生態系統多穩態研究進展
2021-10-09 01:16:00趙東升張雪梅
生態學報 2021年16期
趙東升,張雪梅
1 中國科學院地理科學與資源研究所,中國科學院陸地表層格局與模擬重點實驗室, 北京 100101 2 中國科學院大學, 北京 100149
由于人類對化石燃料的大規模應用,導致空氣中CO2的濃度迅速升高,使得地球溫度持續上升。IPCC 第五次評估報告指出,在1880年至2012年期間,地球平均溫度上升了0.85℃[1],氣溫升高也導致了極端氣候事件頻率和強度顯著增加。隨著人類對資源的過度開發利用,導致環境破環,湖泊和河流中富營養化、有毒化學物質濃度升高,平原地區地下水位下降,海洋捕撈率上升,森林景觀破碎化、草原過牧和生物多樣性減少等。過去的研究大都認為氣候變化和人類活動對生態系統的影響是漸進的、可預測的和可逆的[2]。但研究發現,生態系統也經常會出現系統性的突變,即在外力的驅動下發生穩態轉換,如珊瑚礁的突然崩潰[3],綠洲突然轉變為沙漠[4],湖泊中突然爆發的水華[5],以及害蟲(蝗蟲)的突然爆發等[6]。漸進影響與突然穩態轉換的相互作用,增加了生態系統響應外界脅迫預測的難度。人類以高效率最佳狀態的驅動力去響應其變化,往往會使得整個生態系統更容易遭受到意外事件的傷害[7]。
目前在微觀水平上對生命的了解正在不斷加深,但是對生態系統響應外界脅迫的穩定性和彈性機理知之甚少。對生態系統突變的研究,國內外學者進行了一些理論探索,但實踐研究還相對較少。生態系統的災難性突變對生態系統結構和功能的穩定性、生態系統服務的持續性和人類生存環境維持起著重要的作用。生態系統一旦發生突變將很難恢復,因此如何利用多穩態理論對生態系統突變進行早期預警,并對退化生態系統恢復提供科學的依據,是國際上生態學研究的熱點[8]。其中彈性的研究受到各國學者們的較多的關注[7],但是對于復雜生態系統突變預警信號的研究仍顯滯后。考慮到目前全球生物多樣性的不斷喪失和生態系統結構和功能退化的情況下,多穩態理論可以為退化的生態系統是否可能在人類干預下進行恢復提供一個關鍵的診斷工具,對生態系統的管理有著重要的意義[9]。
本文通過對多穩態概念的整理得出了一個更加綜合的多穩態定義,并以放牧模型為例,解釋了多穩態理論的相關概念,對生態系統穩態轉換的各個預警信號的數學原理、優缺點和應用條件進行了分析,并對不同尺度下生態系統多穩態的研究方法進行了總結。最后展望了多穩態理論未來研究的問題和重點,為多穩態理論和實踐的深入研究提供科學支撐。
1 生態系統多穩態理論
1.1 多穩態的概念
多穩態(alternative stable states)的概念是1969年由 Lewontin[10]研究“在一個給定的生境中是否會有二個或二個以上的穩定生物群落結構存在?”時提出的。Scheffer[2]和Dakos[3]對多穩態的理解是系統在相同條件下可能收斂到不同的狀態。Clements 和Ozgul[11]則認為多穩態是生態系統的另一種形態,其特征是通過負反饋調節維持其功能和組成的變化。總而言之,多穩態是指在相同的外力驅動或干擾的情況下,其生態系統內生物群落的結構、物質和能量都會發生變化,并且可能表現為由負反饋調節維持的兩種及以上不同的穩定狀態。穩態在生態學領域是指生態系統現有的結構和功能在一定的時間和空間尺度上保持不變。
多穩態理論中的穩態是由負反饋調節所維持的,需要引入一定量的負熵流,才能保持穩定的狀態,而系統突變是由正反饋所驅動的,需要外界條件朝一定的方向持續的變化才會發生穩態轉換[12]。生態系統的演替也可以理解為穩態轉換的過程,生態學家們需要了解生態系統可能存在的不同的狀態和狀態變化是否容易逆轉,以及可能在哪發生轉換,以預測突然變化可能發生的時間和原因。所以,多穩態理論在生態學上有著重要意義,有助于對生態系統狀態變化的預測,并影響對生態系統的管理與決策[8]。
1.2 多穩態理論的闡釋
1.2.1放牧模型
為了深入理解生態系統多穩態系統的動態,需要用數學模型建立一個理論框架來理解其動力學機制。Noy-Meir 在1975開發的牧場放牧模型可以很好的解釋其動力學機制,在這個模型中植被的生長取決于資源的可獲得性,而食草動物的密度是由人類管理者控制的。植被(V)隨時間(t)的動態變化可寫為[13]:
(1)
式中,G(V)為植被生長的函數;c(V)為食草動物(H)消耗植被的函數。當植被(V)隨時間(t)的動態變化率為0時出現平衡,即dV/dt=0,此時G(V)=c(V)H。在圖形上把植被生長函數和消耗植被函數繪制在一起時,交點即為平衡點(圖1)[14]。
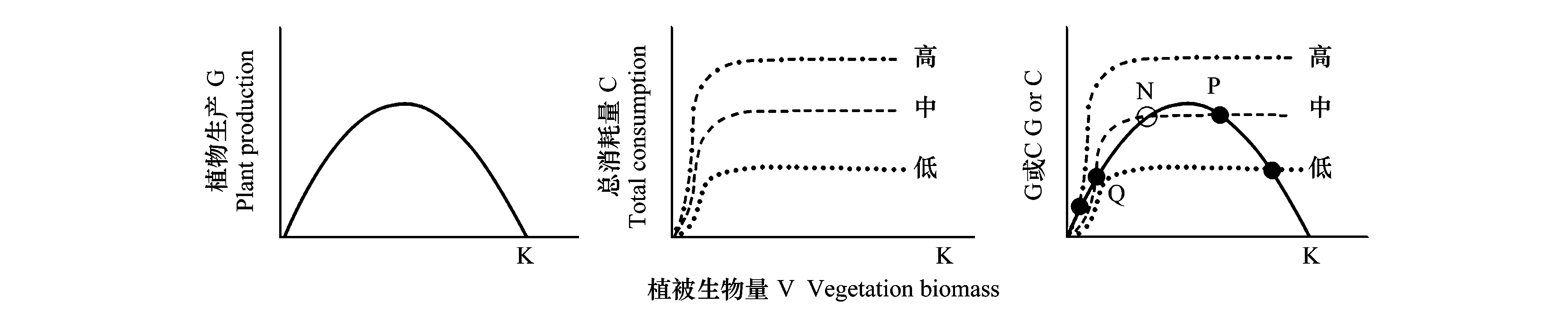
