荒漠結皮層蘚類植物死亡對表層土壤水分蒸發和入滲的影響
2021-10-09 01:18:42李繼文尹本豐索菲婭周曉兵李永剛張元明
生態學報 2021年16期
關鍵詞:植物
李繼文,尹本豐,索菲婭,周曉兵,陶 冶,張 靜,李永剛,張元明,*
1 新疆大學生命科學與技術學院, 烏魯木齊 830046 2 荒漠與綠洲生態國家重點實驗室, 中國科學院新疆生態與地理研究所, 烏魯木齊 830011
水是限制荒漠生態系統生產力和植被演替的關鍵因素,水分的入滲和蒸發作為荒漠地表水文循環的重要環節深刻影響著降水在土壤中的再分配過程[1]。由于水分的限制,荒漠生態系統很難形成大面積連續分布的維管植物,但以藍藻、地衣、苔蘚等隱花植物組成的生物土壤結皮卻能發育成荒漠地表的重要覆被類型,其蓋度在部分區域可達70%以上[2]。生物土壤結皮除顯著影響土壤養分、土壤結構外[3- 4],在地表水分的入滲、蒸發等水文過程中也扮演重要角色[5- 6],但對其在水分入滲和蒸發方面的研究結論還存在較大爭議,主要觀點為結皮對水分的入滲和蒸發過程具有促進作用[7- 10]、抑制作用[10-12]及無作用[13-16]3種觀點。
蘚類結皮作為生物土壤結皮的高級階段,是生物土壤結皮的重要組成部分和生物量的最主要貢獻者。與維管束植物不同,蘚類植物多不具有維管束,屬于典型的變水植物,能夠隨環境水分的變化而變化[17],在截留降水、涵養水源等方面扮演重要角色[18-21]。尤其對于荒漠蘚類植物而言,為應對干旱少雨的惡劣環境,荒漠蘚類植物在個體水平和群體水平上均表現出較強的環境適應性[22-26]。研究發現,在干燥環境中齒肋赤蘚(Syntrichiacaninervis)葉片緊貼于莖,通過減少暴露于空氣的表面積來減少水分蒸發,而其毛尖結構能夠收集空氣中的水分,較無毛尖齒肋赤蘚毛尖能夠多收集10.26%的凝結水量[27]。在群體水平上,荒漠蘚類植物密集叢生并呈現墊狀分布,既提高了土壤毛細管系統的持水力,也在其表面形成了一個靜止層,減弱了水分蒸發速率[28]。
