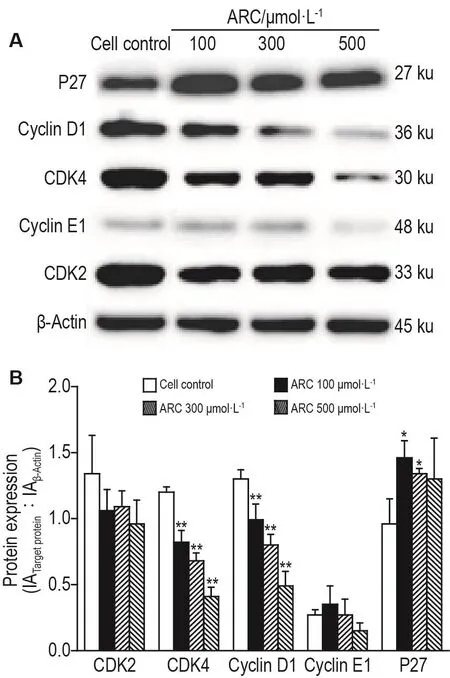
Fig.5 Effect of ARC on protein expressions of cyclin dependent kinases(CDK)2,CDK4,cyclin D1,cyclin E1 and P27 in MDA-MB-453 cells by Western blotting.See Fig.2 for the cell treatment.IA:integrated absorbance.B was semi-quantitative result of A.±s.n=3,*P<0.05,**P<0.01,compared with cell control(ARC 0 μmol·L-1)group.
3 討論
本研究實驗結(jié)果表明,ARC不僅能有效降低MDA-MB-453細胞的存活率,還可顯著減少單個細胞的克隆形成率,提示ARC對三陰性乳腺癌MDAMB-453細胞體外增殖能力具有抑制作用,ARC或能應用于三陰性乳腺癌的治療。
惡性腫瘤的發(fā)生發(fā)展通常與細胞周期異常具有密不可分的關(guān)系。目前,細胞周期抑制劑已成為三陰性乳腺癌治療策略的研究方向之一,如CDK4/6抑制劑(帕博西尼)對Rb基因高表達的三陰性乳腺癌細胞具有明顯抑制作用[15-18]。本研究結(jié)果顯示,ARC干預MDA-MB-453細胞后,S期細胞百分比明顯下降而G0/G1期細胞百分比明顯升高,推測ARC可能干擾細胞周期進程,可導致MDA-MB-453細胞發(fā)生G0/G1期阻滯。細胞周期受到精確且復雜的程序調(diào)節(jié),其中CDK/細胞周期蛋白異源二聚體和CDK抑制因子是推動細胞周期運行的重要因素。細胞周期蛋白D是腫瘤細胞惡性增殖的重要因素之一,在多數(shù)乳腺癌細胞中均高表達[19]。一般而言,細胞周期蛋白D主要在G1期表達,能與CDK4/6結(jié)合形成復合物,磷酸化Rb蛋白,促進E2F轉(zhuǎn)錄因子釋放和一系列相關(guān)基因轉(zhuǎn)錄,協(xié)同CDK2/細胞周期蛋白E推動G1/S期轉(zhuǎn)化[20-21]。P27屬非特異性CDK抑制因子,是G1/S檢查點的關(guān)健分子,能與細胞周期蛋白D競爭性結(jié)合CDK4,導致G1期無法向S期轉(zhuǎn)換[22-23]。CDK4和細胞周期蛋白D表達降低或P27表達升高均會導致G0/G1期阻滯,從而促使腫瘤細胞增殖抑制。既往研究顯示,ARC可通過下調(diào)細胞周期蛋白D1表達以抑制骨肉瘤、肺癌、結(jié)直腸癌和乳腺癌等腫瘤細胞增殖[12]。本研究發(fā)現(xiàn),經(jīng)不同濃度ARC干預后MDA-MB-453細胞內(nèi)P27蛋白水平顯著上調(diào),CDK4和細胞周期蛋白D1蛋白水平明顯下調(diào),CDK2和細胞周期蛋白E1表達未受影響,與先前報道一致[12]。由此可見,ARC可能通過調(diào)控CDK4、細胞周期蛋白D1和P27的表達來干擾MDA-MB-453細胞周期的進程。
細胞凋亡失衡是導致腫瘤無限增殖的另一重要因素,故調(diào)控腫瘤細胞凋亡過程也是腫瘤治療的熱點策略。胱天蛋白酶家族是存在于細胞質(zhì)中一組具有高度同源性的蛋白酶,在細胞凋亡中扮演著重要角色,其中胱天蛋白酶3是細胞凋亡途徑上的終末剪切酶,可作為標志性蛋白反映細胞凋亡狀態(tài)。研究表明,ARC可通過胱天蛋白酶依賴性線粒體途徑誘導多發(fā)性骨髓瘤[13]、肝癌[24]和胰腺癌[25]等腫瘤細胞發(fā)生凋亡。然而,本研究結(jié)果顯示,MDAMB-453細胞在經(jīng)過ARC處理48 h后,胞內(nèi)胱天蛋白酶3酶活化水平和早期凋亡水平均無顯著提高,提示凋亡誘導可能并非ARC發(fā)揮抗腫瘤效應的作用方式。由于ARC組細胞壞死率較細胞對照組有小幅度提升(3.4%→10.0%),推測ARC或許可造成MDA-MB-453細胞非凋亡性壞死,與李孝慶等[14]報道相似,這可能不是造成MDA-MB-453細胞生長抑制的主要原因。
綜上,ARC能有效抑制三陰性乳腺癌MDAMB-453細胞增殖且可干預細胞周期進程,其作用機制可能與下調(diào)CDK4、細胞周期蛋白D1蛋白水平和上調(diào)P27表達有關(guān)。本研究擴展了ARC在抗腫瘤方面的潛在應用,在持續(xù)深入開展機制研究的基礎(chǔ)上,ARC有望開發(fā)為一種靶向細胞周期治療的抗腫瘤藥物。但本研究僅在細胞水平進行了探討,結(jié)果具有局限性,是否存在其他作用機制有待進一步研究。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
現(xiàn)代臨床醫(yī)學(2022年1期)2022-02-12 02:04:58
甘肅科技(2020年20期)2020-04-13 00:30:42
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
幸福家庭(2019年14期)2019-01-06 09:15:38
祝您健康(2018年5期)2018-05-16 17:10:16
癌癥進展(2016年9期)2016-08-22 11:33:20
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:08
