短期增溫對青藏高原高寒草甸不同植物根際叢枝菌根真菌的影響
2021-10-19 13:50:54王譚國艷馬志遠(yuǎn)李沛洋郭俊宏張嘉懿蔣勝競
草地學(xué)報 2021年9期
王譚國艷, 馬志遠(yuǎn), 李沛洋, 郭俊宏, 張嘉懿, 蔣勝競*
(1. 蘭州大學(xué)草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室/蘭州大學(xué)農(nóng)業(yè)農(nóng)村部草牧業(yè)創(chuàng)新重點實驗室/蘭州大學(xué)草地農(nóng)業(yè)與科技學(xué)院, 甘肅 蘭州 730020; 2. 北京大學(xué)城市與環(huán)境學(xué)院, 北京 100871)
自工業(yè)革命以來,由于化石燃料的過多使用和人類活動的增加,全球氣溫正不斷升高[1]。以全球氣溫升高為主要特征的氣候變化問題正改變著陸地生態(tài)系統(tǒng)的結(jié)構(gòu)和功能,影響人類的生存和健康[2-3]。青藏高原作為地球上海拔最高和面積最大的高原對氣候變化尤為敏感[4]。近些年,許多研究證實了氣候變暖不僅會改變植物物候[5]及群落組成[6],對土壤微生物的多樣性及功能同樣也具有重要影響[7-8]。然而,關(guān)于土壤微生物的重要類群之一的叢枝菌根(Arbuscular mycorrhizae,AM)真菌如何響應(yīng)氣候變暖的研究相對較少。
叢枝菌根真菌隸屬于球囊菌門(Glomeromycota),在自然界中廣泛分布,可以與陸地生態(tài)系統(tǒng)中80%以上的高等植物建立共生體系,形成菌根結(jié)構(gòu)[9]。在菌根共生體中,AM真菌不僅能夠促進(jìn)植物對礦質(zhì)營養(yǎng)(磷素和氮素等)的吸收、增強(qiáng)植物抗逆抗病等生理作用[10],還會影響植物的群落組成[11-13]。此外,菌根真菌還能夠影響土壤細(xì)菌群落結(jié)構(gòu)[14],穩(wěn)定土壤結(jié)構(gòu)[15],促進(jìn)土壤養(yǎng)分循環(huán)[16]。AM真菌有重要的生理生態(tài)功能,但目前對AM真菌如何響應(yīng)氣候變暖還沒有一致的結(jié)論。