荒漠蘚類植物葉片多為單層細胞,對環境變化十分敏感,被譽為環境變化的指示劑。近幾十年來諸如增溫、氮沉降及極端天氣等事件的發生致使荒漠蘚類植物生存受到嚴重威脅。研究發現,荒漠蘚類植物對環境變化的敏感性要遠高于相同生境下的維管束植物,3 g N m-2a-1以上的氮沉降及夏季頻繁的小降水事件均會導致荒漠蘚類植物的死亡[29-31]。然而,荒漠蘚類植物的死亡這一過程是否會影響表層土壤的水分蒸發和入滲目前尚不清楚,因此,我們提出科學問題:荒漠結皮層蘚類植物死亡是否對表層土壤水分蒸發和入滲存在影響,若存在是如何影響的?為解答此科學問題,我們選取古爾班通古特沙漠優勢蘚類植物齒肋赤蘚為研究對象,以裸沙為對照,通過對比活蘚類結皮與蘚類植物死亡結皮層對土壤水分入滲和蒸發的影響,以期為深入探討荒漠蘚類結皮對荒漠生態系統水文過程的影響提供理論依據。
1 材料與方法
1.1 研究區概況
古爾班通古特沙漠是我國最大的固定半固定沙漠,海拔300—600 m,面積為4.88 ×104km2。年平均氣溫7.19℃,極端最高氣溫41.5℃;多年平均降水量128.6 mm,年均蒸發量大于2000 mm。該沙漠冬季具有穩定的積雪,積雪厚度通常在20 cm左右,積雪覆蓋持續100—160 d。該沙漠地表廣泛發育著生物土壤結皮和草本植物層片,早春積雪融化為生物土壤結皮及草本植物的生長提供了充足的水分。同時,維管束植物與生物土壤結皮鑲嵌分布是該沙漠主要地表景觀之一,其中生物土壤結皮主要包括藻結皮、地衣結皮和蘚類結皮3類,藻結皮多分布于沙壟的中上部,地衣多分布于沙隴中下部及丘間低地,而蘚類結皮主要分布在丘間低地[32]。
1.2 樣品采集及處理
在丘間低地隨機選取發育良好且生長相對一致的以齒肋赤蘚為優勢種的蘚類結皮,用PVC管(內徑10 cm,高度20 cm)垂直插入沙土中,小心取得未擾動的蘚類結皮樣品10份,在裸沙處,以同樣方法取得5份樣品,封底后運回至實驗室備用。為避免采樣時對蘚類結皮結構的擾動,采樣前用噴壺將結皮表面噴濕。在實驗室,采用65℃濕熱處理4 h將蘚類結皮滅活,設置5個重復。室溫放置24 h后,復水30 min,利用Mini-PAM 2500 (Walz, Hernz, Germany) 測定其熒光活性,確認其完全處于死亡狀態;存活結皮組與裸沙組施加等量的水分,各設置5個重復。將所有樣品放至溫度23℃、濕度16%的培養箱中自然蒸發15 d,以保證所有樣品中具有相同的初始含水量。
1.3 水分入滲與蒸發測定
1.3.1蘚類結皮水分入滲的測定