圖1 放牧模型(圖片引用于Beisner[14])Fig.1 Model of the Grazing Animals on Pasture Land
圖1左用Logistic方程作為植被生物量函數,其中K代表環境承載力;圖1中是食草動物對植被生物量的消耗作為植被生物量的函數,三條線分別代表高強度、中強度和低強度放牧情況下植被的總消耗量。圖1右為植物生長和植被消耗的結合,其中黑色的實心點表示穩定的平衡點。在中度放牧的情況下,以P點為例,當V向右移動時(C線>G線),也就是植被的消耗量大于植被的生產量;當V向左移動時(G線>C線),也就是植被的生產量大于消耗量,因此系統總是會返回到原來的平衡點。而空心點N則表示不穩定的平衡點或臨界點。V的輕微增加會使得植被的增長大于消耗(G線>C線),并隨著植被的生長繼續向右移動,直到它們達到平衡點P;V的輕微下降會導致植被的消耗量大于生產量(C線>G線),因此V會進一步向左移動,到達較低的平衡點Q[13-14]。
1.2.2相位空間圖
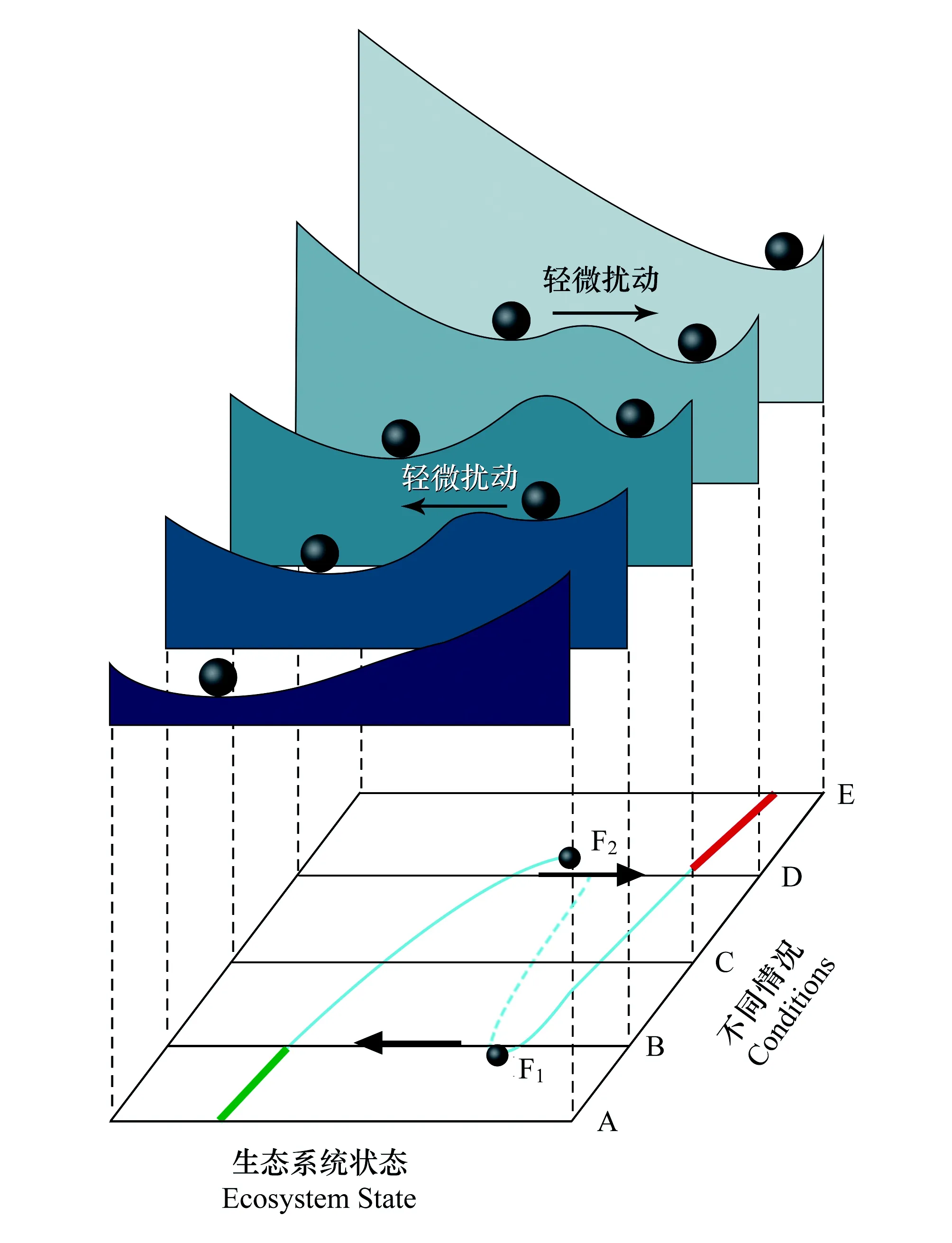
圖2 多穩態相位空間(圖片改編于Scheffer和Carpenter[15])Fig.2 Phase Space of Alternative Stable States
為了更好的理解多穩態理論,可以使用相位空間在V與H的關系圖中繪制所有的可能平衡點(圖2)[15]。圖2中的小球表示生態系統的狀態,更傾向于落在穩定平衡的山谷中。山頂表示不穩定平衡的藍色虛線,并標志著不同吸引盆地之間的邊界。外部條件的變化影響著穩定的格局。當外部條件發生不利的變化時,吸引盆地可能會縮小,當它最終消失時(F2),就會突變到另一個吸引盆地。
圖2中的狀態A是放牧模型中的低強度模式,AB段的綠線為低強度放牧下的穩態,為草原生態系統。BD段的藍色實線為中等放牧模式下的穩態,此時草原生態系統草的數量和質量大大減少。當放牧強度達到高強度時(圖2中的狀態D),輕微的擾動,便會使得草原生態系統轉變為荒漠。DE段的紅線為穩定的荒漠生態系統。其中圖2的F1和F2點為臨界點、閾值或分歧點(Tipping point / Threshold /Bifurcation Point),是指在系統非常不穩定的時候,即使是很小的擾動也會導致狀態發生巨大的變化。從數學上講,當占主導地位的特征值達到零時,系統從一個穩定狀態的吸引盆地通過一個不穩定的區域快速轉移到第二個吸引盆地[11]。指示臨界點到來的一些特征或統計信號被稱為預警信號(Early Warning Signal)。從草原生態系統到荒漠生態系統的轉變稱為穩態轉換(Regime Shift/Critical Transition),是指生態系統由一種穩定狀態向另一種穩定狀態的轉變,由系統的外部強迫引起[11]。
AB段的綠線為低強度放牧下的穩態;BD段的藍色實線為中等放牧模式下的穩態;DE紅線為高強度放牧下的穩態;F1和F2點為臨界點在多穩態的生態系統當中,初始狀態的擾動將決定系統可能達到的穩定狀態。這也稱為“路徑依賴”,足夠嚴重的擾動可以誘導向另一種穩定狀態的轉變。如果當前狀態周圍的吸引盆地較小,則系統更容易轉化到另外一種狀態[15]。其中吸引盆地的大小也稱為彈性(Resilience),通常是指系統在保持功能的同時可以吸收的干擾量。當系統受到壓力時,系統的彈性會發生變化,因為系統的外力會改變吸引盆地的形狀,從而改變系統的彈性[10]。外部條件的變化可能會降低彈性,卻不會對平衡狀態產生較大影響。雖然此時的生態系統看起來沒有改變,但是已經變得很脆弱,很小的擾動都會導致其向另一種狀態轉變[15]。
1.2.3生態系統恢復和演替
從荒漠生態系統恢復到最初的草原生態系統,僅僅恢復崩塌前的條件是遠遠不夠的。如圖2所示需要恢復的程度要遠遠超出分歧點F2,需要多恢復DB距離,直到分歧點F1,在那里系統才會自動突變到之前的狀態。而多恢復的距離DB稱之為滯后(Hysteresis),是指在災難性突變之后缺乏可逆性,系統保持在一個狀態,除非它達到另外一個分歧點才能恢復[11]。但是系統很難恢復到之前的狀態,而是變成了另外一種自組織的狀態,有可能重復上一循環,也有可能進入新的不同的循環(生長、積累、重組和更新),稱為擾沌(Panarchy)[16]。如圖3所示當系統由狀態b恢復到F1點的時候,他可能突變到a,從而恢復到之前的狀態,也有可能突變到e 從而達到另外一種狀態,之后又開始一輪新循環。
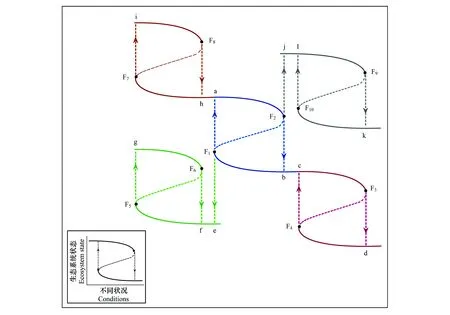
