小麥高配對材料CS-PhI的鑒定
2021-11-05 02:21:48裴自友程天靈溫輝芹辻本壽王宏兵張立生
種子 2021年9期
裴自友, 程天靈, 溫輝芹, 辻本壽, 李 雪, 王宏兵, 張立生
(1.山西農(nóng)業(yè)大學農(nóng)學院, 太原 030031; 2.鳥取大學干燥地研究中心, 日本 鳥取 680-0001)
染色體重組是對基因組重新排列組合的過程,對育種和遺傳研究都有重要意義。普通小麥(TriticumaestivumL.)是異源六倍體(2 n=6 X=42),染色體組為AABBDD。在普通小麥的減數(shù)分裂中只有同源染色體可以發(fā)生配對重組,這種類二倍化的減數(shù)分裂配對機制是受小麥5號染色體長臂上的主效基因Ph1控制。操控Ph基因誘導小麥-外源部分同源染色體配對重組,形成高遺傳補償性的染色體易位,是利用近緣屬種優(yōu)良基因的重要方式[1-3]。
有些擬斯卑爾脫山羊草(Ae.speltoides,SS)可以在5 B染色體存在的情況下提高小麥×擬斯卑爾脫山羊草雜種F1的部分同源染色體配對水平。原因是擬斯卑爾脫山羊草含有抑制Ph基因效應的顯性上位基因(PhI,又稱高配對基因),該基因能夠誘導普通小麥與小麥近緣種屬的部分同源染色體配對和交換[4-5]。Dvorak等[6]定位了擬斯卑爾脫山羊草中能夠抑制Ph1的兩個主效QTL(Su1-Ph1和Su2-Ph1),分別位于3 S長臂和7 S長臂上,利用Su1-Ph1抑制系統(tǒng)定向誘導部分同源染色體間的重組交換[7]。
Chen等[8]將擬斯卑爾脫山羊草中PhI基因轉(zhuǎn)移到普通小麥中國春上,育成含有PhI基因的普通小麥中國春高配對材料(Chinese Spring PhI,以下簡稱CS-PhI),利用該材料與硬簇麥雜交育成了小麥-簇毛麥T 5 VS·5 VL-5 DL易位系[9]。
目前對高配對材料CS-PhI的染色體組成僅進行了染色體C帶分析,初步定為小麥4 D易位[8],但尚無準確的定論,且沒有有效的連鎖標記。隨著技術(shù)的進步,利用基因組原位雜交(Genomic in situ hybridization, GISH)、重復序列探針雙色或多色熒光原位雜交(fluorescence in situ hybridization,F(xiàn)ISH)技術(shù)相結(jié)合可以更高效地識別染色體的身份[10-14]。