采用Mini-Disk Infiltrometer (Decagon公司,美國)便攜式滲透計測定入滲速率,將滲透計底部多孔燒結鋼盤平行放置于蘚類結皮表面,選擇適于蘚類結皮的4 cm (相當于0.4 kpa)壓頭,記錄儲水室初始水量后,間隔30 s記錄一次儲水室的水量,以時間的平方根為x,累積入滲深度為y,然后將得到的數據擬合到Zhang[33]提出的描述圓盤式滲透計滲透情況公式中,得到C1和C2常數值。
y=C1x1/2+C2x
利用得到的C2計算水分入滲速率:
K=C2/A
式中,A為給定土壤類型的van Genuchten參數與吸力率和入滲儀圓盤半徑之間的關系值。
累積入滲量=Vt-Vf(i)
式中,Vt為儲水室初始水量,Vf(i)為第i次測量時儲水室的水量。
1.3.2蘚類結皮水分蒸發測定
研究發現齒肋赤蘚在春季固定了其全年固碳量的50%以上[34],因此,本研究選擇了凍融季節的20 mm降水量來模擬測定齒肋赤蘚死亡對水分蒸發的影響[35]。將20 mm降水用小型噴壺均勻噴灑于蒸發儀[16]PVC管內的齒肋赤蘚表面,稱量其初始重量。將所有樣品放置于室溫為23℃的溫室內培養,在192 h內每間隔8 h測定一次樣品重量,192 h后間隔12 h測定一次,直至恒重,結合蒸發時間和蒸發量計算蒸發速率。
蒸發速率=(Wf(i)-Wf(i-1))/t
累積蒸發量=Wt-Wf(i)
式中,Wt為蒸發儀加水后的初始重量,Wf(i)為第(i)次測量時蒸發儀重量,Wf(i-1)為第(i-1)次測量時蒸發儀重量,t為第(i)次測量與第(i-1)次測量的時間間隔。
1.4 數據處理
用Excel對所得的數據進行常規處理。使用 SPSS 25.0軟件分別對活蘚類結皮與死亡蘚類結皮及裸沙的水分蒸發、入滲數據進行正態性檢驗和方差齊性檢驗,隨后對這3種地表類型間的水分蒸發、入滲數據進行單因素方差分析(one-way ANOVA)和多重比較(LSD)。采用Origin 9.0軟件進行繪圖。
2 實驗結果
2.1 蘚類結皮存活與否對土壤水分入滲速率的影響
3種不同地表類型在初始階段入滲速率差異較大,裸沙入滲速率表現為由最大速率逐漸降低,活蘚類結皮入滲速率表現為先增大后減小,而蘚類植物死亡的結皮層入滲速率則表現為逐漸增大,活蘚類結皮較蘚類植物死亡的結皮層及裸沙更快進入穩定入滲階段(圖1;表1)。在初始入滲階段和穩定入滲階段,與裸沙相比活蘚類結皮水分入滲速率均降低了近85%。同時,蘚類植物死亡的結皮層進一步限制了水分入滲,初始階段入滲速率較活蘚類結皮降低了60.11%,即使在穩定入滲階段,蘚類植物死亡的結皮層水分入滲率也只達到活蘚類結皮的85.91%。