AM真菌的生長只能在一定的溫度范圍內(nèi)進(jìn)行并且具有最適的生長溫度,因此溫度上的任何變化都會直接影響AM真菌。例如,Heinemeyer等在2004年曾發(fā)現(xiàn)溫度升高可以促進(jìn)AM真菌根外菌絲的生長[17]。此外,因AM真菌對宿主植物或土壤環(huán)境具有一定的選擇性[18-20],所以氣候變暖也可以通過改變植物群落結(jié)構(gòu)或土壤性質(zhì)從而影響AM真菌[21-22]。近些年,學(xué)者在青藏高原針對AM真菌對氣候變暖的響應(yīng)做了大量研究[23-26]。如Jiang等[24]和Wei等[26]發(fā)現(xiàn),AM真菌的多樣性及群落結(jié)構(gòu)并不受溫度升高的顯著影響,而Shi等卻發(fā)現(xiàn)模擬增溫降低了AM真菌的多樣性,但對AM真菌的群落組成沒有顯著影響[25],這一結(jié)果與Yang等在2013年的研究結(jié)果相反,他們發(fā)現(xiàn)模擬增溫雖然對AM真菌的多樣性沒有顯著影響,卻顯著影響了AM真菌的群落組成[23]。作為專性共生菌,AM真菌的群落組成與植物群落特征密切相關(guān),不同的植物群落組成可能影響了AM真菌對模擬增溫的響應(yīng)。因此,在植物物種水平上探究AM真菌對模擬增溫的響應(yīng),繼而得出普遍性規(guī)律對AM真菌的研究至關(guān)重要。
近些年由于高通量測序技術(shù)的快速發(fā)展,對AM真菌的研究大多集中于土壤或根系的分子學(xué)研究,對以AM真菌孢子形態(tài)鑒定為主的報道相應(yīng)減少。然而,AM真菌孢子作為重要的繁殖體,無論在AM真菌種群生長或是群落構(gòu)建中都具有重要作用。此外,形態(tài)鑒定的方法較分子鑒定方法具有更好的直觀性,這一方法避免了因核酸提取及PCR反應(yīng)等實驗所帶來的誤差,因此對探究氣候變暖對AM真菌孢子的影響仍具有重要意義。本試驗借助野外模擬增溫平臺,以盆栽的單物種植物為研究對象,通過探究模擬增溫對不同植物根際土壤AM孢子組成的影響,旨在探討不同植物根系A(chǔ)M真菌孢子的分布模式及差異以及在植物物種水平上AM真菌如何響應(yīng)模擬增溫。
1 材料與方法
1.1 研究地點概況
本項目在中國科學(xué)院海北高寒草甸生態(tài)系統(tǒng)定位研究站(北緯37°37′,東經(jīng)101°19′,海拔3 200 m)進(jìn)行。該研究站地處青藏高原東北隅祁連山區(qū)門源縣境內(nèi)。該地氣候類型屬于高原大陸性氣候,具有典型的高原寒溫濕潤性氣候特征。海北站所在地區(qū)主要的土壤類型包括高山草甸土、高山灌叢草甸土和沼澤土主要為草氈寒凍雛形土(高山草甸土)[27],土壤中有機(jī)質(zhì)貯量豐富,氮、磷絕大部分以有機(jī)態(tài)形式存在,礦化作用較弱[28]。該地區(qū)為典型的以嵩草屬植物為建群種的高寒嵩草草甸[29],主要物種有矮嵩草(Kobresiahumilis),羊茅(Festucaovina),垂穗披堿草(Elymusnutans)等[30]。
