外源水楊酸對(duì)水稻苗期生長(zhǎng)與防衛(wèi)相關(guān)基因表達(dá)的影響
2021-11-03 03:45:44戴遠(yuǎn)興呂明芳袁正杰嚴(yán)成其張恒木
浙江農(nóng)業(yè)學(xué)報(bào) 2021年10期
劉 寒,戴遠(yuǎn)興,呂明芳,袁正杰,李 靜,嚴(yán)成其,張恒木,*
(1.浙江師范大學(xué) 化學(xué)與生命科學(xué)學(xué)院,浙江 金華 321004; 2.浙江省農(nóng)業(yè)科學(xué)院 病毒學(xué)與生物技術(shù)研究所,浙江 杭州 310021)
自然界中植物常受到多種病原菌的侵害,在與病原菌的長(zhǎng)期斗爭(zhēng)過程中,植物進(jìn)化出了獨(dú)特的免疫系統(tǒng)。一方面,植物通過模式識(shí)別受體(pattern recognition receptors,PRRs)識(shí)別病原模式分子(pathogen-or micro-derived molecular patterns,PAMP或MAMP),激發(fā)PAMP免疫(PAMP-triggered immunity,PTI)反應(yīng),啟動(dòng)抗性相關(guān)基因表達(dá),阻止病原菌侵染;另一方面,病原體通過釋放效應(yīng)子(effector)減弱植物PTI反應(yīng),植物R蛋白識(shí)別效應(yīng)子,啟動(dòng)免疫反應(yīng)(effector-triggered immunity,ETI)[1-2],激活防御信號(hào),使信號(hào)從病原感染部位傳導(dǎo)至未感染部位,產(chǎn)生系統(tǒng)獲得性抗性(systemic acquired resistance,SAR)。水楊酸(salicylic acid,SA)是PTI和ETI過程中激活SAR的重要信號(hào)分子,大量研究發(fā)現(xiàn),外源SA處理能提高植物抵抗生物和非生物逆境脅迫[3-4]。
植物體內(nèi)SA存在2種生物合成途徑:一是異分支酸(ICS)途徑,為擬南芥合成SA的主要途徑;二是苯丙氨酸(PAL)途徑,為水稻合成SA的主要途徑[5-6]。健康擬南芥和本氏煙中SA含量均較低,每g鮮重中通常含0.03~1.00 μg SA[7-13]。擬南芥和本氏煙對(duì)外源SA敏感,0.1~1.0 mmol·L-1外源SA即可穩(wěn)定誘導(dǎo)其防衛(wèi)基因的表達(dá),增強(qiáng)其對(duì)真菌、細(xì)菌、病毒等病原菌的抗性[14-17]。單子葉模式植物水稻SA含量較高,每g鮮重通常含SA 8~37 μg[18-19],推測(cè)水稻對(duì)外源SA的敏感性可能與雙子葉植物不同。不同學(xué)者曾采用0.01~400 mmol·L-1外源SA處理水稻[20-25],但由于濃度跨度大,測(cè)試的基因不一致,難以總結(jié)出SA對(duì)水稻的生物效應(yīng),給SA的生產(chǎn)應(yīng)用造成困擾。水稻苗期是水稻生長(zhǎng)發(fā)育過程中對(duì)許多病原體和非生物逆境脅迫的敏感期,在此發(fā)育階段誘導(dǎo)抗性對(duì)水稻穩(wěn)產(chǎn)具有重要意義。