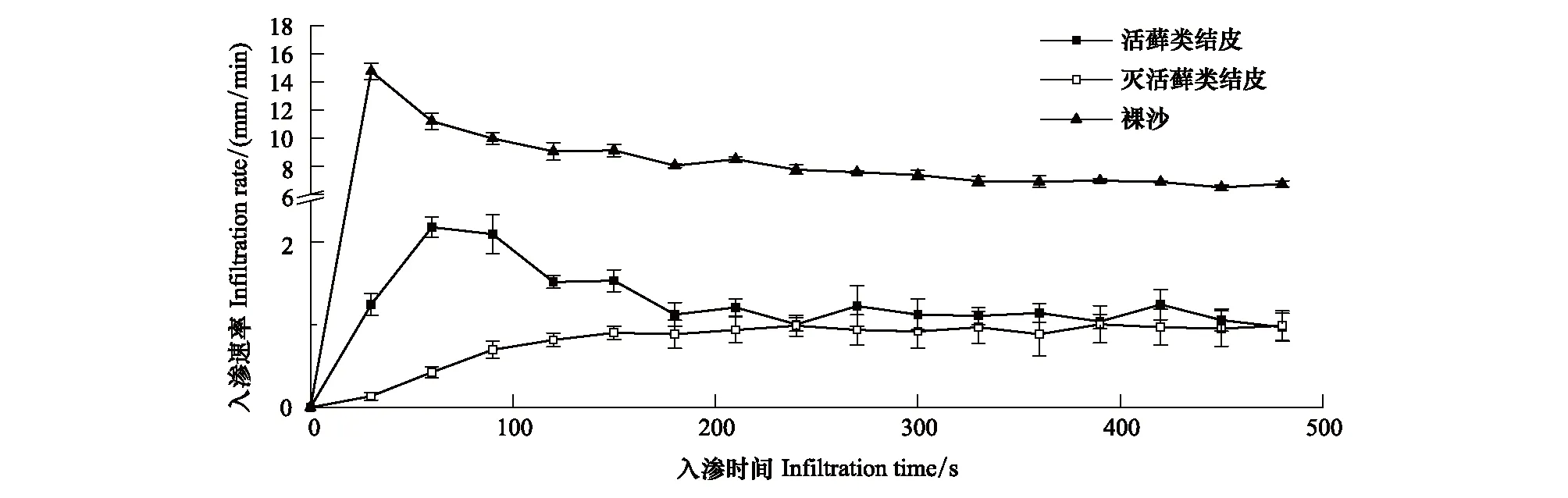
圖1 3種地表類型的水分入滲速率Fig.1 Infiltration rate of three surface types

表1 3種地表類型入滲特征參數
2.2 蘚類結皮存活與否對土壤水分累積入滲量的影響
與入滲速率結果一致,活蘚類結皮的累積入滲量也顯著低于裸沙,結皮層蘚類植物死亡后其累計入滲量顯著低于活蘚類結皮,且其抑制作用最大,累積入滲量分別是活蘚類結皮的64.48%,僅為裸沙的20.42%(圖2,P<0.05)。
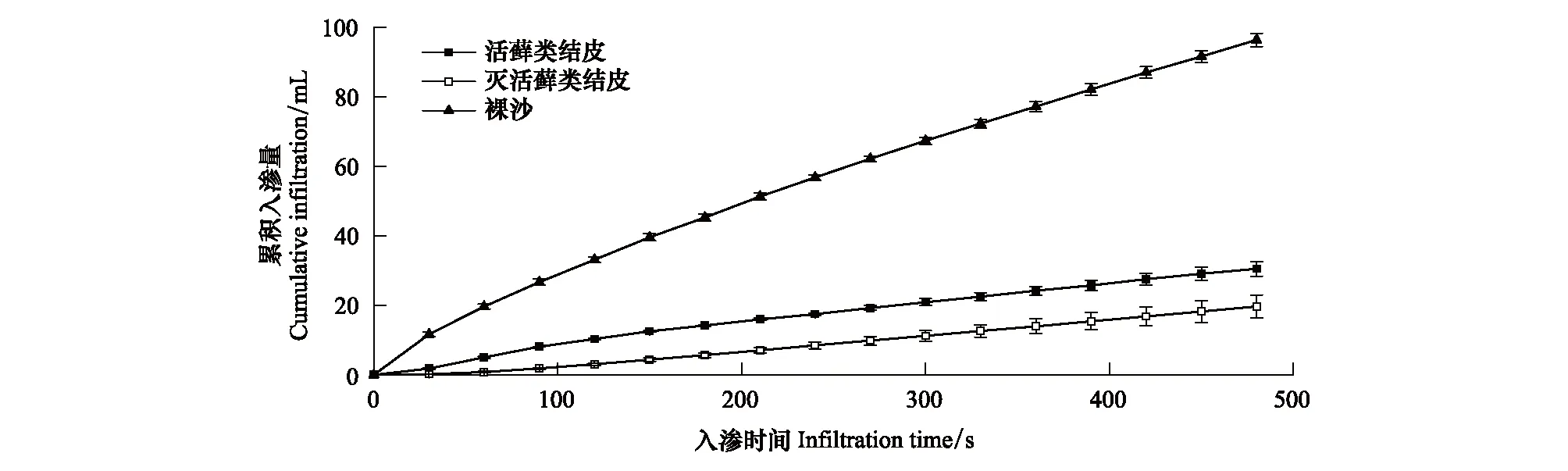
圖2 3種地表類型的水分累積入滲量Fig.2 Cumulative water infiltration of three surface types
2.3 蘚類結皮存活與否對土壤水分蒸發速率的影響
由圖3可知,裸沙、活蘚類結皮及蘚類植物死亡的結皮層蒸發過程均經歷了常速率、減速率、殘余速率3個階段,但3種不同地表覆蓋類型在各蒸發階段時長具有顯著性差異。常速率階段蘚類植物死亡的結皮層蒸發時間(70 h)顯著大于活蘚類結皮(34 h)及裸沙(40.4 h),減速率階段表現為活蘚類結皮蒸發時間(141 h)顯著大于蘚類植物死亡的結皮層(96.8 h)及裸沙(84.8 h),殘余速率階段蘚類植物死亡的結皮層(189.2 h)及裸沙(201.2 h)蒸發時間顯著高于活蘚類結皮(145 h)。在整個蒸發過程中,活蘚類結皮在常速率階段較裸沙平均蒸發速率降低31.03%,而殘余速率階段蒸發速率則加快30.59% (圖4)。

圖3 3種地表類型的水分蒸發速率Fig.3 Water evaporation rate of three surface types
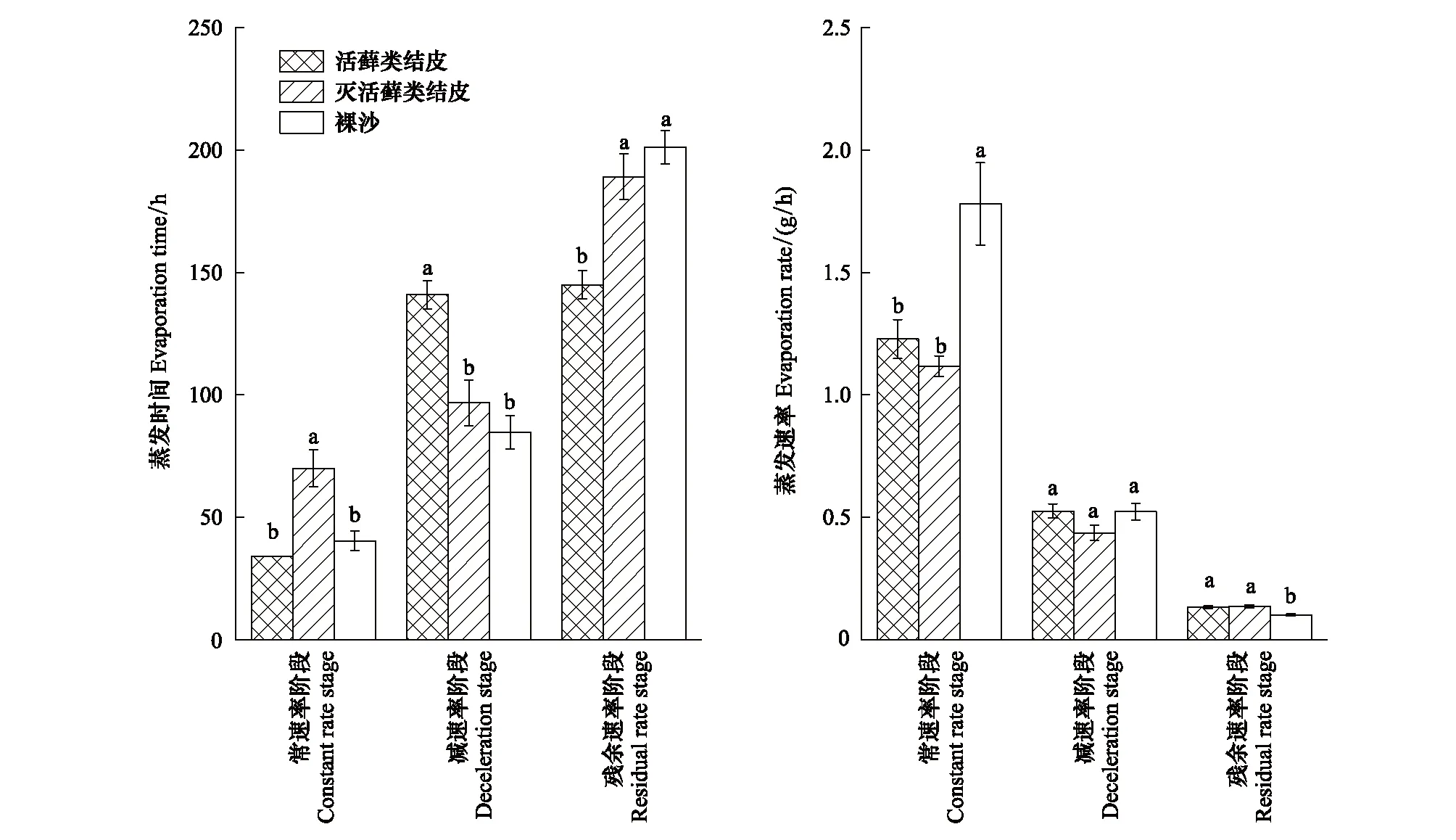
圖4 3種地表類型蒸發過程中蒸發時間和速率的差異性Fig.4 Differences in duration and rate of evaporation among three surface types during evaporation processes不同字母表示同一速率下不同地表類型之間在0.05概率水平上存在顯著差異(P<0.05)