為了明確小麥高配對材料CS-PhI的分子細胞遺傳學組成,本研究利用基因組原位雜交和簡單重復序列(AAG)5為探針的FISH技術(shù)鑒定CS-PhI中小麥與擬斯卑爾脫山羊草的易位情況。同時開展白粉病抗性基因推導與成株期抗性鑒定,旨在了解其遺傳多樣性,為今后的利用提供科學依據(jù)。
1 材料與方法
1.1 材 料
小麥高配對材料CS-PhI由南京農(nóng)業(yè)大學細胞所提供,山西農(nóng)業(yè)大學農(nóng)學院優(yōu)質(zhì)小麥課題組保存。簡單重復序列(AAG)5由日本葛萊娜有限公司合成,擬斯卑爾脫山羊草和普通小麥中國春由日本鳥取大學農(nóng)學部植物遺傳育種實驗室提供。
用于苗期小麥白粉菌抗譜比較分析的39個攜帶已知抗病基因品種(系)、2個感病對照(阿夫和Chancellor)和23個不同毒性的小麥白粉菌菌株均由中國農(nóng)業(yè)科學院植物保護研究所周益林研究員提供。
成株期鑒定所用白粉病菌株為混合菌株,具體見文獻[15]。
1.2 方 法
1.2.1根尖有絲分裂中期染色體制片
參照郭明慧等[16]的方法進行根尖有絲分裂中期染色體制片。
1.2.2GISH分析
用tetramethyl-rhodamine-5-dUTP標記的擬斯卑爾脫山羊草基因組DNA為探針,普通小麥中國春基因組DNA作封阻DNA,雜交程序參照Cho等[17]的方法。
1.2.3順序FISH-GISH分析
參照Cho等[17]和溫輝芹等[18]的方法進行順序原位雜交,首先用tetramethyl-rhodamine-5-dUTP標記合成的簡單重復序列(AAG)5進行FISH分析,觀察、照相后將信號洗脫, 再以1.2.2方法進行GISH分析,用Olympus BX 61熒光顯微鏡對同一細胞觀察、照相。
1.2.4苗期抗白粉病基因推導
試驗在中國農(nóng)業(yè)科學院植物保護研究所溫室進行,具體參考曹學仁等[19]的方法。
1.2.5成株期白粉病抗性鑒定
試驗在山西農(nóng)業(yè)大學東陽試驗基地優(yōu)質(zhì)小麥課題組白粉病鑒定圃進行,具體參照程天靈等[15]的方法。
