生境對藥食兩用植物接骨草的生長、葉綠素含量與葉綠素熒光特性的影響
2021-11-13 05:45:34陳蘭英許航徐正剛肖娟
生態科學 2021年5期
陳蘭英, 許航, 徐正剛, 肖娟 , *
生境對藥食兩用植物接骨草的生長、葉綠素含量與葉綠素熒光特性的影響
陳蘭英1, 許航2, 徐正剛2, 肖娟2, *
1. 西華師范大學, 生命科學學院, 南充 637000 2. 西華師范大學, 環境科學與工程學院, 南充 637000
探討接骨草對不同生境適應能力和響應機制, 以川東北丘陵地區野外自然分布的藥食兩用植物接骨草(Lind.)為研究對象, 研究林內(生境Ⅰ)、林窗(生境Ⅱ)、林緣(生境Ⅲ)不同生境下接骨草植株高度、葉片形態、葉綠素含量和葉綠素熒光特性等的影響, 結果表明: (1) 不同生境接骨草葉片的植株高度、葉片長、葉面積具有顯著的差異性, 林窗生境的植株最高, 葉片長、葉面積最大, 且與相對光強呈顯著負相關, 與濕度呈顯著正相關。(2) 葉綠素含量具有顯著的差異性, 與植株形態的生長具有一致性, 均是林窗生境顯著高于林內和林緣生境。(3) 不同生境葉綠素熒光參數差異顯著, 林窗生境的接骨草實際光化學效率(ΦPSII)值較高, 而非化學猝滅系數()較低, 同時較高的表觀電子傳遞速率(), 表明接骨草有較高光能利用能力和光呼吸能力。接骨草兼有陰生和向陽的特性, 在林窗生境下接骨草的長勢、葉綠素含量和光能利用能力均優于林內和林緣生境, 接骨草的植株形態、光合生理特征是對不同生境適應能力的主導因素。在引種栽培接骨草時, 林窗的相對光照(56.05±0.78%)和相對濕度(52.78±0.63%)可作為遮光種植和田間管理的參照。
接骨草; 植株高度; 葉片性狀; 葉綠素含量; 葉綠素熒光特性
0 前言
接骨草(Lind.)為忍冬科接骨木屬(Lind.)的草本或亞灌木, 因莖有八棱又稱八棱麻[1]。目前, 該種自然居群分布廣, 幾乎遍布北半球溫帶和亞熱帶地區, 在我國主要分布在秦嶺以南, 包括陜西、廣西、湖南、四川、貴州、浙江、江西、福建、臺灣等省區[2]。接骨草作為傳統中藥, 具有祛風利濕, 舒筋活血的功效, 可有效治療跌打損傷、風濕痹痛、筋骨疼痛及活血、行瘀、止痛等病癥[3]。除此之外, 接骨草還含有許多人體必需的微量元素、氨基酸和其他營養成分, 具有很高的營養保健價值, 可作保健食品的開發[4]。