2.4 蘚類結皮存活與否對土壤水分累積蒸發量的影響
在累計蒸發量方面,蘚類植物死亡的結皮層與活蘚類結皮沒有顯著性差異 (圖5)。與裸沙相比,蘚類結皮在前期減少了土壤水分累積蒸發量,而在蒸發后期則呈現相反趨勢,表現為蘚類結皮顯著大于裸沙,尤其是蘚類植物死亡的結皮層累計蒸發量最大。

圖5 3種地表類型的水分累積蒸發量Fig.5 Cumulative water evaporation of three surface types
3 討論與結論
生物土壤結皮是荒漠地表的重要覆被類型,能夠顯著影響地表土壤水文過程[36-37]。以往在生物土壤結皮對土壤水文過程影響的研究中,尤其是在水分入滲及蒸發方面一直存在爭議,主要觀點為結皮對水分的入滲和蒸發過程具有促進作用[7- 10]、抑制作用[10-12]及無作用[13-16]3種觀點。這可能由于研究地區、測定指標及所選生物土壤結皮類型不同導致。針對蘚類結皮而言,主流觀點認為蘚類結皮的存在可造成土壤水分淺層化,且隨著結皮蓋度增加水分入滲深度越淺[12, 38- 39]。這在本研究中也得到了證實,即蘚類結皮的存在顯著阻礙了水分的入滲,其原因可能與蘚類結皮層細沙含量較高有關[12, 40],蘚類植株及其與微生物分泌的多糖物質通過吸水膨脹,堵塞土壤孔隙且增強覆蓋土壤表面的能力[8, 41],大大提高了水分的滯留時間,這可以降低入滲速率,從而顯著影響土壤的含水量。
在荒漠生態系統中,蒸散是水平衡的關鍵過程[42-43],超過90%的年降水量會因這一過程而損失[44]。研究發現,蒸發過程主要受地表覆蓋類型的影響,例如植被類型和生物土壤結皮等[45]。本研究結果顯示,在20 mm降雨量條件下,蘚類結皮覆蓋對水分蒸發的影響隨不同蒸發階段而異,這與前期諸多學者研究結果相似,認為生物土壤結皮對水分蒸發不是簡單促進或抑制[16, 46-47]。在蒸發前期,齒肋赤蘚表面蠟質結構、葉片毛尖及毛尖間形成的交叉網絡降低了水分蒸發,且群體水平上墊狀的分布使其表面形成了一個空氣靜止層,減弱了水分蒸發。而在蒸發后期,蘚類結皮的存在卻提高了蒸發速率,這可能由于齒肋赤蘚已進入休眠狀態,降低保水能力,且蘚類植物的覆蓋形成了毛細管作用,從而促進了土壤水分的蒸發,而裸沙表面形成的硬殼狀土壤干燥層可能是造成蒸發后期裸沙蒸發速率較低的主要原因。值得注意的是,在蒸發測量結束后,蘚類結皮土壤水分累積蒸發量顯著大于裸沙,結合蘚類結皮對水分入滲的抑制作用,可以得知蘚類結皮的存在顯著了降低土壤含水量。
