基于三江源高寒草甸群落結構變化評估圍欄封育對草地恢復的影響
2021-10-12 01:12:42李寶林袁燁城祁佳麗
生態學報 2021年18期
關鍵詞:雜草
劉 巖,李寶林,*,袁燁城,祁佳麗,李 影,李 睿
1 中國科學院地理科學與資源研究所 資源與環境信息系統國家重點實驗室,北京 100101 2 中國科學院大學,北京 100049 3 國家環境保護青藏高原生態環境監測與評估重點實驗室,西寧 810007
三江源是中國江河中下游地區以及東南亞國家生態安全和區域可持續發展的重要生態屏障[1]。作為國家重點生態功能區的重要組成部分,過去幾十年間三江源在全球氣候變化與人類活動的雙重影響下,生態系統退化明顯[2-3]。鑒于此2005年和2014年國務院分別規劃投資75億元和161億元啟動了三江源生態環境保護與建設工程一期和二期工程,實施退牧還草、黑土灘治理、濕地保護等生態保護與修復項目[4-5]。因此,客觀評價區域生態質量狀況及其變化,對國家生態保護成效評估及生態保護政策的制定具有重要意義。
遙感數據覆蓋范圍廣泛且獲取方便,是進行大區域生態評估常用的技術手段。目前在三江源地區進行的生態評估主要是利用遙感手段和地理信息系統技術,側重從生態系統類型、生態系統生產力和生態系統服務角度進行評估。在國家實施三江源保護工程后,三江源地區生態系統格局穩定少動,水體與濕地生態系統整體有所恢復[6-9];植被指數、凈初級生產力、地上生物量等表征草地生產力指標的監測也表明植被呈現恢復趨勢,生態狀況逐漸改善[10-19],但也有一部分研究結果表明高寒草地生產力下降或基本保持穩定[20-23];另外,生態系統服務功能也呈好轉趨勢[24-26]。
遙感技術雖然對三江源區域宏觀生態狀況評估具有優勢,但無法反映生態系統結構的變化。生態系統整體生產力提高,不一定代表著草地群落結構向好的方向轉變。目前對三江源草地群落結構變化評估主要是基于地面樣方調查數據來進行,多數研究結果表明圍欄封育提高了地上生物量,草地禾本科和莎草科等優良牧草增加,生態狀況改善[10,16-17,27-30]。但部分學者也發現部分地區在圍欄封育后,雖然地上草地蓋度與地上生物量恢復,但沒有改變雜草的主導地位,莎草科等優良牧草覆蓋度減少[31-33],雜草生物量比禾本科增加幅度更大,甚至出現了禾本科和莎草科植物幾乎消失的情況[34]。圍欄封育導致毒雜草比例增加在其他區域的研究中亦有過很多報導,Li等在內蒙古高原沙漠化草地禁牧26年后發現一年生雜草顯著增加并在群落中占據了主導地位[35],1998—2016年在內蒙古進行的禁牧實驗發現在長達18年禁牧后雜草生物量占比顯著增加,而禾本科的百分比顯著減少[36],在美國[37-38]、土耳其[39]、捷克[40]、蘇格蘭[41]、伊朗[42-43]等地區的圍欄實驗也觀測到了禁牧后雜草生物量占比顯著提高的現象。從現有的研究成果來看,圍欄封育并不一定會改善草地植物群落結構。在三江源地區,由于地面監測數據獲取困難,現有研究多樣點數量有限且缺乏連續長時間的觀測,給三江源區圍欄封育效果評價帶來了很大的不確定性。本文擬基于多監測站點長時間序列草地各經濟類群地上生物量的變化來評價三江源草地群落結構的變化,從而評估三江源地區生態保護與修復措施的成效,為生態保護與修復績效評估以及生態保護政策制定提供可靠的依據。