同時, 接骨草花序頂生聚傘圓錐花序, 外形美觀, 可用于觀賞; 還具有較強的抗環境污染能力, 在污染較嚴重的工礦區周圍也能良好生長[5-7]。因此, 接骨草具有極高的經濟開發價值和廣闊生態應用前景。目前對接骨草的研究多集中于藥用化學成分、藥理活性和臨床應用上的研究, 光合生理方面僅見董磊磊等[5]和王瓊等[8]在特定環境和短期時間下對接骨草進行光合-光響應和光合-CO2響應研究探討接骨草的光合特性, 而對接骨草的野外生存環境, 特別是連續時間下生境異質性對接骨草的生長、葉綠素熒光特性及葉綠素含量的影響研究還未見有相關的報道。前期的野外調查發現, 接骨草主要分布林內、溝旁、山坡雜草叢中, 不同的生境因子對其生長和生理特性可能產生不同的影響。植物的葉綠素含量、葉片形態會影響葉片光合效率, 同時又受到光照強度、氣溫、空氣相對濕度、土壤含水量等外界因素影響, 故揭示不同生境下接骨草的光合生理特性機制對深入了解接骨草的生態分布和移植栽培非常重要[9]。同時, 通過對接骨草生長特性的長期監測利用變異系數對不同生境接骨草的植株高度、葉片生長形態、葉綠素含量及葉綠素熒光特性的變異性進行評價是挖掘其有利基因并加以綜合利用的有效途徑, 也是種質資源研究最直接和最基本的方法[10]。因此, 本文在2017—2019年對3種不同的生境(林內、林窗、林緣)接骨草的葉片生長狀態、葉片葉綠素含量和葉綠素熒光特性進行測定, 并比較不同生境接骨草的差異并分析其變異性; 探討生境異質性和環境因子對接骨草生理生長的影響, 為今后深入研究接骨草精細化栽培及種植技術、提高接骨草的生態適應性提供理論基礎。
1 材料與方法
1.1 研究地及接骨草生境概況
實驗于2017—2019年5月在南充市西山風景區進行(30°48'54'' N, 106°03'47''), 該區域屬亞熱帶濕潤型季風氣候, 年平均氣溫17.4 帶, 年平均降雨量1021 mm, 該地植物的生長期較長, 年無霜期約300 d, 土壤為紅棕紫泥。在林緣、林窗和林內生境下分別選取有接骨草生長的20 m×20 m的固定樣方, 3個樣地的環境因子見表1, 3個樣地中相對光強差異呈極顯著(<0.01), 相對濕度具有顯著差異(< 0.05)。其中, 相對光強為林緣>林窗>林內, 相對濕度為林內>林緣>林窗。

表1 接骨草不同生境主要環境因子概況
注: Ⅰ, 林內; Ⅱ, 林窗; Ⅲ, 林緣。同列數據凡具有不同小寫字母表示在<0.05或<0.01水平上差異顯著, 下同。