SA是誘導(dǎo)植物抗性的重要信號(hào)分子,然而,其處理水稻苗的最適濃度及其對(duì)水稻苗期生長(zhǎng)、防衛(wèi)相關(guān)基因表達(dá)的影響還不清楚。為了分析外源SA對(duì)水稻苗期生長(zhǎng)和防衛(wèi)相關(guān)基因表達(dá)的影響,本研究通過梯度濃度SA處理苗期水稻,并采用分光光度計(jì)、qRT-PCR等方法分析了水稻苗期生長(zhǎng)、抗性相關(guān)基因表達(dá)水平與外源SA濃度的關(guān)系,為SA應(yīng)用和深入研究功能機(jī)制奠定基礎(chǔ)。
1 材料與方法
1.1 材料與試劑
本研究使用的水稻品種均為日本晴(Nipponbare),由本實(shí)驗(yàn)室繁種保存。水楊酸(SA,貨號(hào)為69-72-7)、丙酮等生化試劑購(gòu)自生工生物工程(上海)股份有限公司;Trizol購(gòu)于上海Thermo Fisher Scientific公司;反轉(zhuǎn)錄與定量試劑盒購(gòu)于上海翊圣生物科技有限公司。
1.2 水稻栽培與SA處理
將日本晴水稻種子經(jīng)75%乙醇和2%次氯酸鈉除菌后[26],置于浸潤(rùn)的濾紙上,37 ℃催芽48 h后用水稻全營(yíng)養(yǎng)培養(yǎng)液[27]培養(yǎng),于28 ℃、12 h/12 h(L/D)條件下培養(yǎng);期間每2 d換1次新鮮的水稻營(yíng)養(yǎng)液,培養(yǎng)2周,達(dá)2葉1心或3葉1心期(多種病原敏感期)后,利用帶有刻度的噴壺分別加入梯度濃度為0.1、0.5、1.0、2.0、4.0、8.0、16.0 mmol·L-1SA,充分噴施水稻苗期植株(每株2.0 mL),對(duì)照為1.6%乙醇。由于SA處理能快速誘導(dǎo)植物防衛(wèi)反應(yīng),因此本研究采集SA處理0、2、4、8、12、24、48 h樣品,樣品保存于-80 ℃?zhèn)溆谩?/p>
1.3 總RNA提取和定量分析
采用Trizol試劑,參照項(xiàng)聰英等[28]的方法提取水稻樣品總RNA;采用Hifair?Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR(上海翊圣生物科技有限公司)試劑合成cDNA,方法參照試劑盒說明書;采用Hieff?qPCR SYBR Green Master Mix (High Rox Plus)(上海翊圣生物科技有限公司)試劑在熒光定量PCR儀7900HT(Thermo Fisher)上進(jìn)行定量分析,反應(yīng)條件設(shè)置為95 ℃ 5 min;95 ℃ 15 s,56 ℃ 20 s,72 ℃ 30 s,40個(gè)循環(huán)。每組實(shí)驗(yàn)設(shè)置3個(gè)生物學(xué)重復(fù)和3個(gè)技術(shù)重復(fù),定量基因引物信息見表1。
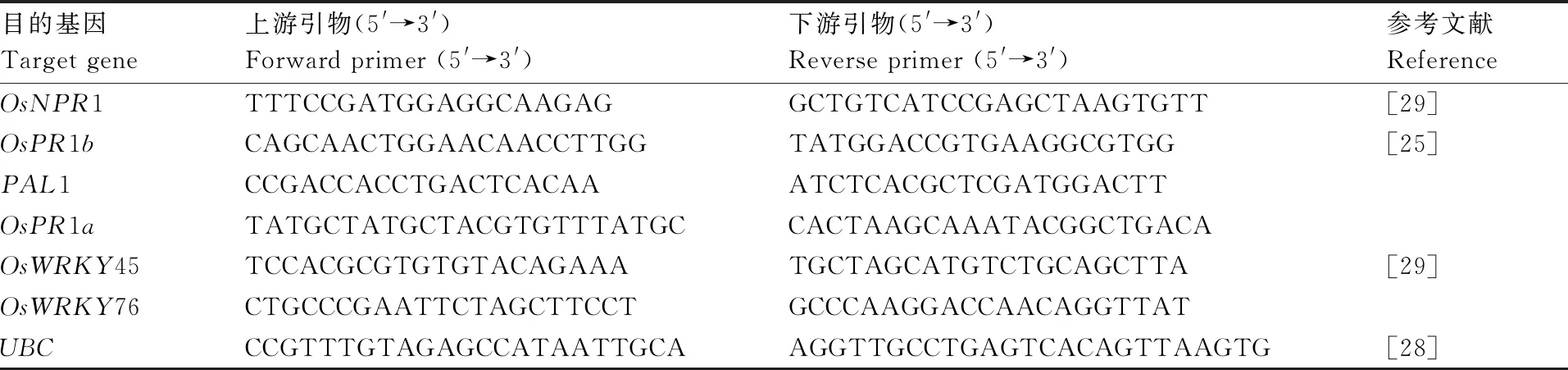
表1 qPCR采用的引物
1.4 水稻葉綠素含量分析