1 研究方法
1.1 研究區概況
三江源是指長江、黃河、瀾滄江的源頭地區,位于中國青海省南部,地理位置介于北緯31°39′—36°16′、東經89°24′—102°23′之間,總面積約36.3萬km2,行政區域包括玉樹、果洛、海南、黃南四個藏族自治州的16個縣和格爾木市的唐古拉山鄉(圖1)。研究區以山原和峽谷地貌為主,海拔4000—6000m,西北-東南走向的巴顏喀拉山和阿尼瑪卿山將三江源分為以果洛州為主體的東北部黃河源區和以玉樹州為主體的西南長江、瀾滄江源區,地勢由東南向西北逐漸升高。三江源屬典型的高原大陸性氣候,冷暖交替、干濕分明、水熱同期,年平均氣溫為-5.6—7.8℃,年總降水量在262—772mm之間。植被類型以草地為主(面積占比約為68%),包括高寒草甸、高寒草原、沼澤濕地和溫性草原等[13],另外還分布有針葉林、灌叢、高山植被等。土壤類型按海拔從高到低依次分布有高山寒漠土、高山草甸土、高山草原土、山地草甸土、灰褐土、栗鈣土和山地森林土,沼澤化草甸土也較為發育,凍土分布廣泛。雖然在三江源生態保護工程開始實施后開始減畜,但三江源地區草地多年平均載畜總量高達2003.7萬羊單位,仍處于超載狀態[44]。

圖1 研究區地理位置、植被類型及采樣點分布圖Fig.1 The location,grassland type and sampling points of the Three River Headwater Region
1.2 數據來源及預處理
研究中使用的數據包括牧草營養枝高度、生殖枝高度、植被蓋度以及地上生物量四個指標,數據來源于青海省草原總站,數據獲取的時段為2005—2017年。數據采集時間一般在夏季天然草地牧草生長盛期,多為8月中上旬。樣地面積不小于1km2,每個樣地布設3—6個樣方,樣方之間的間距為250m左右,樣方面積為1m×1m。地上生物量測定時在樣方內將牧草齊地面刈割,按禾本科、莎草科、豆科、可食雜草、不可食雜草和毒草等經濟類群分別稱重,其中禾本科和莎草科牧草統計的是牲畜喜食優良牧草,兩個科中的毒雜草統計到其他相應的各毒雜草經濟類群中。在各經濟類群生物量基礎上,統計了總生物量和可食草總量以反映草地總生產力與經濟效用,統計了禾本科和莎草科牧草總生物量以及毒雜草總量來反映植被退化狀態。另外,為反映圍欄封育對草地植物群落結構的影響,還計算了各經濟類群、可食草總量、毒雜草總量以及禾莎總量占總生物量的比例。
2005—2017年三江源區共布設了227個樣地,由于布設的樣地是逐年增加的,樣地數量在不同年份差異較大。為保證數據的可比性,本研究只選擇了研究時段內序列完整的監測數據。其中,圍欄封育樣地從三江源保護工程開始實施的2005年開始圍封,且在研究時段內持續禁牧,放牧樣地從2005年開始持續放牧。在圍欄和放牧樣地中分別有39個和18個滿足要求,其中包括八個對照樣地,樣點分布如圖1所示。在數據分析時,對比圍欄內外各個指標統計量大小時采用了8對對照樣地數據。為盡量反映三江源圍欄封育整體效果,在對比圍欄內外變化趨勢時,圍欄內外都采用了全部樣地數據進行分析。
