谷子GH5基因家族全基因組鑒定和表達分析
2021-11-03 03:45:46孟亞軒孫穎琦趙心月王鳳霞甕巧云劉穎慧
浙江農業學報 2021年10期
孟亞軒, 孫穎琦, 趙心月, 王鳳霞, 甕巧云, 劉穎慧
(河北北方學院 農林科技學院,河北 張家口 075000)
纖維素酶(cellulase)是能特異降解β-1,4-糖苷鍵的一類復合酶的總稱,在植物細胞壁的合成分解中發揮重要作用[1]。根據蛋白序列相似性,纖維素酶分布在至少17個糖苷水解酶家族(glycoside hydrolase,GH)中,不同GH基因家族具有不同的起源和演化過程[2]。GH5家族作為最大且功能最多樣化的GH家族,擁有最多的植物纖維素酶[2-3]。GH5蛋白由催化結構域(CD)和碳水化合物結合模塊(CMD)通過附件模塊(linker)鏈接組成,其結構上具有特異的TIM桶狀蛋白,具有保守型谷氨酸催化機制[4-5]。GH5家族成員具有廣泛的溫度和pH耐受性,其TIM桶折疊結構決定其loop元件具有高度可變性[6],故GH5基因家族可作為研究纖維素酶分子功能的良好素材。Henrissat等[7]首次將GH5描述為纖維素酶家族,并解讀其遺傳機制。隨著基因組學的發展,不同植物的纖維素酶基因功能也相繼得以揭示。研究發現,PtrCel9A6通過調控次生壁形成參與楊樹(Populus)木質部分化過程[8];Cel4基因參與番茄(Solanumlycopersicum)雌蕊和葉肉細胞的擴增作用,Cel1和Cel2在果實成熟與花藥開裂過程中具有重疊功能[9];擬南芥(Arabidopsisthaliana)纖維素酶基因CELLULASE6與角果分化有高度相關性,可通過改變CEL6酶的活性影響角果開裂進程[10];陳茹佳[11]研究發現,水稻(Oryzasativa)基因OsCel5-11與短日照條件下的晚花有關,GH5-11可能編碼一種新的內切葡萄糖聚酶。解纖維熱酸菌基因AcCel5A在擬南芥中過量表達會破壞其細胞壁結構,導致產生畸形植株[12]。王麗珊[13]利用生物信息學方法構建擬南芥和水稻Cel基因進化關系發現,Cel基因具有明顯的多樣性特征,蛋白功能存在特異性。
Science雜志在創刊125周年匯報中指出,植物細胞壁的合成將作為未來重大科學問題[14]。近年來關于細胞壁合成降解的相關研究已取得突破性進展,植物纖維素酶的作用機理已基本明確。谷子(Setariaitalica)屬1年生禾本科作物,基因組小且為二倍體,其測序已完成,基因組約500 M左右[15]。GH5具有大量寡糖、多糖等作用于β-鏈接的酶類,這些酶類在生物學過程中發揮重要作用。研究表明,GH5在植物細胞壁的合成和降解中發揮重要作用,但關于谷子GH5基因家族的研究卻鮮有報道。本文通過生物信息學方法對谷子GH5基因家族進行挖掘鑒定,分析其基因功能、蛋白結構、表達模式等,以期為進一步深入研究谷子GH5家族基因提供理論參考。
1 材料與方法
1.1 谷子GH5基因家族鑒定與亞細胞定位
利用Pfam數據庫獲得GH5基因家族Pfam代碼(PF00150)[16],在谷子基因組數據庫中獲得其基因與轉錄本ID,通過CDD[17]和InterProScan[18]進行篩選,去除冗余,并鑒定其基因信息,使用MG2C繪制染色體定位圖。利用ProtParam[19]預測蛋白理化性質,并通過Plant-mPLoc[20]進行亞細胞定位。