取新鮮水稻苗葉片0.5 g,加入2.0 mL抽提液(乙醇、丙酮、H2O體積比為4.5∶4.5∶1.0)進(jìn)行抽提,每組實(shí)驗(yàn)設(shè)置3個(gè)生物學(xué)重復(fù)和3個(gè)技術(shù)重復(fù)。以抽提液為空白對(duì)照,使用多功能酶標(biāo)儀Spect-raMax?M5(美國(guó)Molecular Devices公司)在波長(zhǎng)645 nm和663 nm處測(cè)定葉綠素提取物吸光度,然后參照修正后的Arnon法公式計(jì)算葉綠素含量[30]。
2 結(jié)果與分析
2.1 外源SA處理對(duì)水稻苗期生長(zhǎng)的影響
葉綠素含量是反映水稻苗生長(zhǎng)狀況的重要參數(shù)之一,采用分光光度計(jì)分析SA對(duì)水稻苗葉綠素含量的影響,結(jié)果如圖1-A所示。低濃度SA(≤2.0 mmol·L-1)處理,不影響水稻葉綠素含量,其中,1.0~2.0 mmol·L-1SA處理的水稻苗葉綠素含量在一定時(shí)間內(nèi)略微上升;中濃度SA(4.0 mmol·L-1)處理后,水稻苗葉綠素含量在24 h后開始下降,在48 h后下降顯著(P<0.05);而高濃度SA(>4.0 mmol·L-1)處理24 h后水稻苗葉綠素顯著(P<0.05)下降。進(jìn)一步觀察表型發(fā)現(xiàn),高濃度SA(>4.0 mmol·L-1)處理對(duì)水稻生長(zhǎng)影響較大,8.0、16.0 mmol·L-1SA處理24 h后水稻莖桿外周的葉鞘開始褪綠(圖1-B);隨著處理時(shí)間的延長(zhǎng),褪綠區(qū)域逐漸擴(kuò)大(圖1-C);處理72 h后,褪綠區(qū)域嚴(yán)重黃(白)化,葉片出現(xiàn)萎蔫(圖1-D);處理72 h后,4.0 mmol·L-1SA處理的水稻苗莖桿外周葉鞘也出現(xiàn)褪綠。表明高濃度SA處理可能通過抑制苗期葉綠素積累影響水稻苗生長(zhǎng)。
2.2 外源SA處理對(duì)SA自身合成的影響
植物體內(nèi)SA存在ICS (isochorismate synthase)和PAL (phenylalanine ammonia lyase)合成途徑,在水稻中PAL途徑占主導(dǎo)地位[31-32]。為了探究外源SA處理對(duì)水稻中SA合成的影響,選取水稻PAL途徑的關(guān)鍵基因OsPAL1進(jìn)行定量分析。結(jié)果(圖2)顯示,低濃度SA(0.1~4.0 mmol·L-1)處理,OsPAL1基因在8 h內(nèi)表達(dá)量顯著下調(diào)(約50%),12 h后其表達(dá)量呈逐步恢復(fù)趨勢(shì),與對(duì)照相比,其表達(dá)量恢復(fù)至80%以上,表明低濃度SA處理對(duì)OsPAL1表達(dá)影響較小。高濃度SA(尤其是8.0~16.0 mmol·L-1)處理水稻后,OsPAL1基因表達(dá)顯著下調(diào)(80%~90%)。上述結(jié)果表明,外源噴施SA可下調(diào)OsPAL1基因表達(dá),其下調(diào)幅度與外源SA濃度呈正相關(guān),推測(cè)高濃度外源SA可能抑制水稻內(nèi)源SA的生物合成。
2.3 外源SA對(duì)水稻SA受體基因表達(dá)的影響