圖3 多穩態與生態系統演替、恢復和擾沌的關系Fig.3 The Relationship Between Alternative Stable States and Ecosystem Succession, Restoration and Panarchya, b, c, d, e, f, g, h, i, j, k, l為不同的生態系統狀態,F1—F10 為不同生態系統穩態轉換的臨界點
用多穩態理論解釋生態系統演替的過程:如圖3所示,狀態i為穩定的生態系統狀態,隨著外界環境條件的變化和內部的競爭到F8時,會突變到狀態h系統; 若環境條件繼續變換,則狀態h會通過穩定的狀態a系統,變化到F2時,之后突變到狀態b系統;若環境條件依舊繼續變化,狀態b 將保持穩定通過狀態c直到F3時,然后突變到d達到另外一種穩定的狀態。從i—h(a)—b(c)—d的過程,是生態系統穩態轉換的過程也是生態系統演替的過程。由內蒙古草原向西北荒漠的演替為例:狀態i為穩定的半濕潤草甸草原,隨著降水量的減少,草甸草原的質量和數量都會大量減少,系統的彈性會降低,直到F8時,系統會由草甸草原突然轉變到半干旱典型草原h(a);隨著降水量繼續減少,典型草原的彈性會下降,典型草原h會通過a漸漸到達F2,之后會突變到干旱荒漠草原b(c);若降水依舊減少,則荒漠草原將會由b通過c,直到F3時,荒漠草原會突變到干旱荒漠狀態d。從半濕潤草甸草原—半干旱典型草原—干旱荒漠草原—干旱荒漠的過程,是生物群落的穩態轉換過程,也是群落演替過程。
2 穩態轉換的預警信號
2.1 臨界減速指標
臨界減速是指在臨界點附近,系統的穩定性下降,導致系統恢復到以前狀態的速度變慢的現象。因為當靠近分歧點時,雅可比矩陣特征值的最大實部趨于零:動力系統從小擾動中恢復的速度越來越慢[17]。其中臨界減速主要的指標有恢復率變慢、自相關、方差和條件異方差的增加[18]。Drake和Griffen為了驗證臨界減速(CSD)發出的預警信號,采取了大型蚤的復制種群進行了實驗,表明由環境條件控制下降引起的穩態轉換的種群,在環境惡化開始之后和臨界轉換之前,表現出CSD的統計特征[19]。
2.1.1恢復率變慢
在發生臨界轉換的分歧點處,占主導地位的實特征值變為零。隨著系統越來越接近分歧點,之前穩定的吸引盆地的斜坡變得不那么陡峭,因此,當系統在向這樣一個臨界閾值移動時,從輕微擾動中的恢復將逐漸變得越來越慢,其速率逐漸變為零。當系統處于穩定的狀態時,即系統的彈性很強,則從擾動中恢復所需要的時間很短;當系統處于臨界轉換的時候,即系統的彈性較弱時,則從擾動中恢復所需要的時間較長。這種現象最直接的含義是,原則上可以通過研究小實驗擾動下的恢復率來探測系統與閾值的距離,這是一種在不破壞系統的情況下探測系統脆弱性的方法[2]。
Veraart等通過在恒化器中培養藍藻,使其暴露在不斷增加的光的脅迫下,并定期進行輕微擾動,測量其藻類的恢復速率,最后表明測量擾動的恢復率是監測臨界減速的穩健方法[20]。Nes和Scheffer也通過生態模型的計算,發現局部擾動實驗的恢復時間可能是過渡前臨界減速的最通用和最可靠的指標[21]。
2.1.2自相關和方差增加(Increase in Autocorrelation and Variance)
在生態系統快要接近分歧點時,自相關和方差都會增加。一個簡單自回歸模型就可以了解到其數學原理[22]。假設每個周期Δt之后存在狀態變量的重復擾動(即增加噪聲)。在干擾之前的恢復速度大約是指數型,且具有一定的恢復速率λ:(公式改編于Scheffer等人[22])
(2)
yn+1=e-λΔtyn+σεn
(3)
這里的yn是狀態變量x偏離平衡點的偏差,εn是來自標準正態分布的隨機數,σ為標準差。若λ和Δt都與yn無關,該模型可以寫成一階自回歸模型:
yn+1=αyn+σεn
(4)
自相關系數α≡e-λΔt,當α=0時為白噪聲,當α=1時為紅噪聲。一階自回歸模型
yn+1=c+αyn+σεn的數學期望為:
(5)
當c=0時,均值為0,方差為:
(6)
當接近平衡時,回歸平衡的速度降低,恢復速率λ趨近于0+,自相關系數α≡e-λΔt趨向于1-,此時方差趨向于無窮大[22]。這就是臨界減速使得方差和自相關系數增加的原因。
方差是一個重要的指標,而且它是一個簡單的特征,理論上可以很容易地衡量。其中Carpenter等[23]研究在湖泊轉變為富營養化條件之前,湖泊水中磷的變化越來越明顯。并且模擬顯示,不斷上升的標準差可能提前大約十年發出早期預警信號。即使并不知道湖泊生態系統動力學的具體情況,通過研究和計算簡單時間序列模型的標準差,也可以預測臨界轉換的到來。然而在某些特定的條件下,在穩態轉換之前,方差可能會下降而不是上升。當環境因素隨機波動,生態系統在臨界值附近對這些因素變得不那么敏感時,或者當嚴重的減速降低了生態系統跟隨環境高頻波動的能力時,就會發生這種情況。此外,當可用數據有限時,由于接近穩態轉換的低頻普遍存在,方差可能被系統性地低估[18]。Veraart等[20]在指標驗證實驗中指出方差的檢驗效果不是很好,甚至呈略微的下降趨勢而非升高。Dakos等[24]也認為并不是所有獨立的潛在的系統轉變都會表現出方差的增長。