2 結(jié)果與分析
2.1 分子細胞學鑒定
根尖細胞有絲分裂中期染色體GISH結(jié)果表明,CS-PhI染色體數(shù)目2 n=42,含一對小麥與擬斯卑爾脫山羊草小片段雙末端易位染色體,易位位置為小麥染色體的端部,且易位染色體為一對隨體染色體(見圖1)。
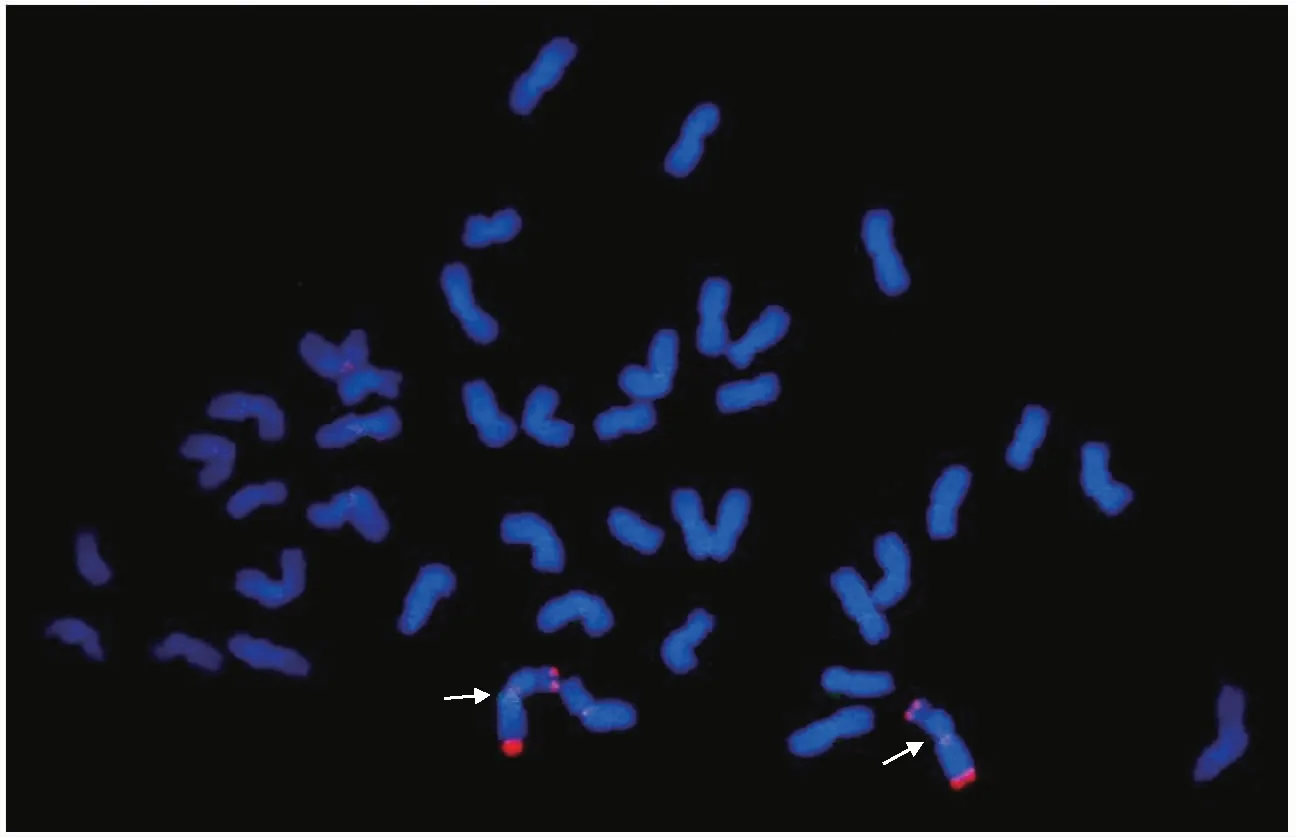
注:箭頭表示小麥-擬斯卑爾脫山羊草易位染色體。
對根尖細胞有絲分裂中期染色體同一制片進行順序FISH和GISH分析,探針分別為重復序列(AAG)5和擬斯卑爾脫山羊草基因組DNA,GISH時普通小麥中國春基因組DNA作封阻,結(jié)果見圖2。研究發(fā)現(xiàn),圖2-A箭頭所示易位染色體在著絲粒附近FISH帶型豐富,根據(jù)小麥(AAG)5標準帶型,進一步確定易位涉及的小麥染色體為6 B。圖2-B箭頭標示小麥-擬斯卑爾脫山羊草易位染色體,易位的擬斯卑爾脫山羊草染色體兩端均有GISH帶型(信號相對較弱)。

圖2 CS-PhI同一根尖細胞有絲分裂中期染色體順序FISH-GISH
2.2 白粉病抗性鑒定
2.2.1苗期小種抗性鑒定與抗性基因推導
基因推導結(jié)果(表1)表明,CS-PhI對3個小種表現(xiàn)免疫或高抗,對其他小種表現(xiàn)中感或高感。初步看,CS-PhI與Pm4c或Pm3b的抗譜有些相似,但考慮品種的背景不同,因此是否是新的基因還需進行綜合分析研究。
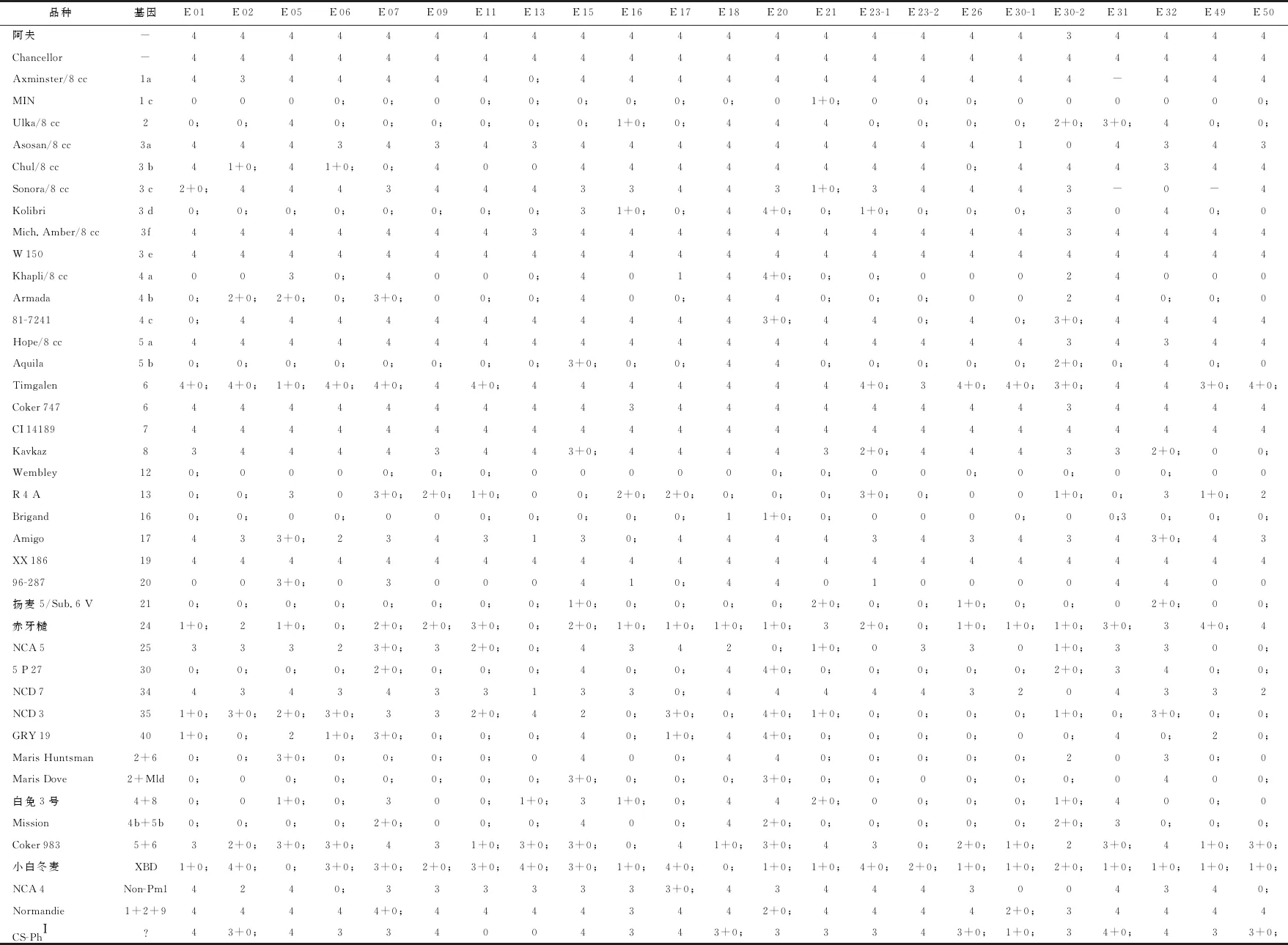
表1 對小麥白粉菌不同菌株的反應型
2.2.2成株期抗性鑒定
成株期白粉病抗病性鑒定結(jié)果表明,感病對照京雙16為8級,CS-PhI病級為0級,表現(xiàn)免疫。
3 討 論
以基因組DNA為探針的GISH推動了外源染色體鑒定,GISH和以單鏈寡核苷酸為探針的FISH技術(shù)的緊密結(jié)合,促進了染色體的精準鑒定,已廣泛用于小麥種質(zhì)材料分析[20-21]。
在小麥起源中由于S染色體組與B染色體組親緣關(guān)系較近,因此為了保證鑒定結(jié)果的可靠性,預備試驗中以S基因組為探針,對已知含殺配子染色體(gametocidal chromosome)的小麥-擬斯卑爾脫山羊草易位系CS-Gc1a(大片段易位)和CS-Gc1b(小片段易位)[22]進行準確的GISH鑒定。