NPR類基因編碼SA受體蛋白,其中,NPR1是關(guān)鍵基因[33-34]。因此,定量分析NPR1基因的表達(dá)量來研究外源SA對(duì)水稻SA受體基因表達(dá)的影響。結(jié)果(圖3)顯示,外源SA處理后水稻NPR1基因表達(dá)顯著上調(diào),在處理24 h時(shí)NPR1基因表達(dá)量達(dá)到最高,提升10~20倍,其中,2.0 mmol·L-1SA處理水稻苗24 h后,NPR1表達(dá)量最高。
2.4 外源SA對(duì)水稻W(wǎng)RKY基因表達(dá)的影響
WRKY類轉(zhuǎn)錄因子與SA誘導(dǎo)的抗性密切相關(guān),SA顯著誘導(dǎo)WRKY轉(zhuǎn)錄因子家族成員的表達(dá),調(diào)控SA信號(hào)途徑下游基因的表達(dá)[35-37]。本研究選取OsWRKY45、OsWRKY76來分析SA對(duì)水稻苗WRKY基因表達(dá)的影響,結(jié)果見圖4。其中,OsWRKY45對(duì)SA處理的響應(yīng)較慢,在處理早期(2 h)其表達(dá)量幾乎不受影響;當(dāng)SA濃度低于1.0 mmol·L-1時(shí),在12 h內(nèi),OsWRKY45表達(dá)量變化的幅度較小;處理24~48 h時(shí),OsWRKY45表達(dá)量顯著上升,其中1.0~4.0 mmol·L-1SA處理的OsWRKY45表達(dá)量較高,與對(duì)照相比上調(diào)8倍以上。SA濃度超過1.0 mmol·L-1時(shí),OsWRKY76的響應(yīng)較快,與對(duì)照相比,在2 h之內(nèi)上調(diào)2倍以上;隨著處理時(shí)間的延長(zhǎng),OsWRKY76的表達(dá)量穩(wěn)步上升,在24~48 h,OsWRKY76的表達(dá)量最高達(dá)4倍以上(圖4-B)。
2.5 外源SA對(duì)病程相關(guān)蛋白基因的影響
病程相關(guān)蛋白基因(PR基因)是一類重要的防衛(wèi)基因,且參與MAPK調(diào)節(jié)植物穩(wěn)態(tài)的過程,大量的研究表明,SA可影響PR基因的表達(dá)[38-40]。由圖5可知,0.1~16.0 mmol·L-1SA處理的水稻苗中OsPR1a表達(dá)量均上調(diào),隨著處理時(shí)間的延長(zhǎng),OsPR1a表達(dá)量也逐漸升高,處理24 h達(dá)到最高。其中,1.0~2.0 mmol·L-1SA對(duì)OsPR1a的誘導(dǎo)效應(yīng)最大;OsPR1b變化趨勢(shì)與OsPR1a不同,低濃度(尤其是0.1 mmol·L-1)SA使OsPR1b的表達(dá)上調(diào),而較高濃度SA(>1.0 mmol·L-1)處理時(shí)則顯著抑制OsPR1b的表達(dá)。
3 討論