1.2 Motif獲取與進化樹繪制
利用MEME在線網站對谷子GH5蛋白家族序列信息進行分析,得到GH5的基序分布,最小長度設置60,最大長度設置200[21]。使用MEGA6.0軟件對GH5蛋白進行比對,使用銜接法繪制進化樹(bootstrap設置為1 000)[22-23],使用同樣方法繪制大豆(Glycinemax)、高粱(Sorghumbicolor)、水稻(Oryzasativa)等的多物種GH5蛋白進化樹,設置取默認值。通過Gramene數據庫檢索谷子與玉米GH5同源基因,利用KaKs_Calculator計算基因同義替換率(Ks)與非同義替換率(Ka),并計算Ka/Ks值,估算選擇壓力[24]。
1.3 GH5基因結構分析與結構域的獲取
利用Gramene數據庫獲取谷子GH5家族基因的編碼與全長序列,通過TBtools軟件分析并繪制其內含子-外顯子結構圖。為獲取GH5家族成員的結構域分布,使用在線軟件ProSite分析其蛋白序列并繪制谷子GH5家族成員結構域分布圖[25]。
1.4 GH5蛋白結構分析
通過PSRSM[26]在線軟件構建GH5蛋白二級結構。利用SWISS-MODEL網站分析其motif基序,構建蛋白三級結構[27],并運用相同方法完成大豆、高粱、水稻等GH5蛋白三級結構構建(以各位點出現頻率最高的氨基酸組成的序列構建不同物種GH5蛋白),使用SuperPose在線軟件比對各物種GH5蛋白的PDB格式文件,進行GH5蛋白三維結構對比。
1.5 谷子GH5家族基因表達分析
利用Phytozome數據庫中已公布的谷子RNA-seq數據,獲得谷子GH5基因在不同誘導下的組織表達譜,包括強光誘導2周的葉片、強光誘導1周的芽、黑暗誘導的地上組織、紅光誘導的地上組織、正常光誘導的根、干旱誘導的根、尿素誘導的根、強光誘導的穗,共11個樣本RNA-seq數據,使用TBtools軟件繪制谷子GH5家族基因在不同誘導下的表達熱譜圖[28]。
在GEO數據庫(GSE36391)和SRA數據庫(SRA048234)中獲得谷子品種張谷的轉錄組數據注釋文件,進行基因雙向同源比對,繪制張谷GH5家族基因在根、莖、葉、花穗中的表達熱譜圖。
1.6 GH5家族基因啟動子分析
使用Gramene數據庫提取谷子GH5基因起始密碼子上游1 500 bp序列,作為啟動子序列上傳至PlantCARE在線軟件,通過GSDS2.0將結果可視化[29]。
2 結果與分析
2.1 谷子GH5基因家族鑒定、染色體定位與結構分析
利用Pfam號碼(PF00150)從Pfam數據庫獲得谷子GH5基因家族隱馬文件,在谷子基因組中獲得GH5家族成員基因序列與轉錄本序列,通過CDD和InterProScan軟件去除冗余,最終獲得18個谷子GH5家族成員,命名為SiGH5-1~SiGH5-18。將得到的蛋白序列上傳至Plant-mPLoc數據庫進行亞細胞定位,使用ProtParam數據庫預測其蛋白質理化性質。