1.3 數據分析
采用Shapiro-Wilk檢驗和Kolmogorov-Smirnov檢驗,對牧草營養枝高度、生殖枝高度、蓋度、各經濟類群生物量、可食草總量、毒雜草總量、禾莎總量、總生物量以及各經濟類群、可食草總量、毒雜草總量以及禾莎草總量占總生物量比例等指標進行統計檢驗后,發現多不服從正態或對數正態分布。因此對各指標在圍欄區與放牧區總體大小差異進行統計檢驗時采用了Mann-Whitney U檢驗方法,總體大小比較時統計量采用中位數。
除了判斷圍欄區與放牧區各指標總體大小差異以外,本文還采用最小二乘法擬合各指標中位值的時間序列變化趨勢,采用F檢驗對擬合方程進行可靠性檢驗,并在此基礎上比較圍欄區與放牧區各指標總體大小變化趨勢的差別,具體計算公式如下:
(1)
ACRi=TCRi/(tn-t1)
(2)
y=a+β×t
(3)
式中,TCRi表示研究時段內第i個指標總變化率,ACRi表示研究時段內第i個指標年變化率,t1和tn分別為研究時段的起始和終止年份。y為某指標值,t為年份,a和β為擬合方程的參數,采用最小二乘法擬合獲得。
2 研究結果
2.1 主要指標總體大小
2.1.1高度、蓋度與總生物量
圍欄封育顯著提高了牧草的高度和地上生物量,蓋度略有提高但不顯著。2005—2017年從圍欄內外對照數據來看,圍欄內營養枝平均高度、生殖植平均高度和地上總生物量都顯著高于圍欄外對照區。圍欄內營養枝平均高度中位值為6.7cm,圍欄外為6.0cm,圍欄內比圍欄外高11.6%(P=0.043),圍欄內生殖枝平均高度中位值為17.5cm,圍欄外為15.1cm,圍欄內比圍欄外高15.9%(P=0.008),圍欄內地上生物量中位值為4569.8kg/hm2,圍欄外為3741.1kg/hm2,圍欄內比圍欄外高22.2%(P<0.001)(表1)。

表1 圍欄與放牧區主要指標總體大小差異Table 1 Significance test between two averages of main indicators between fenced and grazed samples
2.1.2各經濟類群生物量
2005—2017年只有禾本科、可食草總量和禾莎總量在圍欄封育區和放牧區有顯著的差異(P<0.05),而莎草科、豆科、可食雜草、不可食雜草以及毒草則沒有顯著差異。圍欄封育區禾本科生物量中位值為1187.7 kg/hm2,放牧區為792.2kg/hm2,圍欄區比放牧區高49.9%。圍欄封育區可食草生物量中位值為3554.5 kg/hm2,放牧區為2861.7kg/hm2,圍欄區比放牧區高24.2%。禾莎總量在圍欄封育區中位值為2207.3 kg/hm2,在放牧區為1654.5 kg/hm2,圍欄區比放牧區高33.4%(表2)。
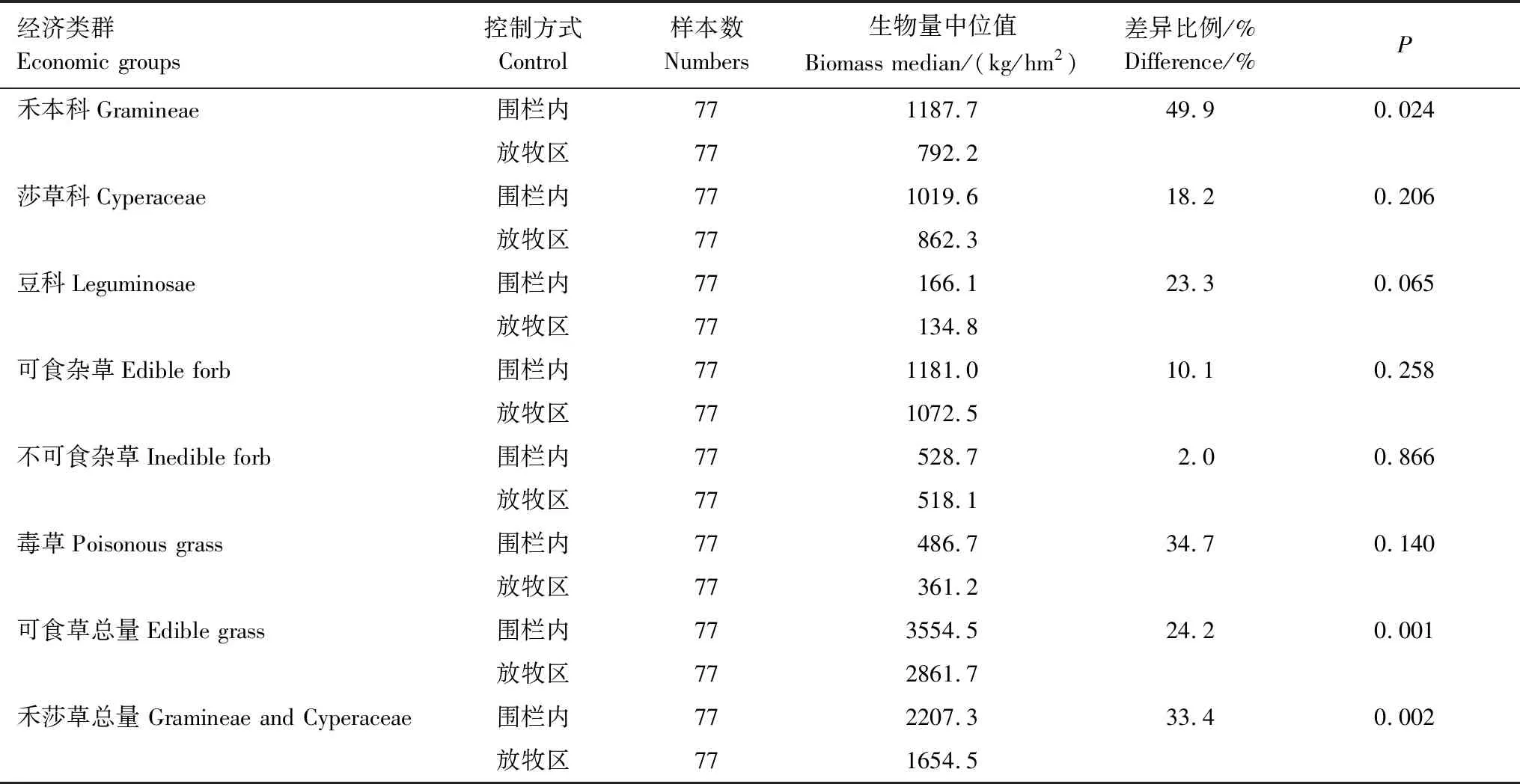
表2 圍欄與放牧區各功能群生物量總體大小檢驗Table 2 Inspection of the difference in biomass of functional groups in fenced and grazed sites
2.1.3各經濟類群生物量占比