1.2 植物生長情況及葉片葉綠素含量測定
在2017—2019年5月1日—10日選擇林緣、林窗、林內樣方下2 a生的接骨草(生長良好、生長基本一致)各10株進行標記, 對接骨草進行植株的高度、植株中部葉片長、寬生長情況測量, 葉面積用LI-3000C便攜式葉面積儀測定, 連續3 a測定的植株高度、葉片生長情況盡量保持同一植株、同一葉片。另在林緣、林窗、林內樣方中隨機標記10株接骨草植株, 每年從標記的10株接骨草植株中采摘鮮葉各300 g, 帶回實驗室進行葉綠素含量的測定, 葉綠素含量采用丙酮乙醇浸提法測定[11]。
1.3 葉片葉綠素熒光參數的測定
2017—2019年5月1日—10日選擇晴朗的3 d, 使用LI-6400便攜式光合儀測定葉綠素熒光特征參數。2017年每個生境隨機選5株, 每株測4—5片葉, 在2018年和2019年盡量選擇同株同片葉片進行測量, 3個生境共選擇15株植株。測定前將待測葉片進行暗適應20 min, 然后用弱光(0.1 μmol·m–2·s–1)照射, 測定初始熒光(F), 再照射飽和脈沖光(3000 μmol·m–2·s–1)2s, 測定暗適應最大熒光(F)。通過計算得到: PSII最大光化學效率F/F= (F–F)/F, 光合效率潛能F/F= (F–F)/F。在葉片充分光適應30 min后測定穩態熒光(), 最后照射飽和脈沖光(3000 μmol·m–2·s–1)0.8 s, 測出光適應最大熒光(F’)和表觀光合電子傳遞速率()。通過計算得到PSII光合量子產額PSII= (F’ –)/F’, 表觀電子傳遞速率= 0.58×φPSⅡ××0.5, 光化學淬滅系數()= (F’ –)/(F’–F’), 非光化學淬滅系數()= (F–F’)/F’[11]。
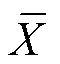
1.4 數據分析
采用SPSS 20.0統計分析軟件進行數據分析, 雙因素方差分析(Two-way ANOVA)用于檢驗不同年份和不同生境交互作用對接骨草生長、葉綠素含量和葉綠素熒光特性的影響; 同一時間不同生境之間的指標比較采用單因素方差分析(One-way ANOVA), 并采用LSD法進行多重比較, 采用相關分析和曲線擬合方法揭示環境差異因子對接骨草植株生長的影響, 使用Excel軟件作圖。