1.2 試驗設(shè)計
本研究于2016年7月份原位選取高寒草甸常見的15種植物(表1)幼苗進(jìn)行室外移栽試驗,每盆4株幼苗,所選植株幼苗長勢相近且無明顯的病蟲害。花盆直徑為20 cm,埋入地下深度為60 cm,培養(yǎng)基質(zhì)采用原位土與沙子按2∶1比例混合。本試驗采取的增溫方式為紅外燈管(220 V,長1 m,寬2.2 cm)加熱,燈管距地面距離1.2 m,增溫幅度約為2℃;對照組僅設(shè)置同等規(guī)格的燈罩。每個處理設(shè)置6個重復(fù),共計180個樣品。

表1 本研究所選取的植物物種
1.3 樣品采集
經(jīng)過連續(xù)兩年的增溫試驗后,于2018年8月底進(jìn)行破壞性采樣,采集花盆內(nèi)的根際土壤樣品,自然風(fēng)干后進(jìn)行AM真菌孢子的分離、計數(shù)與鑒定。AM真菌孢子的分離采用濕篩傾析-蔗糖離心法[31]并稍作修改,具體操作如下:稱取25 g風(fēng)干土壤于1 L的鋼制燒杯中,加入1 000 mL的自來水,用玻璃棒快速攪拌,使得底部的土壤全部懸浮。靜置3分鐘后再次攪拌,如此反復(fù)3~4次,使得土壤的團(tuán)聚體結(jié)構(gòu)破碎,孢子全部釋放到水中。最后攪拌使土壤顆粒懸浮后靜置約7秒后將渾濁液迅速傾倒至兩層篩網(wǎng)(上層孔徑為200 μm,下層孔徑為38 μm)過濾,將下層網(wǎng)篩上的過濾物轉(zhuǎn)移到100 mL離心管中,3 000 r·min-1離心3 min后棄上清,加入50%(m/v)蔗糖溶液充分震蕩后3 000 r·min-1離心7 s,此時AM孢子懸浮于蔗糖溶液層,而絕大部分泥沙沉在底部。將上層蔗糖溶液倒入孔徑38 μm篩網(wǎng)的一側(cè),用自來水小心沖洗后將篩子上的孢子泥沙混合物快速的抽到濾紙上(直徑7 cm),轉(zhuǎn)移到培養(yǎng)皿中,體視鏡下用鑷子小心將孢子挑取到一張新的做好標(biāo)記的濾紙上。分離出的孢子樣品放在4℃冰箱中(保持濕潤)保存用于后續(xù)的試驗。
1.4 AM真菌孢子的形態(tài)鑒定