通過對比GH5基因家族信息發現,GH5基因家族成員差異較大:具有2(SiGH5-10)~10(SiGH5-5、SiGH5-9)個外顯子,開放閱讀框長度為1 704 bp(SiGH5-2)~6 737 bp(SiGH5-18),氨基酸長度為285(SiGH5-14)~578 aa(SiGH5-10),分子量為31.69(SiGH5-14)~62.46 Ku(SiGH5-10),等電點為4.93(SiGH5-9)~9.34(SiGH5-7)(表1)。亞細胞定位分析發現,谷子GH5家族基因多位于細胞壁中,其次是細胞質,少量分布在葉綠體和細胞膜中,表明該基因家族參與細胞壁構建。
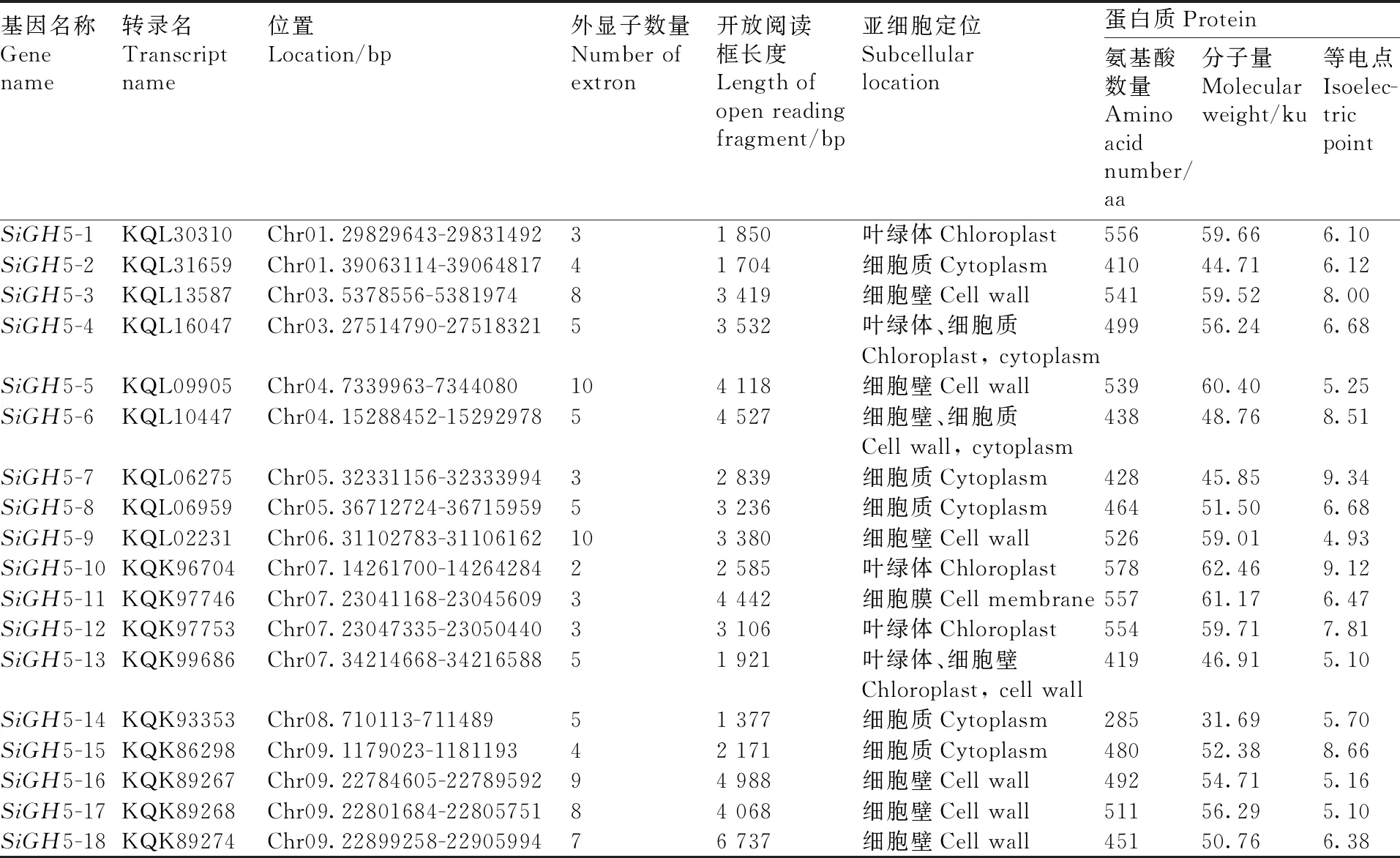
表1 谷子GH5基因家族信息
由圖1可知,18個GH5家族成員不均勻分布在谷子的8條染色體上,分布最多的為7號和9號染色體(各有4個),分布最少的為6號染色體和8號染色體(各有1個),1號、3號、4號、5號均含有2個GH5家族基因。
通過Gramene數據庫獲取GH5基因家族的全長序列,使用TBtools分析GH5基因結構,并將結果可視化繪制其內含子-外顯子結構。結果(圖2)表明,GH5家族基因均含有內含子,但差異較大,SiGH5-5、SiGH5-9內含子數量最多(均為9個),SiGH5-10內含子數量最少(1個)。
2.2 谷子GH5蛋白基序分析
利用MEGA6.0軟件對GH5家族成員氨基酸序列進行比對,并使用銜接法繪制進化樹;將比對文件上傳至MEME在線網站獲得其保守基序,最終獲得谷子GH5家族成員進化水平的基序分布圖(圖3)。GH5家族蛋白可分為3組,大部分蛋白分支具有100%的booststrap支持率。聚為同一分支的蛋白具有相似的基序分布,但組Ⅰ蛋白SiGH5-14缺失motif 3和motif 7;組Ⅲ蛋白SiGH5-18缺失motif 5,SiGH5-3缺失motif 3,推測在GH5家族蛋白分化過程中,SiGH5-14、SiGH5-18、SiGH5-3具有不同的進化軌跡。在預測到的8個motif中,motif 3幾乎分布在所有GH5家族成員中,可進一步用于GH5蛋白結構構建。
2.3 谷子GH5蛋白結構解析
通過在線軟件ProSite分析谷子GH5家族成員蛋白序列,并將結果可視化,得到谷子GH5蛋白結構域分布圖(圖4)。圖4表明,GH5蛋白均含有保守的GH5結構域,SiGH5-2除含有GH5結構域外還含有跨膜結構域,SiGH5-11含有RICIN結構域,SiGH5-18含有FASCIN結構域。蛋白的結構決定蛋白功能,這些特殊的結構域可能賦予GH5蛋白新的功能。
GH5蛋白二級結構(圖5-A)表明,GH5蛋白結構以無規卷曲為主要方式,具有81個,占40.7%;其次是α-螺旋(66個,33.17%);再次是延伸鏈(40個,20.10%);β-折疊僅有12個,占6.03%。通過SWISS-MODEL在線軟件預測各物種GH5蛋白三級結構,結果(圖5-B)發現,其三級結構主要由無規卷曲和α-螺旋構成,具有(β/α)8拓撲折疊。利用蛋白質疊加技術,在SuperPose網站進行GH5蛋白結構疊合對比,結果(表2)顯示,GH5蛋白在RMSD小于2?時具有高度的保守性(RMSD為疊加蛋白質骨架原子平均距離的量度,RMSD值越小證明兩物種蛋白結構相似性越高)。