2 結果與分析
2.1 不同年份和不同生境下接骨草植株高度、葉片生長的動態變化差異及變異系數
3種生境下接骨草的植株高度、葉片生長均存在明顯的年份動態變化(圖1, 表2)。隨著年份的增長, 接骨草的植株高度、葉片長、寬和葉面積均增大。從重復測量方差分析結果來看, 不同生境對植株高度、葉片長、葉面積具有顯著的影響(<0.05), 而對葉片寬無顯著的影響(0.05), 不同年份對接骨草植株高度、葉片生長均有顯著影響(<0.05), 而生境和年份的交互作用對接骨草的植株高度、葉片生長無顯著影響(0.05)。
3種生境接骨草的植株高度隨著年份的增長均呈線性增長, 在葉片生長動態中, 2017—2019年植株高度、葉片長及葉面積隨著時間的增長不斷增加(圖1, 表2)。在2018—2019年, 林窗生境Ⅱ植株高度增長顯著大于林內生境Ⅰ和林緣生境Ⅲ(< 0.01), 均是林窗>林內>林緣。在葉片長中, 生境Ⅱ顯著大于林內生境Ⅰ和林緣生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ無顯著差異(>0.05); 在3個生境下, 葉片寬度生長情況無明顯區別; 葉面積在不同生境和不同年份均具有不同程度的差異(<0.05), 生境Ⅱ顯著大于其他兩個生境, 葉面積的增長基本上與葉片長呈一致性的規律。整體上, 3個生境的接骨草植株高度, 葉片長、寬和葉面積的變異系數均較低, 植株高度的變異系數變化幅度為1.376%—5.375%, 葉片長的變化幅度為1.005%—6.598%, 葉片寬的變化幅度為3.009%—12.872%, 葉面積的變化幅度為1.500%—7.966%, 這說明3個生境的接骨草的植株高度、葉片長和葉面積的表型性狀遺傳穩定性較高。
植物地上部分的生長會影響光能的獲取和水分利用, 同時植物通過對形態特征的適應調整去適應環境的變化, 從而獲得更多有效的資源[12-14]。3個樣地中相對光強和相對濕度具有顯著差異(表1), 為進一步表述不同生境中環境差異因子對接骨草表觀形態的影響, 對接骨草的植株高度, 葉片生長性狀和相對光強和相對濕度進行線性擬合。如圖2可見, 株高、葉長和葉面積與相對光強呈顯著負相關(p< 0.05), 葉寬與相對光強無顯著相關性(p>0.05); 株高、葉長、葉寬和葉面積與相對濕度均呈顯著正相關(<0.05), 這說明3個生境中具有顯著差異的環境因子濕度、光照條件影響接骨草的生長, 與相對光強的負相關和相對濕度正相關也進一步論證了接骨草屬喜陰性植物, 在其他自然條件無顯著差異的條件下, 光照強度越高越不利于接骨草的生長, 而濕度越大越利于接骨草的生長。