將分離好的孢子取出,在體視鏡下通過觀察其顏色、大小、形狀、有無連孢菌絲等特征將所有孢子做初步的聚類、拍照、計數(shù)。取載玻片,滴一滴Melzer-PVLG染液,用鑷子將初步聚類的各組的孢子全部夾到染液中,用蓋玻片蓋上,大拇指輕輕按壓,使盡可能多的孢子破壁以保證染液充分的進(jìn)入孢內(nèi),使染色充分。在染液未干之前,在體視鏡下拍照記錄。觀察孢子壁的層數(shù)、顏色、連孢菌絲的特征,通過與最新的分類信息(http://www.zor.zut.edu.pl/)比較,進(jìn)一步將初步聚類的孢子進(jìn)行物種鑒定。
1.5 統(tǒng)計分析
試驗數(shù)據(jù)用Excel進(jìn)行匯總整理,并用R語言進(jìn)行統(tǒng)計分析。所有數(shù)據(jù)在分析前都進(jìn)行正態(tài)性及方差齊性檢驗,對不符合整體分布的數(shù)據(jù)進(jìn)行l(wèi)g轉(zhuǎn)化。AM真菌孢子組成根據(jù)每個孢子類型的數(shù)量進(jìn)行相應(yīng)計算。采用雙因素方差分析檢驗?zāi)M增溫與不同植物對AM真菌孢子多樣性的影響,不同處理間的顯著性差異分析使用Post Hoc Test(Tukey法)在95%置信水平上進(jìn)行檢驗。基于Bray-Curtis距離,對AM真菌的孢子群落進(jìn)行主成分分析(Principal Co-ordinates Analysis,PCoA),使用“vegan”程序包中的“adonis”函數(shù)(PERMANOVA)檢驗?zāi)M增溫與植物物種對AM真菌孢子組成的影響。
2 結(jié)果與分析
2.1 AM真菌孢子的形態(tài)鑒定
本研究共分離獲得1 489個AM真菌孢子,按形態(tài)特征的不同可分成17種AM真菌形態(tài)種(圖1)。這17個種分別屬于8個屬,其中球囊霉屬(Glomus)6種,無梗囊霉屬(Acaulospora)2種,巨囊霉屬(Gigaspora)1種,盾巨囊霉屬(Scutellospora)3種,平囊霉屬(Pacispora)2種,內(nèi)生囊霉屬(Intraspora)1種,內(nèi)養(yǎng)囊霉屬(Entrophospira)1種和雙孢囊霉屬(Ambispora)1種。在數(shù)量方面,金黃球囊霉(Glomusaureum)的孢子數(shù)量最多(288個),其次為Glomuscaesaris和淺窩無梗囊霉(Acaulosporalacunosa),孢子數(shù)量分別為249個和170個。
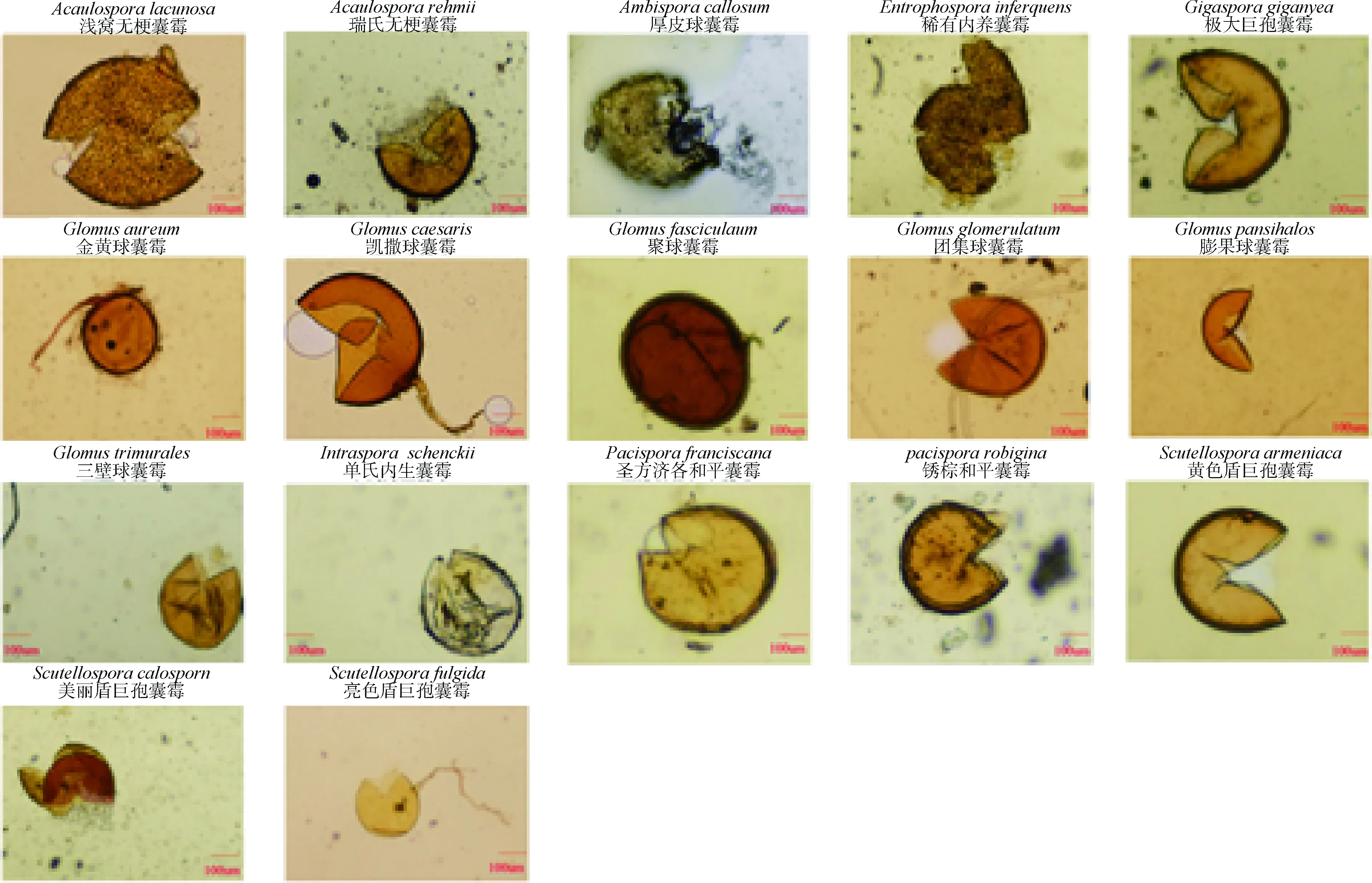