SA可通過調(diào)節(jié)植物基因表達(dá)水平參與植物生長(zhǎng)發(fā)育、抵抗逆境脅迫[41]。本研究選取的6個(gè)基因均屬于SA生物合成或其抗性信號(hào)傳導(dǎo)途徑的關(guān)鍵基因,結(jié)果均表明外源SA能影響這些基因的表達(dá),影響效果與外源SA濃度密切相關(guān),反映了外源SA濃度的重要性。在擬南芥和本氏煙等雙子葉模式植物中通常采用的外源SA濃度為0.1~1.0 mmol·L-1[13-14];在水稻中,不同學(xué)者采用外源SA濃度差異較大[20-22,24-25,35,37],這給處理效果的比較與實(shí)驗(yàn)重復(fù)造成諸多不便。SA在水中的溶解度較低,難以用水溶液配制高于16.0 mmol·L-1的SA,若要配制更高濃度的SA需加有機(jī)溶劑(如乙醇),而過多的有機(jī)溶劑會(huì)給植物造成新的逆境脅迫[42];因此,本研究未采用16~400 mmol·L-1SA處理,采用0.1~16.0 mmol·L-1SA。2.0 mmol·L-1SA不僅能顯著誘導(dǎo)防衛(wèi)反應(yīng)信號(hào)通路(NPR1、WRKY45、WRKY76、PR1a)基因的表達(dá),且對(duì)葉綠素含量、SA合成產(chǎn)生的影響較小,這為后續(xù)外源SA處理提供了實(shí)驗(yàn)依據(jù)。8.0 mmol·L-1以上SA能夠顯著誘導(dǎo)防衛(wèi)反應(yīng)信號(hào)通路基因(NPR1、WRKY45、WRKY76、PR1a)的表達(dá),但明顯抑制水稻苗期葉綠素的積累,處理24 h后甚至可觀察到葉片褪綠、黃化、萎蔫等癥狀;但Xie等[25]采用400 mmol·L-1處理水稻,卻未見其報(bào)道如此高濃度的SA對(duì)水稻生長(zhǎng)產(chǎn)生不利影響,其原因尚有待進(jìn)一步探究。有研究顯示,高濃度SA降低小麥、擬南芥葉綠素含量,低濃度SA促進(jìn)葉綠素積累[6]。與此相似,本研究中1.0~2.0 mmol·L-1SA處理48 h,水稻苗葉綠素含量升高,高濃度SA(>4.0 mmol·L-1)處理,會(huì)抑制葉綠素的合成。綜合分析顯示,2.0 mmol·L-1外源SA處理苗期水稻較為合適,在實(shí)際使用時(shí)可根據(jù)其內(nèi)源SA含量適當(dāng)調(diào)整外源SA濃度,避免盲目使用造成人力物力浪費(fèi)。
猜你喜歡
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
軍事文摘(2021年22期)2021-11-26 00:43:51
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
 浙江農(nóng)業(yè)學(xué)報(bào)2021年10期
浙江農(nóng)業(yè)學(xué)報(bào)2021年10期
- 浙江農(nóng)業(yè)學(xué)報(bào)的其它文章
- 植物根系應(yīng)答鹽堿脅迫機(jī)理研究進(jìn)展
- 基因編輯技術(shù)在水稻抗病基因與育種研究中的應(yīng)用進(jìn)展
- 2022年《浙江農(nóng)業(yè)學(xué)報(bào)》征訂啟事
- 社會(huì)網(wǎng)絡(luò)視角的農(nóng)戶農(nóng)產(chǎn)品電商參與行為分析
——基于湖北、山東、安徽、甘肅349份農(nóng)戶抽樣調(diào)查數(shù)據(jù) - 中國(guó)化肥施用強(qiáng)度的驅(qū)動(dòng)因素分解與控制路徑研究
- 雙層平篩式半夏收獲機(jī)設(shè)計(jì)與試驗(yàn)