相比方差,自相關在穩態轉換時總是增加的。Liu等[25]利用DLM和相關氣候數據,對美國加利福尼亞州林區的歸一化差異植被指數(NDVI)進行了時變自相關估計,表明異常高的滯后一個時間步長的自相關性可作為森林死亡的早期預警信號。Cailleret等[26]也發現方差的增加可能是裸子植物死亡風險的有力預測因子。諸多的實驗和模型模擬也都證明自相關是一個強有力的工具,但是自相關需要長時間且等距的時間序列,并且需要足夠的分辨率來捕獲系統內部動態的特征時間尺度[27]。數據的去趨勢化是至關重要的,因為未經過濾的趨勢可能會導致自組織模式,而這些模式可能會影響擾動系統動態響應的檢測[17]。Dakos等曾使用模型來描述沙漠化過程,在其中一個模型中,放緩未能轉化為方差和相關性的增加。這是由該模型產生規律性的自組織植被模式造成的[28]。這一發現表明了作為臨界轉換指標的方差和自相關的局限性。雖然臨界減速在穩態轉換時可能是一種普遍現象,但在特定系統中通過間接指示器檢測它可能有一定的條件限制。
2.1.3條件異方差
條件異方差是指一個時間步長的方差與一個或多個先前時間步長的方差呈正相關關系。這表明高變化期將傾向于緊隨高變化期,而低變化期將傾向于緊隨低變化期。因為臨近轉變的時間序列的部分將呈現為高變異性的集群,而遠離該轉變的時間序列的部分將呈現為低變異性的集群,所以條件異方差可以作為穩態轉換的預警信號。條件異方差基于拉格朗日乘數檢驗,該檢驗首先將擬合模型的殘差提取到時間序列來計算,然后對殘差進行平方,最后將殘差本身回歸滯后一個時間步長,滯后殘差的線性回歸的正斜率表明條件異方差[29],其一般用于模型模擬研究。
2.2 空間性指標
2.2.1空間相關性
對于具有多穩態的生態系統模型的空間單元,如果假設每個單元的條件不同,那么單元之間的擴散交換將不斷地趨向于減小單元之間的這種差異。準確地說,由反應部分(F)和擴散部分(D)控制的兩個相鄰單元(x1和x2)之間的運動,其方程[18]為:
(7)
(8)
式中,pi為兩個單元之間的異質性的參數,而c是驅動系統到臨界點的控制參數,計算其雅可比矩陣的特征值。當系統遠離臨界點時,單元由“反應”和“擴散”過程支配。當系統靠近臨界點時,因為臨界減速現象,“反應”單元會變得很小,而“擴散”單元與穩態轉換的距離無關,僅取決于兩個單元之間的梯度。此時“擴散”將占主導地位,從而使兩個單元之間的差異相等。導致一個單元中的狀態將強烈依賴于其鄰居的狀態,所以在接近過渡的時候,單元之間將會有更強的相關性[18]。
空間相關性的指標包括空間方差和空間自相關,Donangelo等[30]用空間湖泊模型來研究湖泊富營養化,并且表明空間方差的增加可以作為湖泊從貧營養狀態轉變為富營養狀態的預警信號,其單變量或多變量可觀測的空間方差在作為穩態轉換的預警信號要優于時間方差。Kéfi等[31]利用元胞自動機模型研究了干旱生態系統的荒漠化過程,表明放牧壓力下植被斑塊大小分布的變化可能是荒漠化開始的預警信號。在由許多耦合單元組成的系統中,增強的空間相關性可以作為更強大的預警信號。如果系統中有足夠的空間異質性和連通性,這種空間相關性的增加代表了比時間序列的指標更好的早期預警信號。空間相關性通常要優于時間序列的預警信號指標,這證實了空間指標可能比時間指標更可靠的說法[32]。空間方差和空間自相關的空間模擬在這些類型的空間擴展系統中表現更好[30, 32-33]。
但是這些空間預警信號有3個主要缺點。首先,它們在很大程度上依賴于狀態變量與其空間均值的偏差。因此,當系統接近穩態轉換時,這些指標受到不同位置的狀態變量的時間均值的變化的影響,這很可能發生在不同的異質系統中。其次,這些方法一次只查看一幅影像,因此限制了它們的信息收集能力。例如,僅使用單幅影像很難識別與穩態轉換相關聯的時間模式[34]。而且空間相關性也可能觸發假警報,如果環境中的異質性隨著時間的變化或小規模干擾增加景觀的不均勻性,這可能導致空間相關性的增加,從而產生對即將發生的變化的假警報[32]。
2.2.2空間異方差