長期增溫和夏季頻繁小降水均會造成蘚類植物的死亡,進而影響荒漠地表的水分和養分。研究發現,不同時間序列的長期增溫(6年和10年)均會顯著降低蘚類結皮對水分的截留率[5]。但在本研究中發現,活蘚類結皮相比蘚類植物死亡的結皮層顯著降低水分的入滲速率和累計入滲量。這可能與蘚類植物死亡時間長短造成的蘚類結皮蓋度差異有關,長期增溫實驗導致苔蘚蓋度明顯降低,而本研究中采用的是即時死亡蘚類植物,不存在蓋度變化。但結皮層蘚類植物死亡造成的水分入滲降低是否與蘚類死亡后其表面疏水/吸水性物質變化、抑或蘚類毛尖、莖葉微結構變化有關還需要進一步深入研究。
生物土壤結皮作為干旱半干旱區重要的地表覆被類型,其重要的作用之一是對荒漠土壤水文過程的影響,我們的研究結果也強化了這種觀點。值得注意的是,盡管蘚類植物的死亡顯著降低了水分的入滲,但與活蘚類植物相比其累計蒸發量無顯著差異,甚至在蒸發后期略高于活蘚類結皮。前期有研究發現,干旱、半干旱區生物土壤結皮會通過影響生態系統的水文過程而造成維管植物的退化[48],尤其是以苔蘚為主的生物土壤結皮能顯著降低土壤濕度而導致人工種植灌木的死亡[49]。綜合考慮蘚類植物死亡對水分入滲的抑制,而對蒸發的無影響、甚至略有促進的研究結果,可以推測齒肋赤蘚的死亡在短期內會顯著降低土壤含水量,進而影響與其共存維管植物的生存。全球氣候的持續變暖已成為不爭的事實,降水及其時空模式也隨之出現不穩定的變化,研究預測顯示在本世紀末土壤含水量將減少5%—15%,全球干旱區的干旱化程度不斷加劇[50- 52]。以荒漠蘚類植物為代表的生物土壤結皮對氣候變化的響應要遠大于維管束植物,在全球氣候變化和土地利用方式的變化下,至2070年覆蓋地球陸地面積約12%的生物土壤結皮將減少25—40%,顯著影響荒漠地表的水文過程和養分循環[53]。因此,探究荒漠蘚類植物死亡后的生態效應對預測荒漠植物變化趨勢和保護荒漠植物物種多樣性和地表穩定性具有重要生態價值。
致謝:陸永興同學幫助寫作,劉鑫、梅月磊和張慶同學幫助實驗及數據處理,特此致謝。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