2005—2017年6個經濟類群中莎草科生物量占比最高,在圍欄封育區和放牧區分別為26.7%和28.6%;其次為可食雜草,占比分別為25.4%和24.5%;禾本科占比分別為22.2%和20.1%;不可食雜草為12.3%和13.0%;毒草占比分別為9.8%和10.1%;豆科牧草占比最小,都為3.6%。各經濟類群生物量占比在圍欄封育區和放牧區呈現出相似的規律,各個指標在圍欄內外都沒有顯著的差異(表3)。

表3 圍欄與放牧區各功能群生物量比例總體大小Table 3 The difference in biomass proportion of functional groups in fenced and grazed sites
2.2 主要指標變化趨勢
2.2.1高度、蓋度與總生物量
2005—2017年總生物量在圍欄封育區和放牧區都沒有顯著的趨勢性變化(P>0.1),營養枝平均高度、生殖枝平均高度與植被蓋度在圍欄封育區都呈現出顯著的減少趨勢(P<0.05),而在放牧區變化趨勢都不顯著(P>0.1)。在圍欄封育區,研究時段內圍欄封育區營養枝平均高度減少了34.8%,平均每年減少2.9%;生殖枝平均高度減少了38.4%,平均每年減少3.2%;植被蓋度減少了4.8%,平均每年減少0.4%(圖2)。

圖2 圍欄封育區與放牧區主要指標變化趨勢 Fig.2 Change trend of main indicators in fenced and grazed sites表示所用擬合方程的顯著性水平;**表示顯著(P<0.05),***表示非常顯著(P<0.01)
2.2.2各經濟類群生物量
2005—2017年各經濟類群生物量多數未表現出明顯的趨勢性變化,只有毒草和可食雜草在圍欄封育區和放牧區表現出趨勢性變化(P<0.1)。在研究時段內圍欄封育區毒草生物量共增加了335.0%,平均每年增加27.9%;可食雜草增加了26.2%,平均每年增加2.2%。在研究時段內放牧區毒草增加了136.6%,平均每年增加11.4%;可食雜草減少了29.5%,平均每年減少2.5%(圖3)。

圖3 圍欄封育區與放牧區各經濟類群生物量變化趨勢Fig.3 Change trend of biomass of different economic groups in fenced and grazed sites表示所用擬合方程的顯著性水平,*表示邊際顯著(P<0.1),**表示顯著(P<0.05)
2.2.3各經濟類群生物量占比
2005—2017年各經濟類群生物量占比多數表現出明顯的趨勢性變化(P<0.1),但在圍欄封育區與放牧區具有明顯的差異。在5個基本經濟類群中,圍欄封育區禾本科、莎草科、可食雜草和毒草各功能群生物量占比有顯著的趨勢性變化;放牧區可食雜草、不可食雜草與毒草具有明顯變化趨勢(P<0.1),豆科牧草生物量占比在圍欄封育區和放牧區都沒有顯著的變化趨勢。在圍欄封育區和放牧區,可食草總量都具有顯著的變化趨勢,而禾莎草總量則只有圍欄封育區具有顯著的變化趨勢(P<0.1)。