空間異方差檢驗首先對數據進行過濾,然后再使用回歸來檢驗過濾值平方之間的自相關,自相關采用Moran′s I指數計算。空間異方差檢驗可以促進使用空間顯性數據,因為它對邊緣的響應很強,特別是在干旱系統的荒漠化方面,因為在干旱環境中,空間相互作用在構建生態系統格局和過程中至關重要[35]。而且空間異方差與臨界減速之間沒有直接的關系,而是對空間變異的聚集性做出響應[36]。此外,空間異方差只需要少量的時間序列數據,并且這些時間的間隔不必有相等的間隔,所以非常適合時間尺度較長的陸地植被系統[25,33],尤其是有觀測誤差和環境特征梯度的實測數據,這一指標很有效。空間異方差也會受到限制,因為這種分析通常不能確定觀測數據中觀察到的模式是由于穩態轉換的生態系統狀態,而不是其他機制。除此之外,它還可能會降低具有高環境噪聲(觀測誤差)的生態系統參數的有效性[35]。
2.3 非臨界減速指標
2.3.1偏度
偏度是多穩態理論中的不對稱指標,受外部大波動和景觀非對稱性對生態系統動力學的影響。偏度信號利用了生態系統的兩個普遍特征:非線性和大的外部波動。無論是彈性降低(動力學參數發生變化)還是外部波動增加(外部噪聲部分的寬度增加),都可能發生穩態轉換。偏度(無量綱)可用τ來表示,其公式[37]如下:P(x)為概率密度函數,μ為均值,σ為標準差:
(9)
式中,P(x)為概率密度函數,μ為均值,σ為標準差。當系統遠離穩態轉換時,偏度為零,偏度越大則越靠近穩態轉換。根據所考慮的生態系統的不同,變化可以由零偏斜到正值或負值,或者從一種偏斜情況到另一個偏斜情況。Guttal和Jayaprakash利用模擬模型和對撒哈拉氣候突變的實測數據進行分析,發現偏度作為穩態轉換的指標是可行的[37]。
2.3.2閃爍
閃爍是指當系統進入導致分歧的雙穩態區域時,如果隨機作用足夠強,開始在兩個可選擇的吸引盆地之間來回擺動[38]。這也是一種早期預警信號,因為如果潛在的條件變化持續存在,則系統很可能永久地轉移到另一種穩態當中。閃爍的一種特殊情況稱為隨機共振,如果隨機擾動與緩慢的周期變化相結合,就會在最終轉變到另一種穩定狀態之前發生一系列周期性轉變[2]。統計上,可以觀察到狀態的頻率分布中的閃爍,表現為方差和偏度的增加以及雙峰(反映了兩種可選的狀態)[39]。
Wang等[40]結合了來自湖泊流域系統的經驗數據和數學模型,表明可以從稀疏的數據中檢測到閃爍,并且如果系統受到足夠大的外部影響,閃爍將成為更強的系統轉變的早期預警信號。Dakos等通過湖水富營養化模型,比較了基于臨界減速指標和閃爍的區別,發現閃爍系統在許多方面與穩態轉換的經典圖像不同,但其動態變化可能會攜帶即將發生穩態轉換有價值的信息[41]。
2.4 生態功能指標
交互網絡可以成為植被變遷的預警信號[42];生產力動態的變化可以成為生物群落尺度上的生態系統狀態轉換的預警信號[43];干旱化的加劇可以成為多種生態系統屬性發生突變的預警信號[44]。植物特性的變化可以使得臨界點更早的到來,也會改變恢復的路徑[3]。基于生物量數據的統計矩陣和體型動態的預警信號可以預測復雜系統的恢復,將這些信號組合成一個單一的度量可以產生最可靠的恢復信號[9]。與協方差矩陣的最大特征值相關的特征向量有助于識別最易受臨界轉變影響的區域,因為它只考慮了系統平衡的波動,從而消除了平衡值的任何變化的影響[34]。
3 不同尺度下生態系統多穩態檢測
在不同的尺度下可以發生不同穩態的轉換。在種群尺度上,恒化器中培養藍藻,藍藻會隨環境的變化迅速轉移到不同的穩態,而且可以采用實驗來測量各個指標[20]。而在群落尺度上,可以從物種和群落水平上測量生物量的時間序列,特有種可能是檢測群落發生穩態轉換的最佳指示物種,將臨界點理論和網絡結構模式相結合可以得出的臨界減速指標[45]。在生態系統尺度上,不同類型的生態系統在不同時間尺度上的穩態轉換也是不同的。在湖泊生態系統中,在日尺度上,浮游生物群落可能會迅速轉移和循環;而在多年尺度上,整個生態系統可能由穩定狀態突然轉化為持續的渾濁狀態。此時湖泊穩態的轉化也可以通過檢測食物網的重組過程來預測[46]。在海洋生態系統中則不能通過相關尺度的實驗來探索。在地球系統中,像厄爾尼諾、PDO等氣候事件幾年就會發生一次;而在更長時間尺度上沙哈拉沙漠的誕生都是突然的穩態轉換[47]。在對大尺度系統的研究時,要通過模型模擬來完成,也必須依賴對自然實驗效果的解釋,對其進行參數的調整。所以尺度越大、系統越復雜,我們對穩態轉換背后的機理的了解就越匱乏,預測也就越困難如圖4所示[2]。
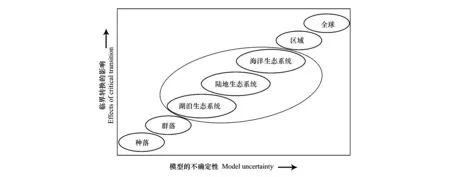
圖4 尺度對模型和穩態轉換影響(圖片改編于Scheffer [2])Fig.4 Effect of Scale on Uncertainty of the Model and Critical Transition
3.1 種群和群落尺度
(1)小型模擬實驗