表2 不同物種GH5蛋白三級結構的RMSD值
2.4 谷子GH5啟動子順式作用元件
利用谷子基因組選取GH5基因起始密碼子上游1 500 bp序列,提交至PlantCARE預測其順式作用元件,通過GSDS2.0將結果可視化(圖6)。生長素響應、脫落酸響應等激素類響應元件存在于所有GH5家族基因的啟動子中,光響應元件分布在除SiGH5-6外的所有GH5家族基因啟動子。此外,厭氧誘導元件、防御與應激響應元件、低溫響應元件等不同類型元件也不均勻分布在SiGH5s的啟動子區。上述結果說明,GH5家族基因可能在谷子生長分化與逆境脅迫過程中發揮作用。
2.5 谷子GH5基因表達分析
從Phytozome數據庫中獲得谷子18個GH5家族基因在不同誘導條件的RNA-seq數據,利用TBtools將結果可視化,繪制GH5家族基因誘導表達熱譜圖(圖7)。除SiGH5-1、SiGH5-2、SiGH5-16外,GH5家族基因在谷子各組織中均檢測到表達量。SiGH5-8在強光誘導1周的芽、紅光誘導的地上組織、強光誘導的穗、黑暗誘導的地上組織、干旱誘導的根中表達量明顯高于其他基因;SiGH5-17在正常光誘導的根、干旱誘導的根、尿素誘導的根、強光誘導生長2周的葉片、黑暗誘導的地上組織中表達量較高;SiGH5-3在黑暗誘導的地上組織中表達量較高;SiGH5-7在黑暗誘導的地上組織、強光誘導的穗、強光誘導1周的芽中表達量較高;SiGH5-9在強光誘導1周的芽中表達量較高。
為進一步研究GH5家族基因在谷子生長發育中的功能,利用GEO數據庫和NCBI的SRA數據庫獲得張谷的轉錄組注釋文件,并對基因進行雙向同源比對,繪制表達熱譜圖(圖8)。結果顯示,SiGH5-1、SiGH5-2、SiGH5-16在不同組織中均具有較低的表達量,說明這些基因可能具有特異的表達模式或為功能冗余基因。SiGH5-8、SiGH5-17在根、莖、葉、花穗中均具有較高表達量,結合誘導表達譜說明這2個基因在谷子生長發育和應對非生物脅迫過程中發揮重要作用。此外,除SiGH5-1、SiGH5-2、SiGH5-16基因外,其他GH5家族基因在根中表達量均相對較高,推測GH5家族基因在谷子根的建成中發揮重要作用。
2.6 多物種GH5蛋白系統發育樹分析
利用MEGA6.0軟件比對谷子、大豆、高粱等不同物種的GH5家族蛋白,利用銜接法繪制進化樹,設置默認值。結果(圖9)表明,大豆、擬南芥、小立碗蘚GH5蛋白呈家族性聚集,谷子、水稻、高粱、玉米GH5蛋白具有較高的親緣關系。說明GH5蛋白可能具有種屬特異性特點。
2.7 GH5家族選擇壓力分析
利用TBtools軟件對檢索到的谷子與玉米的13對GH5同源基因進行選擇壓力分析,結果(表3)顯示,13對同源基因的進化選擇壓力均<1,表明谷子與玉米的同源基因在進化中受到純化選擇,未發生結構功能上的改變。

表3 谷子、玉米GH5家族基因進化選擇壓力分析
3 討論