在圍欄封育區禾本科和莎草科生物量占比都呈顯著減少趨勢,研究時段內分別減少了48.2%和23.9%,平均每年分別減少4.0%和3.3%;毒草占比增加了230.2%,平均每年增加19.2%;可食雜草增加了42.0%,平均每年增加3.5%;禾莎草總量占比減少了31.2%,平均每年減少2.6%。在放牧區,可食雜草占比在研究時段內減少了37.7%,平均每年減少3.1%;不可食雜草占比增加了113.9%,平均每年增加9.5%;毒草在研究時段內增加了170.3%,平均每年增加14.2%。圍欄封育區和放牧區的可食草總量占比在研究時段內分別減少了13.5%和14.9%,平均每年減少1.1%和1.2%(圖4)。
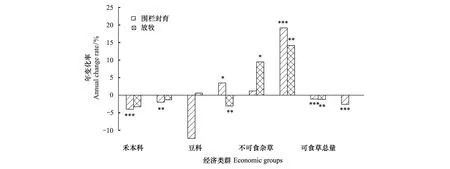
圖4 圍欄封育區與放牧區各經濟類群生物量占比變化趨勢Fig.4 Change trend of biomass ratios of different economic groups to total biomass in fenced and grazed sites表示所用擬合方程的顯著性水平,*表示邊際顯著(P<0.1),**表示顯著(P<0.05),***表示非常顯著(P<0.01)
3 討論
3.1 圍欄封育區群落構成變化形成機制
圍欄封育區草地地上生物量在時間序列上表現為波動中保持穩定(圖5),但其內部種群結構發生了變化。具體表現為減少未退化狀態下作為建群種的禾本科和莎草生物量占比,增加雜草與毒草占比。研究時段內禾本科和莎草科生物量占比分別減少48.2%和23.9%,毒草增加了230.2%,可食雜草增加了42.0%,而從整個可食草量占比來看,也降低了13.5%(圖3)。從放牧區來看,禾本科和莎草科生物量占比都較穩定,未呈現顯著性變化,生物量占比變化主要通過減少可食雜草增加不可食雜草和毒草來實現(圖4)。研究時段內,可食雜草減少了37.7%,不可食雜草和毒草分別增加了14.6%和170.3%。可見圍欄封育區盡管總生物量保持穩定,但群落結構已呈現出顯著的惡化趨勢,且比同期自由放牧區惡化更為突出。這與Li等[31]在三江源的研究結果類似。
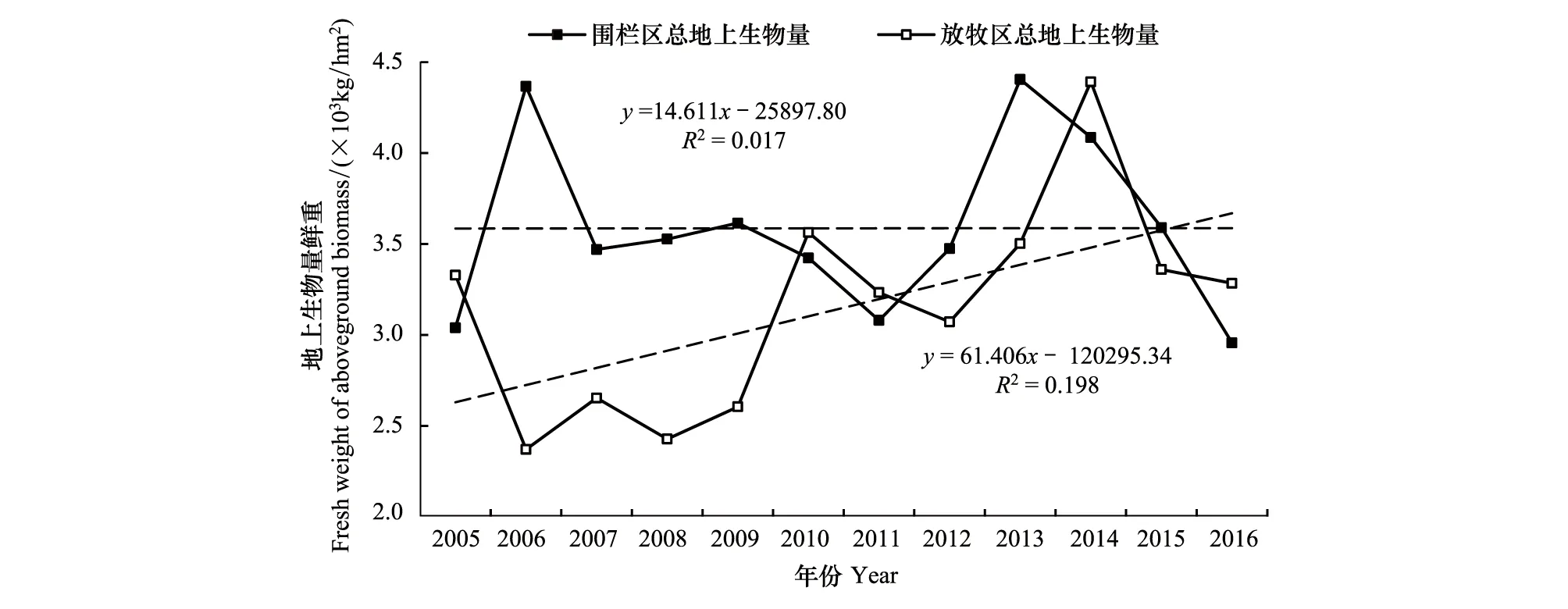
圖5 圍欄封育區與放牧區地上生物量年際變化Fig.5 Variation of aboveground biomass in fenced and grazed sites
圍欄區莎草科生物量占比下降主要是由于莎草科牧草在與禾本科和毒雜草對光照的競爭中處于劣勢。在放牧壓力消除后,由于禾本科牧草和毒雜草等多植株高大,枝葉繁盛,在光熱資源競爭中處于優勢地位[45-46],而植株低矮的莎草科牧草作為下繁草,由于植株被遮陰導致光合速率降低[47],在競爭中處于弱勢地位,盡管不再被牲畜啃食,生物量占比也會逐年下降。
毒雜草尤其是毒草快速增長,首先與放牧壓力消除后其對光熱資源的強競爭能力有關。青藏高原的毒雜草主要有黃花棘豆(Oxytropisochrocephala)、狼毒(Stellerachamaejasme)、醉馬草(Achnatheruminebrians)、黃帚橐吾(Ligulariavirgaurea)等[48]。毒雜草大多株體較高,具有繁茂的葉層以及較大的比葉面積,在與禾本科和莎草科尤其是莎草科植物對光熱資源的競爭中處于優勢地位,導致天然草地植物禾本科和莎草科牲畜喜食的優良牧草逐漸減少,草地經濟價值逐漸降低[49-50]。其次,由于放牧壓力的消除,毒雜草不再遭受牲畜的踩踏,植株相對較高大的莖不再折斷,葉層也就更為繁盛,使禾本科與莎草科這些耐牧牧草進一步喪失了競爭優勢。另外,毒雜草多以有性生殖為主,繁殖能力強,且生長迅速,適應性強,進一步加劇了毒雜草的蔓延,從而使毒雜草生物量占比逐漸增加[51-53]。