在種群尺度上,可以通過設置小型的模擬實驗,通過改變外部的環境狀況使系統發生穩態轉換,采用觀測值進行各個預警指標的計算,從而預測或驗證多穩態理論。Veraart 等通過在恒化器中培養藍藻,使其暴露在不斷增加的光脅迫下,并定期進行輕微擾動,通過測量光衰減率來衡量藍藻生物量,并間接測量藍藻的恢復速率。這個模型顯示了多穩態和臨界點,藍藻為自己的生長提供了所需的蔭蔽創造了正反饋,但太高的光強對初級生產力是有害的,這構成了雙穩態背后的機制。最后的實驗結果表明,即測量擾動的恢復率是監測臨界減速的穩健方法[20]。
(2)網絡結構模式
在群落尺度上,可以從物種和群落水平上測量生物量的時間序列,特有種可能是檢測群落發生穩態轉換的最佳指示物種,將臨界點理論和網絡結構模式相結合可以得出的臨界減速指標。Dakos和Bascompte[45]使用79個互利共生網絡的結構來模擬一個環境逐漸變化的場景,該場景第一次導致一個物種的突然滅絕事件,然后是一系列物種的滅絕,直到群落完全崩潰。其結果表明,首次滅絕的物種擁有最強的變異系數,互利共生網絡的嵌套結構會增加穩態轉換的概率。
3.2 生態系統尺度
(1)大型模擬實驗
在湖泊生態系統尺度上,在日尺度上,浮游生物群落可能會迅速轉移和循環;而在多年尺度上,整個生態系統可能由穩定狀態突然轉化為持續的渾濁狀態。此時湖泊穩態的轉化也可以通過檢測食物網的重組過程來預測。Carpenter等在3年多的時間里通過逐漸增加了頂級捕食者來破壞湖泊的食物網,與另一個相鄰的湖泊作為參考生態系統被同時監測。在食物網完成穩態轉換前一年多的食物網重組期間,在實驗組的湖泊中,穩態轉換的預警信號是明顯的[46]。
(2)脆弱性結構分析
在生態系統尺度上可以采用脆弱性的結構進行分析,雖然生態系統的復雜性阻礙了精確的機械建模,但某些特征是脆弱性的通用標志,這些標志通常出現在一些穩態轉換之前。這類系統的總體響應有兩個主要特征:組件的異質性和它們的連接性,這些特性如何影響穩定性取決于網絡中交互作用的性質。單個節點的響應異質性和低連接度可能會導致整個網絡響應環境變化是漸變的而不是突變的。這是因為相對阻力較小,所以不同的節點將會逐個轉移到另一種狀態。相比之下,同質性的節點和高連接度的網絡可能會使得穩態轉換出現阻力,直到臨界點時,其所有的節點都同步移動到另一個狀態[38]。
3.3 區域尺度
(1) 空間代替時間
在有一定梯度的區域可以采用空間替代時間的方法,而且要想用空間代替時間的方法找到穩態轉化的點需要比較苛刻的條件:首先需要一個環境梯度,例如我國的內蒙古地區的草地生態系統的演變是由降水梯度所控制的,并且緯度基本一致,就可以建立一個天然的實驗場。基于這個梯度就可以假設如果將來的降水增多,那么荒漠草原就可能會轉化成典型草原,典型草原就可能轉化成草甸草原;如果將來的降水減少,那么典型草原可能會轉化成荒漠草原,草甸草原可能會轉化成典型草原。Hu等人基于中國內蒙古地區由東到西降水梯度遞減的事實,通過變異系數、不對稱性和對降水敏感性這3個指標,量化了地上凈初級生產力(ANPP)的動態空間格局,并把干旱荒漠草原、半干旱典型草原到半濕潤草甸草原的過渡聯系起來。其表明這3個ANPP動態指標均表現出明顯的空間分布特征,并呈現出草地類型間的峰值信號轉換。因此,監測ANPP動態變化有可能預測未來生態系統狀態的變化。但是,使用空間代替時間的方法來預測生態系統穩態轉換對氣候變化的響應是具有局限性的。例如在土壤屬性上,氣候變化、植被動態和響應之間存在時空不一致性[43]。
(2) 衛星遙感監測
在區域尺度上可以使用衛星遙感監測的方法來驗證或預測穩態的轉換。具體有兩種方法:第一類是遙感影像目視解譯與計算機自動分類相結合的方法;第二類是基于遙感或其所衍生的指標,利用數理統計進行時間序列對比分析[8]。例如Hirota等利用 MODIS(Moderate Resolution Imaging Spectroradiometer)的植被覆蓋度產品數據揭示了非洲、澳洲和南美洲的熱帶和亞熱帶地區存在著三種穩態,即森林態、稀樹草原態和無樹態[48];Miller分析了美國科羅拉多高原半干旱區草原的地面樣方數據,發現該區存在生物結皮-多年生草地態、裸地-多年生草地態以及裸地-單年生草地態[49]。Berdugo等結合地面樣點數據和來自 Google Earth的高分辨率衛星數據,發現旱區在全球尺度上存在著高功能區和低功能區兩種狀態[50]。
3.4 全球尺度