圖1 不同植物根際AM真菌種類
2.2 模擬增溫及植物物種對AM真菌孢子的影響
模擬增溫處理后,AM真菌孢子的豐富度并沒有顯著變化(圖2),但模擬增溫顯著降低了AM真菌孢子的密度(P<0.05)。AM真菌孢子豐富度及孢子密度在不同宿主植物物種間差異顯著(P<0.05)。如圖3所示,珠芽蓼根系A(chǔ)M真菌孢子的豐富度顯著高于甘肅棘豆、線葉龍膽、麻花艽和金露梅根系A(chǔ)M真菌的孢子豐富度(圖3)。此外,植物物種及增溫處理的交互作用對AM真菌的孢子豐富度和密度均沒有顯著影響。在所調(diào)查的15種植物中,矮嵩草(F=13.71,P=0.004)和線葉龍膽(F=12.46,P=0.005)兩個植物物種根際的AM真菌孢子密度在模擬增溫后均顯著降低(P<0.05);但AM真菌的孢子豐富度在不同物種中都沒有顯著變化。
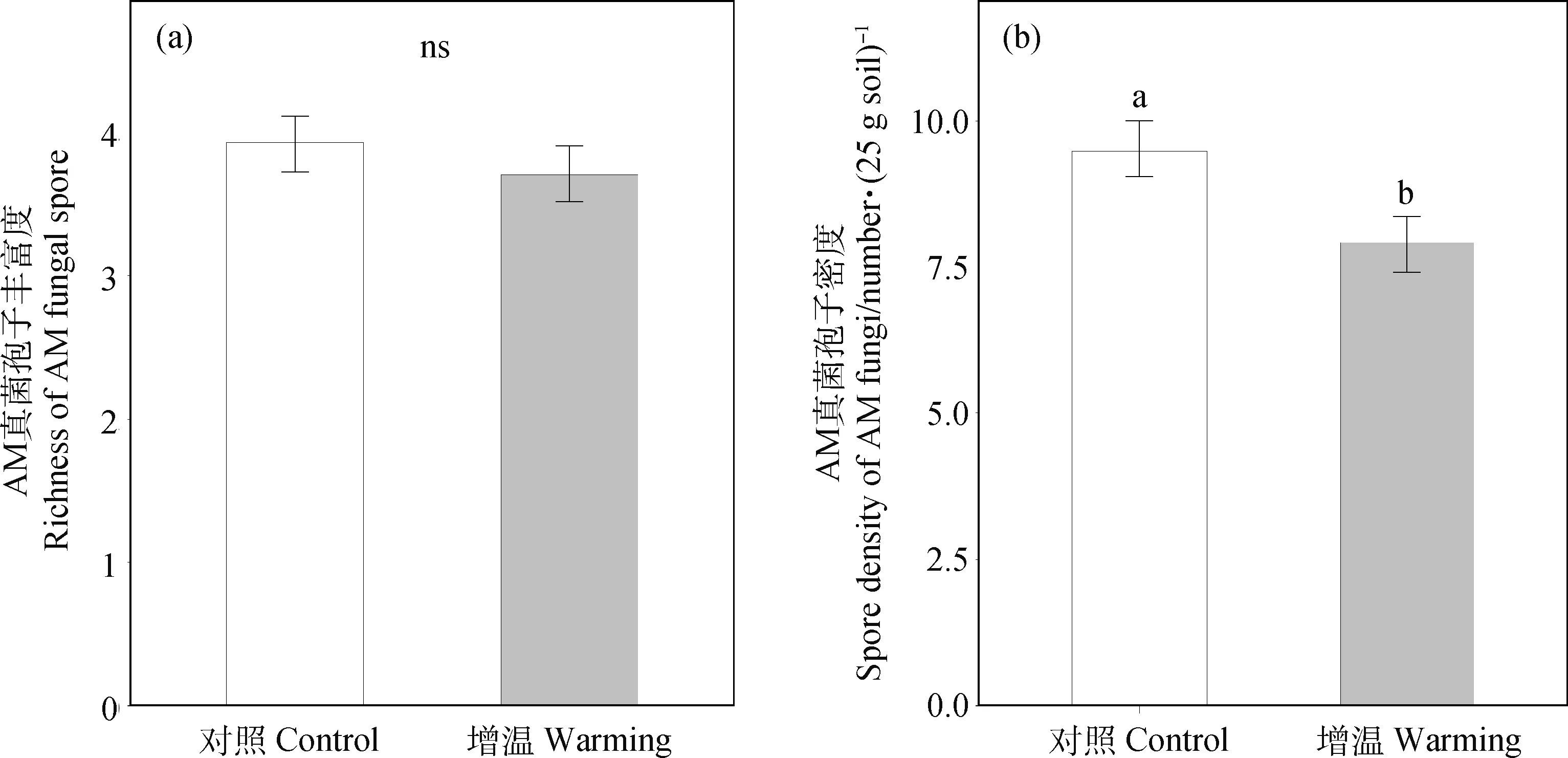
圖2 AM真菌孢子豐富度(a)及密度(b)在不同處理間的差異
AM真菌孢子組成的PCoA分析和PERMANOVA結(jié)果表明:增溫處理(F=1.26,R2=0.007,P=0.26)、植物物種(F=0.99,R2=0.08,P=0.52)及二者的交互作用(F=0.82,R2=0.07,P=0.90)均對AM真菌孢子組成無顯著性影響(圖4)。此外,在不同植物物種水平上,模擬增溫同樣也沒有引起AM真菌孢子組成的顯著變化。另一方面,本試驗結(jié)果發(fā)現(xiàn),AM真菌的孢子組成在不同植物物種間并沒有顯著性的差異(圖5a),但在不同植物科之間差異顯著(F=2.02,R2=0.14,P=0.001,圖5b)。
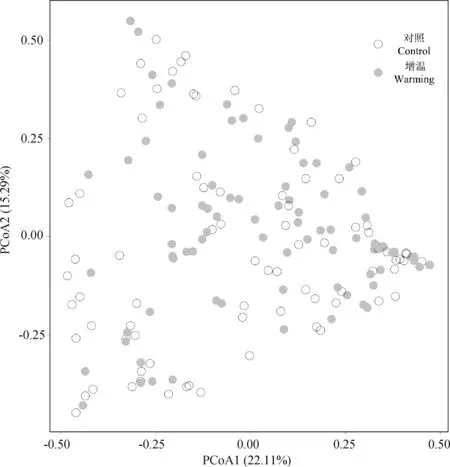
圖4 不同處理下植物根際AM真菌孢子的PCoA分析
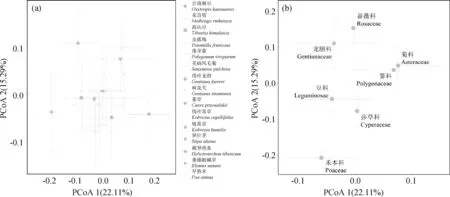
圖5 不同宿主植物在物種(a)和科(b)水平上根際AM真菌孢子的群落組成
3 討論
本研究在植物物種水平上探究了短期增溫對根際AM真菌孢子組成的影響,研究結(jié)果將豐富我們對青藏高原AM真菌孢子多樣性及其響應(yīng)全球變暖的認(rèn)識。研究結(jié)果共鑒定出17種AM真菌,分屬8個屬,孢子群落與青藏高原其他的研究結(jié)果類似。Gai等分別在莎草科植物的根際土壤和3種不同類型草地的根際土壤中檢測出26[32]和23[33]種AM真菌。這些研究結(jié)果暗示了青藏高原這一環(huán)境獨特的生態(tài)系統(tǒng)中具有較為豐富的AM真菌多樣性。此外,我們發(fā)現(xiàn)美麗盾巨孢囊霉(Scutellosporacalospora)在這3個研究中均普遍存在,說明其可能是青藏高原的廣布種。