本研究根據(jù)普通小麥中國春(AAG)5標準染色體分子核型[12-13],結(jié)合GISH結(jié)果,明確CS-PhI為雙末端小片段擬斯卑爾脫山羊草易位系,易位涉及的小麥染色體為6 B,尚不能確定易位的擬斯卑爾脫山羊草染色體為第幾條S染色體,因此,CS-PhI中易位染色體為T 6 BS·S/T 6 BL·S。對CS-PhI材料的識別和開展PhI基因的標記研究具有極其重要的價值。已建立了中國春與CS-PhI遺傳標記群體,并對F2單株分析染色體減數(shù)分裂中期染色體配對情況(含PhI基因會出現(xiàn)多價體),為進一步篩選與PhI基因緊密連鎖標記的分子標記奠定基礎(chǔ)。
Chen等[8]通過染色體C-分帶分析發(fā)現(xiàn),CS-PhI的4 D染色體異常,判斷其為小麥4 D與S染色體的易位系。本研究中易位的小麥染色體不是4 D而是6 B,推測可能是所用材料的差異所致。CS-PhI分不同的品系,組合來源均為(中國春×擬斯卑爾脫山羊草)/中國春BC2F3后代單株,GISH鑒定發(fā)現(xiàn)在鳥取大學辻本壽教授研究室保存的CS-PhI材料中沒有外源基因信號,可能為小麥-擬斯卑爾脫山羊草漸滲系(另文發(fā)表)。
目前小麥白粉病在育種和生產(chǎn)上面臨著抗源單一化和抗病性容易喪失的嚴峻現(xiàn)狀[23],因此有必要挖掘與利用不同基因資源,其中利用小麥成株抗性是未來實現(xiàn)品種持久抗性的最佳選擇[24]。劉成等[25]研究表明,來源于小麥近緣植物的抗白粉病基因有41個,其中包括來自擬斯卑爾脫山羊草的Pm12、Pm32和Pm53。本研究中,CS-PhI表現(xiàn)苗期對絕大多數(shù)白粉病生理小種感病、成株期抗病,為成株期抗病優(yōu)異材料,后期應進行白粉病抗性遺傳分析。
CS-PhI株高120 cm左右,穎殼有絨毛,有抽穗遲、晚熟、農(nóng)藝性狀較差,抗寒性弱等缺點,不宜直接作育種親本,為了利用其成株期高抗白粉病、耐熱等優(yōu)異特性,已通過與長麥6686和長7080等栽培品種雜交改良,培育出農(nóng)藝性狀優(yōu)良的抗白粉病新種質(zhì)。