全球尺度的研究大部分采用模型模擬或者全球實地調研和遙感觀測相結合的方法。在古氣候變化和物種大滅絕的研究中,最好的方法就是利用模型模擬古氣候事件發生的時間序列,再進行穩態轉換的驗證。古氣候突變事件包括:大約5000年前,撒哈拉突然從一個有湖泊的植被繁茂的區域變成了沙漠[47];大約1.3萬年前,由于流入大西洋淡水的減少,導致溫鹽環流的驟停,使得格陵蘭島的氣溫驟降超過10℃,將該地區推回到幾個世紀以來的冰河時代[51];以及歷史上五次著名的物種大滅絕。Dakos等從地質記錄中重建了8個古代氣候突變的時間序列,采用了簡單的氣候模型來模擬地球從溫室到冰球的轉變;溫鹽環流模型,模擬冰河時代;北非沙漠化氣候模型,模擬了撒哈拉沙漠的誕生。通過模擬的數據分析了它們的自相關變化,表明在氣候變化之前就已經有了波動開始減緩的特征,用數學方法證明了自相關增加是臨界點的標志[17]。全球實地調研和遙感觀測相結合做的很好的研究有Berdugo等調查了20個生態系統結構和功能屬性對全球干旱的響應,表明干旱化導致多種生態系統屬性發生系統性的突變。這些變化依次發生在干旱值為0.54、0.7和0.8時,表現為植物生產力、土壤肥力、植被蓋度和豐富度的急劇下降[44]。
總而言之,定量預測僅限于較小尺度的系統,這種系統可以觀察到多穩態并從實驗上探索臨界點。比如我們可以知道,在一個淡水湖中當磷負荷在什么時候可能會變成渾濁的狀態。在較大尺度的預測則比較困難,比如我們很難知道亞馬遜熱帶雨林被燒掉多少之后會不可逆轉的轉化為稀樹草原狀態[2],所以在較大尺度上多采用模型模擬的方法來找到臨界點。而且生態系統每增加一個單位面積,該系統崩潰所需的時間單位就會越來越小,這意味著大系統往往比小系統移動得更慢,但速度卻不成比例[52]。
4 問題與展望
4.1 主要問題
4.1.1多穩態的機理研究不足
目前仍無法探測一個系統究竟存在哪些穩態,在什么條件下會轉變成另一種穩態。現有的多穩態模型是否可以起到通過樹木看到森林的作用,還需要通過模型和實驗相結合的方法來進行驗證。多穩態的突變推動機制一般為正反饋機制,是否還存在其他的推動機制值得深入探討。性狀動態的變化表明穩定性的喪失,還是對新的當地條件的適應也是值得思考的,因為性狀分布的變化可以表明群體處于壓力之下[53]。表型轉變之后可能會出現突然的種群或群落范圍的崩潰[54],或可以使種群應對不斷變化的環境條件,而不會出現豐度的顯著下降。雖然群落水平的穩定性的喪失仍可能通過改變個體大小的種間相互作用的強度來傳播。但是在某些條件下,例如當現有的遺傳變異耗盡時,性狀反應可能不會領先于即將到來的群落崩潰[11]。因此,量化基于特征的早期預警信號,從而減少預測的不確定性,是值得深入研究的。
4.1.2數據收集和處理問題
在實踐中,要想檢測到時間序列的預警信號,實時檢測可能來得太遲而無法采取行動,因為高質量和高分辨率的時間序列的數據很難快速獲得[22,55]。如何克服數據質量差的影響也是一大難點,例如,抽樣中人為因素的作用。以及如何消除噪音,目前在多穩態模型計算中的一個重要假設是模型系統的隨機動力學是生態數據的完整表示。而真實的生態數據容易產生多種誤差源,包括外部誤差和觀測誤差,以及識別和模擬動態過程所涉及的不確定性[38]。一些觀察到的時間序列中的不對稱性變化不一定意味著接近臨界轉換,因為它可能不是由于內在動力學而是由于外部噪聲中的不對稱性。現在普遍的做法是使用高斯白噪聲或零模型來模擬自然的誤差,但是這些方法過于簡單并不能很好的消除相關趨勢。所以,用適合的噪聲來消除嘈雜的外部變量,將真實的系統變化與其他的噪聲源分離是需要深入研究的。
4.1.3多穩態的尺度推繹問題
在多穩態的研究中,不同尺度系統有著不同的研究方法。例如,定量觀測實驗適用于小尺度的研究,而在較大尺度的研究則一般采用簡化的模型來模擬研究。在空間上,選擇正確的尺度有可能極大地改變空間信號的可靠性[56],而在時間序列中,時間尺度對于系統進程速率有重要的影響[53],了解尺度對這些信號可靠性的影響,從而選擇正確的尺度來應用這些方法是至關重要的。一些預警信號往往涉及的特定機制而不能推廣到其他系統[57]。如何將小尺度的方法和結論推演到更大的尺度上,如何在較大尺度上設計實驗來校正模型的參數,以及不同尺度的預警信號是否可以通用,仍需進一步的研究。
4.1.4預警信號問題
在穩態轉換的預警中,經常會出現兩種錯誤。第一類錯誤是如果系統災難性轉換正在到來,但是預警信號卻沒有顯示,所以我們沒有做出預警。此時我們就要承擔穩態轉換之后的后果,例如湖泊變渾濁后的治理和草原變成荒漠的治理。這可能因為在系統突然轉變之前,沒有漸進地接近臨界點。例如,信號可能保持在與臨界點一定的距離,但被罕見的極端事件驅動到另一個穩定狀態[22]。如果外部擾動機制隨著時間的推移而改變,就會出現統計困難,這可能會扭曲或抵消預警信號。