溫度升高往往可以增加植物根系的生物量[34],促進(jìn)植物分配給AM真菌更多的光合產(chǎn)物[35],所以增溫一般認(rèn)為會對AM真菌的生長產(chǎn)生促進(jìn)作用。然而本研究結(jié)果表明,在青藏高原高海拔地區(qū),短期增溫并不會增加AM真菌的孢子多樣性,甚至還會降低AM真菌的孢子密度。依托分子鑒定技術(shù)的許多報道也證明模擬增溫對植物根系內(nèi)[24]或土壤中[26]的AM真菌多樣性沒有顯著性影響。Hiiesalu等在2014年指出,AM真菌的多樣性與植物的多樣性呈正相關(guān)關(guān)系[36],而本研究采用的是單植物物種的盆栽試驗,植物物種的單一可能是AM真菌多樣性較為穩(wěn)定的原因。此外,有研究表明模擬增溫并不會顯著改變青藏高原植物的根系生物量或根冠比[37-38],因此,穩(wěn)定的AM真菌多樣性也可能與植物對短期增溫的不敏感相關(guān)。另一方面,本研究發(fā)現(xiàn)模擬增溫會降低植物根際AM真菌的孢子數(shù)量,這與Yang等在2013年的試驗結(jié)果并不一致,其研究表明4年的模擬增溫并不會影響AM真菌的孢子密度[23],這可能是由于植物物種組成的差異造成的,Yang等的試驗是以自然草地為研究對象,而本研究則是以單物種的盆栽為研究對象。在植物物種水平上,本研究發(fā)現(xiàn)只有在矮嵩草和線葉龍膽兩個物種水平上,模擬增溫才顯著降低AM真菌的孢子密度。這種植物物種間的差異或許與不同植物的生理特性相關(guān),未來的研究需進(jìn)一步探討其具體的內(nèi)在機(jī)理。在AM真菌物種水平上,本研究中AM真菌孢子密度的降低主要是由盾巨孢囊霉真菌孢子的豐度降低引起。王淼焱等在之前的研究中表明,盾巨孢囊霉真菌多分布于沿海等較為濕潤的生態(tài)系統(tǒng)[39],由增溫所引起的土壤含水量降低可能不利于盾巨孢囊霉真菌的生長。
AM真菌群落組成受氣候變化的影響在青藏高原生態(tài)系統(tǒng)中已被多次報道[23-26]。在本研究中,AM真菌群落結(jié)構(gòu)并不受模擬增溫的影響。一般認(rèn)為,AM真菌的群落結(jié)構(gòu)受宿主植物和土壤性質(zhì)共同影響,在本研究中,穩(wěn)定的植物生物量一定程度上解釋了AM真菌群落結(jié)構(gòu)的穩(wěn)定性。