禾本科牧草在圍欄封育后占比降低,主要與其同毒雜草競爭中失去對光熱資源的競爭優勢有關,可能還與禾本科與莎草科分布區地表會產生更多枯落物有關。天然草地中禾本科與莎草科植株多呈現為緊伏地表的流線型墊狀體,有效地減少了植物體的受風面積,可以保留更多的枯落物,而毒雜草植株高大且比葉面積大,在枯黃后枯落物尤其是葉子會快速被秋冬季大風所侵蝕,地表很難保留較多的枯落物。從表4可以看出,禾本科生物量占比與返青期和主要生長季平均氣溫具有良好的相關關系,而毒雜草受溫度影響則小得多(表4)。在三江源低溫嚴寒環境下,較多的枯落物限制了禾本科(包括莎草科)牧草對光熱資源的獲取,進一步導致了禾本科牧草生物量占比逐漸減少。另外,禾本科植物多數植株較高,對富含木質素結構成分的氮投資更多,其枯落物質量較差,分解速率慢,導致相對較高的地表遮蔽和較低的土壤養分,不利于植物生長[53]。

表4 生長季降水與溫度與各功能群生物量比例相關系數Table 4 Relationship between temperature,precipitation and biomass proportion in growing season
另外,毒草對禾本科牧草存在化感作用,可通過自身的分泌物或揮發物抑制種子的萌發和胚根的生長來限制禾本科牧草的發育[54]。馬瑞君等對黃帚橐吾(Ligulariavirgaurea)的研究發現其體內富含的揮發物會減少禾本科植物種子的萌發率、抑制種子的萌發速率,從而獲得競爭優勢[55]。唐燕研究表明白苞筋骨草(Ajugalupulina)和鐵棒棰(Aconitumpendulum)的水浸提液對當地禾本科牧草具有顯著的化感抑制作用[56]。
3.2 放牧區群落結構變化與放牧壓力
國家在十二五(2011—2015)和十三五(2016—2020)開始實施“草原生態保護補助獎勵政策”,其中對禁牧區域以外的草原根據承載能力核定合理載畜量,實施草畜平衡管理,并按照每年每公頃0.17元的測算標準給予草畜平衡獎勵。從監測數據來看,盡管總生物量都沒有表現出顯著的趨勢性變化,但放牧區草地群落結構已發生了明顯變化,說明該政策對三江源高寒草甸區群落結構確實產生了深刻的影響。
從圍欄封育區來看,禾本科、莎草科生物量占比呈顯著減少趨勢,毒草和可食雜草呈增加趨勢;而在同時段放牧區的群落結構可以看出明顯的階段性特征,禾莎草生物量占比先減少后增加,2005—2011年快速減少,而2012—2017年又呈明顯增加趨勢(圖6)。可食雜草生物量占比在2005—2011年維持較高的比例,而2012—2017則維持較低的比例。毒草生物量占比2005—2011年持續增加,而2012—2017則持續降低。