第二類錯誤是如果系統的災難性轉換還沒有到來,但是預警信號卻顯示臨界點即將到來,于是我們做出了預警。此時我們就需要承擔過度反應所承擔的損失,例如過度抑制放牧來阻止荒漠化的過程,這雖然使得草原適當生長,但是減少了食草動物的數量,損害了放牧者的利益,使得整體的生態系統服務價值沒有達到最大。第二類錯誤可能是由于預警信號的統計困難造成的。例如,增加的自相關的檢測可能需要長時間序列[58]。如果假想的預警信號不是接近穩態轉換的結果,就會出現誤報。這可能是偶然發生的,也可能是由于系統內部的混亂趨勢或外部擾動造成的[22]。
在到達臨界點之前,擾動通常會觸發一個轉變。因此,盡管穩態轉換指標的趨勢可能起到警示作用,但轉化的實際時刻仍然很難預測。當在實際應用時,一個關鍵問題是信號是否能夠足夠早地被檢測到,以便采取行動為防止系統發生穩態轉換。當涉及到即將到來的穩態轉換中做出決策時,我們面臨著巨大的不確定性。這種不確定性源于多種因素,包括預測不準確、數據不足和隱藏的非線性,以及對風險的感知和容忍度的特殊性,所以如何提高預測的準確性,量化正確的預警信號的概率,并對錯誤預警信號的比例進行加權是避免犯這兩類錯誤的關鍵。
4.2 未來發展方向
4.2.1基于生態系統恢復實踐不斷完善多穩態理論
系統在發生轉化之后,其結構和功能與之前的系統往往有很大的差別,內部的機制也發生了很大的變化。如果要想恢復到原來的狀態,僅僅重建系統到崩潰之前是遠遠不夠的,就像放牧模型一樣,因為過度放牧使得草原生態系統轉化為荒漠生態系統,通過減少放牧的數量并不能使荒漠生態系統恢復到草原生態系統,而需要做更多的努力,例如改善土壤的有機質含量、人工灌溉等等,才有可能恢復到之前的狀態。加強生態系統修復實踐的結果和數據的收集,將其應用在多穩態理論研究系統的臨界點以及反饋控制機制的完善上,是重點的研究方向[12]。
基于生物量和特征的指標具有廣泛的普適性,因為它們幾乎沒有對系統的基本結構和功能做出假設,因此可以為退化系統的恢復提供一個關鍵工具[9]。此外,現有的生態系統復原理論只考慮了超過臨界點的外部條件變化引起的穩態轉換,因此缺乏進化觀。而外部條件的變化對生態影響很小,也不會將系統推到臨界點之外。因此,這種變化不會立即引起穩態轉換,而是會觸發一個進化過程,使表型性狀超過臨界點,從而導致(在經歷了相當長的延遲之后)由選擇引起的穩態轉換,所以將進化動力學納入生態系統多穩態檢測是未來研究的方向之一[59]。
4.2.2多尺度下的生態保護管理和政策實踐
將多穩態的理論運用在生態保護管理實踐中,是多穩態理論運用的重要部分。可以通過分析不同尺度臨界點的計算和研究方法,并根據現有數據了解這些方法的局限性和優勢,不斷完善多尺度分析方法,更好地服務于設計目標。這其中的一個關鍵部分是將這些預警信號與決策框架[60],資源有限保護的現實[61],利益相關者和管理者之間的沖突[62]和適應性管理戰略[63]適當地聯系起來[11]。建立和保持預期生態系統狀態的彈性可能是生態系統管理最實際和有效的途徑。鑒于生態系統和環境變化的復雜性,如何將早期預警信號整合到最優決策理論中,是生態保護研究的重要內容[63]。此外,如何更好的將多穩態的理論和生態保護管理和政策相結合,是未來研究的重點[64]。
4.2.3時間和空間預警信號的綜合運用
將來自統一數據的多個信號組合在一起,可能會提供更可靠的穩態轉換的信號,例如把空間和時間預警信號以及性狀動力學等相結合[19,65]。因為一般的預警信號,如時間序列增加的自相關性和方差,檢測這些領先指標的滯后時間通常很長,而空間模式可能提供更強大的領先指標,因為它們比時間序列中的單個數據點包含更多的信息[30],但是空間指標很難識別與穩態轉換相關聯的時間模式,而且對于具有自組織圖案形成的系統,存在特定的信號,所以將多個指標相結合可能成為更準確的穩態轉換的指標。然而,目前在這一領域已經使用了非常有限的相對簡單的方法,例如多個統計信號的歸一化和求和[19,54]。更復雜的統計工具,如時間序列的多變量分析或機器學習算法,提供了簡單模型選擇框架的擴展,以及各種信號之間的互相關系[66]。所以將多元的預警信號整合到一起從而提供更準確的預測,是未來研究的方向。
致謝:張雪蓮女士繪制本文圖形,特此致謝。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
鴨綠江(2021年35期)2021-04-19 12:24:18
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
家庭影院技術(2017年9期)2017-09-26 03:41:45
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