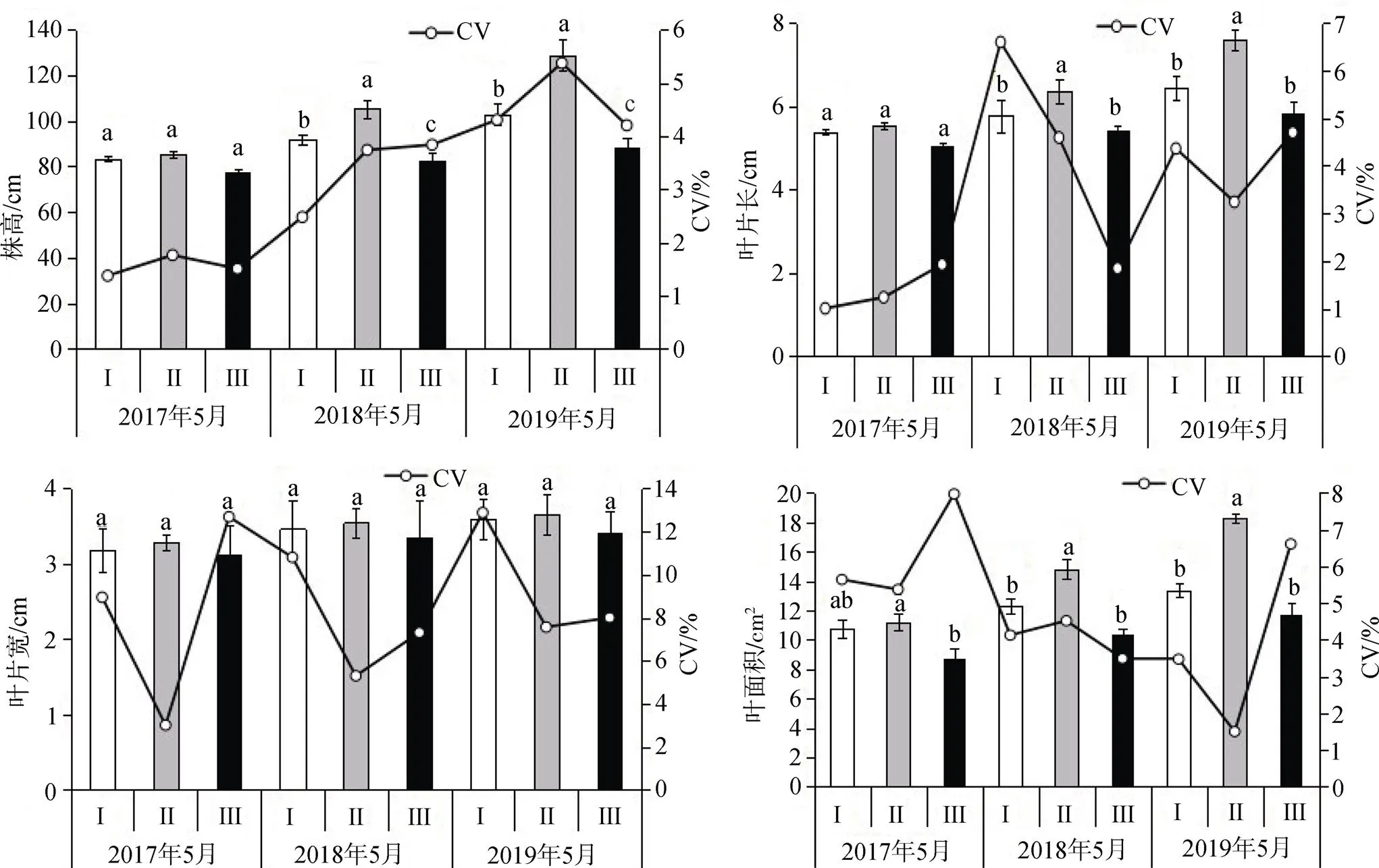
圖1 不同生境接骨草植株高度、葉片生長動態差異及變異系數
Figure 1 Comparison of plant height, leaf growth dynamics and variation coefficient ofin different habitats

表2 植株高度、葉片生長對不同生境和年份的雙因素方差分析
注: *表示差異顯著(<0.05), **表示差異極顯著(<0.01); 下同。
2.2 接骨草葉綠素含量的動態差異及變異性
3種生境下接骨草的葉綠素含量均具有顯著差異(表3)。不同年份、生境和年份的交互作用對接骨草葉綠素含量無顯著影響(0.05)。
葉綠素是維持植物光合作用的主要因子, 其含量和比例的變化是植物適應和利用環境因子的重要指標, 也是植物對環境脅迫的適應反應。由表3可知, 2017—2019年同一生境的葉綠素a()、葉綠素b()、總葉綠素(總)含量和葉綠素()無顯著差異, 但不同生境差異顯著(<0.05), 這表明不同的生境中的接骨草在吸收和利用光能方面差異較大(表4)。由表4可知, 在中, 生境Ⅱ顯著大于生境Ⅰ和生境Ⅲ, 生境Ⅰ和生境Ⅲ間無顯著差異;和總中, 均是生境Ⅱ>生境Ⅰ>生境Ⅲ(0.01); 在中, 生境Ⅱ>生境Ⅰ和生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ無顯著差異。變異系數中, 生境Ⅱ的變異系數均低于9%, 說明該生境植株葉片的葉綠素含量的變異性較低。

圖2 接骨草植株高度、葉片生長特征與環境差異因子相關性分析
Figure 2 Correlation betweenplant height, leaf growth characteristics and environmental difference factors

表3 葉綠素含量對不同生境和年份的雙因素方差分析

表4 接骨草不同生境葉綠素含量的差異及其變異性
2.3 不同生境接骨草葉綠素熒光參數的差異及變異性