圖6 各經濟類群生物量占比年際變化Fig.6 Interannual variation of biomass proportion of each functional group
各經濟類群生物量占比變化的時間轉折點與國家天然草原保護政策實施節點非常吻合。《青海省草原生態保護補助獎勵機制實施意見(試行)》青政辦[2011]229號明確要求,在2011—2015年對全省1.6千萬公頃中度以上退化天然草原實施禁牧,《新一輪草原生態保護補助獎勵政策實施方案(2016—2020年)》青政辦〔2016〕195號文要求在2016—2020年開始的新一輪草原生態保護補助獎勵政策中,繼續對這1.6千萬公頃天然草原實施禁牧。可見,實施草畜平衡明顯改善了草地群落的物種結構。在草畜平衡政策開始的第二年(2012年),草畜平衡在非禁牧區開始全面實施,在減少了牲畜壓力后,禾本科與莎草科等優良牧草得到一定程度的修養生息,生物量占比由未實施前的持續減少而改變為持續增加,相應地可食雜草則占比迅速減少,基本穩定在20—30%之間,不可食雜草生物量占比改變了持續增加的趨勢,從2014年開始生物量占比也基本穩定在10%以下。
需要注意的是,毒草生物量占比在實施草畜平衡政策后,迅速增加而后逐漸減少。這反映了在實施草畜平衡政策前,在牲畜壓力減輕后,這些具有較大葉面積的毒雜草受牲畜踩踏的機會減少,而退化草地又為其擴張提供了生長空間。然而,這些闊葉毒草畢竟耐牧性差,在后續牲畜不斷踩踏下植株受損,影響了毒草的生長與繁殖,其生物量占比逐漸減小,可見牲畜的踩踏對控制毒雜草擴張就有重要意義。
3.3 持續圍欄封育負面效應與對策
三江源地區目前采取圍欄封育的區域并不能顯著提高草地的生產力,反而造成了草地群落結構變差等負面效應。未退化狀態下作為建群種的禾本科與莎草科牲畜喜食的優良牧草生物量占比持續下降,毒雜草尤其是毒草增加迅速,經過10余年的封育草地群落結構變差且沒有出現好轉的跡象。草地群落結構變差首先影響的是草地經濟價值,禾莎草生物量占比在研究時段內降低了31.2%,即使可食雜草增加了42.0%,但可食草總量總體上降低了13.5%,而且減少的主要是優良牧草。
現有的圍欄封育措施還可能帶來生態風險。禾本科和莎草科植物一般具有較發達的須根,尤其是莎草科植物根冠比明顯高于雜類草[57-58]。禾本科和莎草科生物量占比下降,尤其是莎草科生物量占比下降,意味著高寒草甸區地下根系量減少[59],從而降低了草地生態系統的水源涵養和水土保持功能。另外,禾本科和莎草科凋落物分解速率明顯低于雜類草[60],雜類草占比增加也會導致地表凋落物存量減少,進一步削弱了草地水源涵養和水土保持功能。
毒雜草增加可能加速圍欄封育區的土壤風蝕。毒雜草植株多高大且比葉面積大,在枯黃期枯落物會快速被秋冬季大風所侵蝕,地表很難保留較多的枯落物,在植被枯萎季節大大加速了土壤風蝕。禾本科與莎草科牧草披針形和針形葉片及以緊伏地表的流線型墊狀體,可有效地減少植物體的受風面積[61],從而保留更多的枯落物,大大減少土壤風蝕影響。