GH5基因家族在高等植物中廣泛存在,在植物細胞壁的合成和降解中發揮重要作用。植物全基因組測序的相繼完成為生物信息學研究搭建了數據平臺,但尚未有谷子GH5基因家族的研究報道。本研究從谷子基因組中鑒定到18個GH5家族基因,與曲霉(4個)[30]、大腸埃希菌(48個)[31]GH5家族基因的研究具有較大差異,推測在物種分化過程中GH5家族基因的特異性擴張導致不同物種之間差異比較大。基因重復是基因家族擴張的主要動力,GH5家族基因在谷子9號染色體中存在簇狀分布現象,說明GH5家族基因擴增中可能存在串聯復制。水稻、二歲短柄草中同樣存在GH5成員簇狀分布現象,還存在基因丟失事件[11]。
亞細胞定位與蛋白質功能具有密切聯系,本研究中大部分GH5成員定位在細胞壁,符合纖維素酶的一般特征,說明GH5成員大多數在細胞壁中參與植物的生理過程。進化水平的基序分布發現,GH5家族同組蛋白具有相似的基序分布,但部分蛋白存在基序缺失現象,這與擬南芥中的研究結果一致[4],推測在進化過程中GH5雖然保守但也發生了部分變異。
基因結構是基因進化的研究依據,從GH5基因內含子-外顯子結構可以看出,同一進化支的GH5家族成員雖然內含子和外顯子數量不同,但是具有相似的排列模式;結合其基序的近似分布,判斷GH5家族成員在進化過程中發生了外顯子改組。不同進化支亞組成員基因結構差異較大,說明GH5基因家族的不同亞組成員具有不同的進化軌跡[6]。
蛋白構建結果顯示,GH5蛋白結構并沒有因為物種的分化而出現較大的差異,說明GH5基因家族具有高度保守性。物種間蛋白進化樹分析表明,GH5蛋白具有種屬特異性特點,暗示單、雙子葉GH5蛋白為不同的起源方式,同源性較低[32]。
順式作用元件是解讀基因功能的重要依據,本研究中谷子GH5家族基因啟動子區預測到大量激素類響應元件,暗示GH5家族成員可能通過響應植物激素信號在調控細胞壁合成分解過程中發揮作用。基因表達分析結果顯示,SiGH5s在響應不同非生物脅迫過程中表達量不同,說明GH5家族成員在不同非生物脅迫過程中可能具有特異的調控途徑。轉錄組數據結果顯示,SiGH5s廣泛參與植株建成過程,但出現了明顯的組織表達差異,如SiGH5-8、SiGH5-17在根中高表達,SiGH5-7在穗中高表達,與水稻、擬南芥的組織表達結果一致[11],說明GH5家族成員在基因復制事件之后可能存在功能分化。
本研究基于谷子基因組和已公布的轉錄組數據,對谷子GH5家族成員進行鑒定和表達分析。結果表明,谷子中有18個GH5家族基因,不均勻分布在谷子8條染色體上,分為3個亞族,同一亞族具有相似的基序分布。SiGH5s在谷子不同組織中差異表達,且在非生物脅迫過程中不同組織的表達量也不相同。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
財經(2017年15期)2017-07-03 22:40:49
財經(2017年2期)2017-03-10 14:35:35
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
財經(2016年15期)2016-06-03 07:38:02
財經(2016年3期)2016-03-07 07:44:46
財經(2016年6期)2016-02-24 07:41:51
現代企業(2015年9期)2015-02-28 18:56:50