另一方面,本研究發(fā)現(xiàn)宿主植物在科水平上對AM真菌孢子組成具有顯著影響,但在植物物種水平上沒有顯著影響。AM真菌的宿主特異性已有較多報道[18-19],而本研究表明,植物科的信息可以更精確地預(yù)測根際AM真菌的孢子組成。這一研究結(jié)果與石國璽等人的發(fā)現(xiàn)相吻合,其研究發(fā)現(xiàn)AM真菌侵染率在親緣關(guān)系近的植物物種內(nèi)并無差異,而在不同植物科之間存在顯著差異[40]。這些研究結(jié)果支持Reinhart等的已有研究成果,表明目前已知的親緣關(guān)系并不能準(zhǔn)確地預(yù)測植物與AM真菌間的共生關(guān)系[41],這可能與植物在科水平上的生態(tài)位分化與AM真菌的共生具有更強(qiáng)的相關(guān)性有關(guān)。
現(xiàn)有研究表明:土壤中AM真菌的孢子一直處于產(chǎn)生和萌發(fā)的動態(tài)過程中[42],本研究對AM真菌的孢子采集處于植物生長季末期,雖然具有一定的代表性,但還需進(jìn)一步在更多采樣時間段進(jìn)行相關(guān)研究,以期能夠獲取對AM真菌響應(yīng)氣候變暖的全面認(rèn)識。此外,AM真菌雖然廣泛分布于土壤中,但卻具有不同的產(chǎn)孢能力,如無梗囊霉科和球囊霉科的AM真菌一般認(rèn)為具有較強(qiáng)的產(chǎn)孢能力[43-44]。因此,以孢子形態(tài)鑒定為手段的方法未必能全面反映AM真菌對氣候變暖的響應(yīng),未來研究需結(jié)合高通量測序等分子技術(shù)展開深入研究。
4 結(jié)論
綜上所述,本研究結(jié)果表明,青藏高原不同宿主植物根際的AM真菌孢子組成在植物科水平上存在顯著差異,模擬增溫雖然對AM真菌孢子的多樣性無顯著影響,卻顯著降低了AM真菌的孢子密度,不利于特定AM真菌物種的生長。本研究結(jié)果系統(tǒng)分析了不同植物物種水平上AM真菌對模擬增溫的響應(yīng),暗示了植物群落組成在影響AM真菌響應(yīng)氣候變暖中的重要性,為探究氣候變暖下AM真菌的空間分布格局提供了生態(tài)依據(jù)。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國塑料(2016年3期)2016-06-15 20:30:00
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