圍欄封育后植被群落結構改變,可能進一步加劇鼠害,進而加劇草地的退化。在三江源地區主要野生鼠類為喜馬拉雅旱獺、高原鼢鼠和高原鼠兔,其食譜狹窄且對鵝絨委陵菜(Potentillaanserina)、甘肅棘豆(Oxytropiskansuensis)、藍花棘豆(Oxytropiscaerulea)、蘭石草(Lanceatibetica)等雜類草有偏好性[62-63]。毒雜草增加為這三類動物都提供了更為豐富的食物來源。而植被生殖枝和營養枝變得低矮,蓋度降低,也會為嚙齒類動物提供了更好的生存環境[64]。
草地禁牧是青海省草地恢復的主要手段之一。然而,從10余年多樣點持續地面觀測結果來看,在三江源地區中重度退化草地中長期持續禁牧并不能顯著提高草地的生產力,反而會導致禾本科與莎草科牲畜喜食的優良牧草生物量占比持續下降,毒雜草尤其是毒草顯著增加,降低草地的經濟價值,也會損害水源涵養與水土保持等生態功能。目前,放牧在草地生態恢復中的作用容易被忽視,牲畜踐踏與采食可以增加牧草分蘗,加速牧草葉片更新,進而可以提高光合能力。牧草種子在牲畜踐踏后更易進入土壤,且牲畜踐踏對闊葉雜草的控制,牲畜糞便對土壤氮、磷平衡的維持以及牲畜活動對鼠類的控制等,對草地恢復都具有正面意義。因此,草畜平衡政策可以在不同退化程度草地上實施,在草畜平衡的前提下對各類退化草地減畜可能是目前草地恢復需要特別重視的途徑。實際操作中可以基于牲畜可以采食的實際產草量以及生態保護目標,確定更為合理的載畜量,對各類退化草地實施更為精準的草畜平衡管理。
4 結論
圍欄封育是三江源生態保護與修復主要措施之一,本文基于2005—2017年多樣地草地各經濟類群地上生物量的監測數據,對草地群落結構變化進行了分析。研究結果表明研究時段內三江源地區圍欄封育顯著提高了群落地上生物量,在圍欄封育期間地上生物量穩定,但群落結構出現惡化趨勢。研究時段內禾本科和莎草科優良牧草生物量占比分別減少了48.2%和23.9%,毒草增加了230.2%,可食雜草增加了42.0%。而放牧區草地群落結構變化具有明顯的階段性特征,禾莎草生物量占比先減少后增加,毒草雜草生物量占比先增加后減少,其變化可能與國家推行的草原生態保護補助獎勵政策中草畜平衡措施有關。從現有研究結果來看,大部分研究中圍欄區與放牧區的毒雜草比例和生物量無顯著性差異,可能與圍欄封育的觀測年限較短有關。現有的持續圍欄封育措施不但降低了草地的經濟價值,也帶來了一定的生態風險。基于草地實際產草量以及生態保護要求,確定更為合理的載畜量,實施更為精準的草畜平衡管理,應該比切斷牲畜與草地關系的長期禁牧封育更為有效。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
今日農業(2021年5期)2021-11-27 17:22:19
當代作家(2021年12期)2021-01-15 21:39:15
高中生學習·閱讀與寫作(2019年2期)2019-09-10 07:22:44
現代園藝(2017年11期)2017-06-28 11:32:57
新農業(2016年19期)2016-08-16 12:00:18
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:11
現代農業(2015年5期)2015-02-28 18:40:49
雜草學報(2012年1期)2012-11-06 07:08:33