通過對葉綠素熒光參數的測定, 可獲得植物光能利用途徑的信息[15-16]。重復測量方差分析的結果表明(表5), 3個生境的葉綠素熒光參數均達到了顯著的水平(<0.05或<0.01), 這表明接骨草在不同生境下對光能的利用有著顯著的差異。不同年份、生境和年份的交互作用對接骨草葉綠素熒光參數無顯著影響(0.05)。
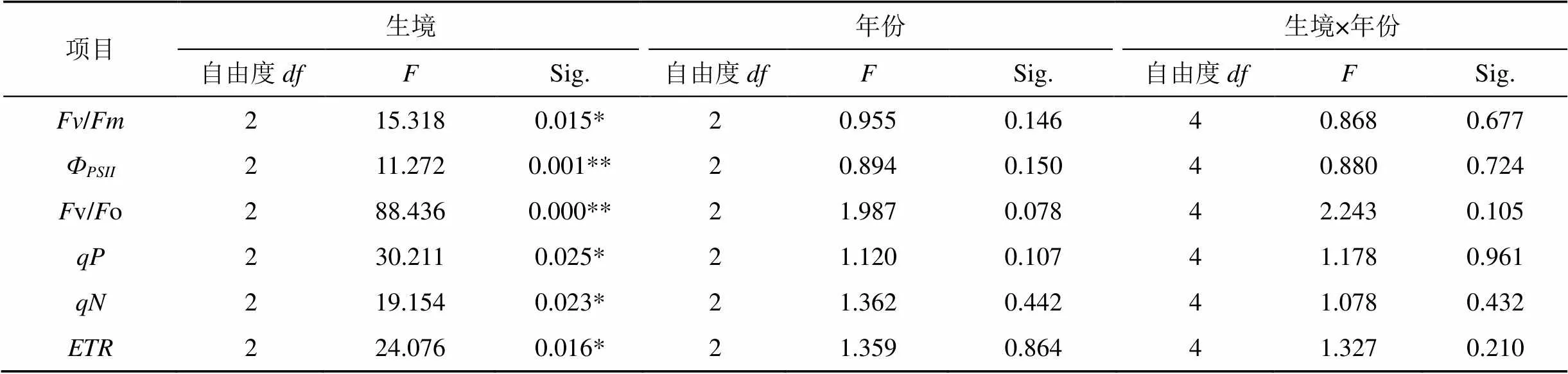
表5 葉綠素熒光特性對不同生境和年份的雙因素方差分析
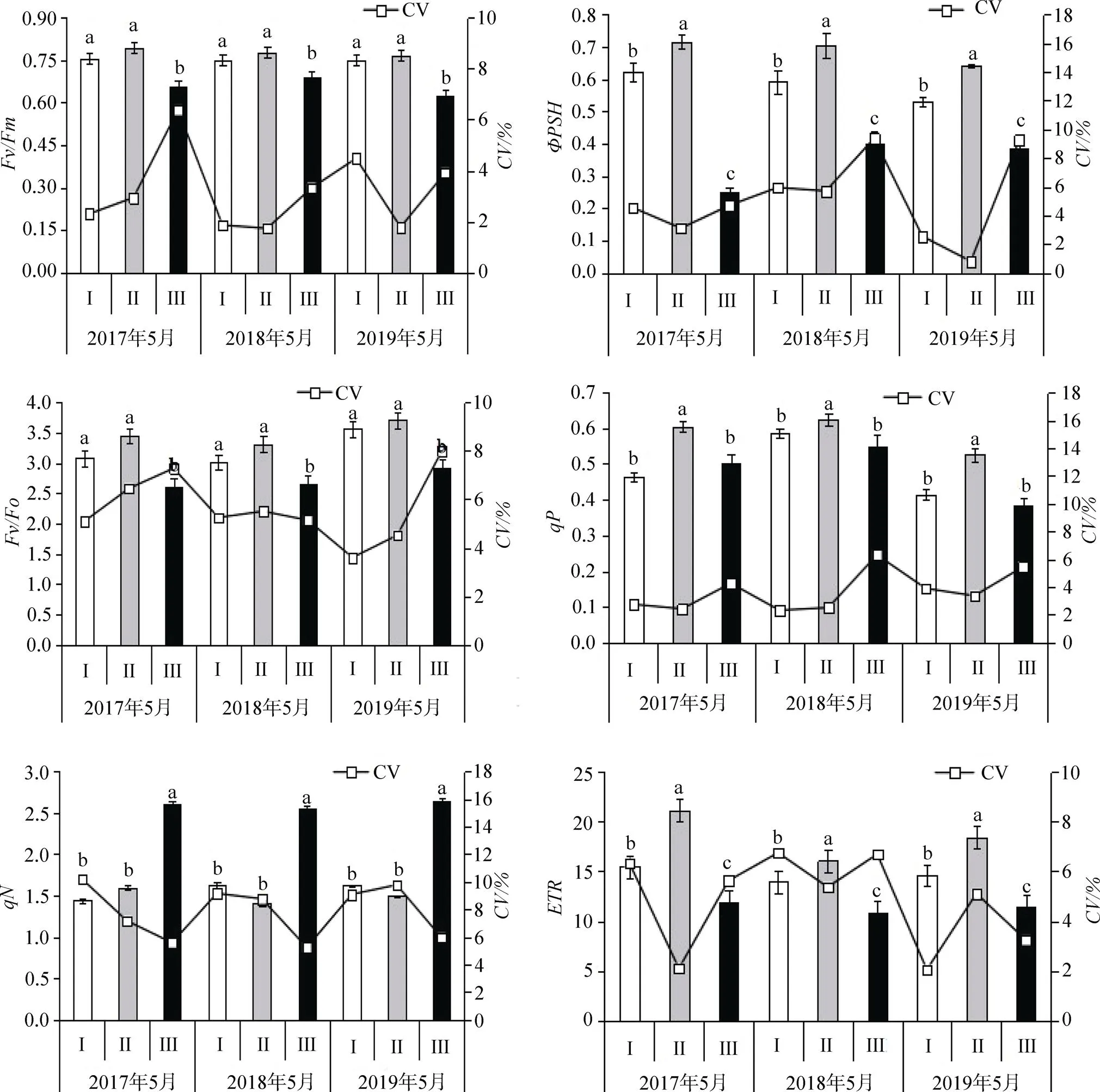
注: Fv/Fm: PSⅡ最大光化學效率, ΦPSa: 實際光化學效率, Fv/Fo: PSⅡ的潛在活性, qP: 光化學淬滅系數, qN: 非光化學淬滅系數, ETR: 表觀電子傳遞速率。
Figure 3 Differences in chlorophyll fluorescence parameters ofin different habitats
/反映是PSII反應中心最大光能轉換效率的反映指標, 反映PS中反應中心內稟光能轉換效率, 也是表征植物是否受到長期光脅迫的一個有效指標[17]。在不受光抑制的情況下,v/m一般介于0.75—0.85之間, 當值低于0.75, 越低則表明植物受光抑制的程度越高, 不利于植株的生長。由圖1可知, 接骨草林緣生境Ⅲ的v/m小于0.75, 在0.627—0.693范圍內, 說明該生境下的接骨草植株的生長受到光抑制, 生境Ⅰ和生境Ⅲ的v/m值無顯著差異, 且值均在0.75—0.85之間, 這說明在這兩個生境下生長的接骨草植株PSII 系統光能轉換效率正常。v/m在非脅迫條件下數值變化極小, 不受物種和生長條件的影響, 而在脅迫條件下明顯下降[18]。3個生境的在2017—2019年測定值中, CV的變化范圍為1.736%—6.373%, 這也進一步說明不同生境中接骨草v /m遺傳變異性較小, 接骨草長期生長在強光條件中會導致PSII 反應中心失活或受到破壞, 進而導致接骨草的光合作用受到光抑制。
反映植物實際的光合效率, 從分析結果可知, 不同生境接骨草的值具有極顯著差異(<0.01), 其中, 生境Ⅱ其生境Ⅰ境生境Ⅲ。在生境Ⅱ中值為0.640—0.715, 與v/m值相差約0.07—0.1; 在生境Ⅰ值為0.532—0.621, 與v/m值相差約0.135—0.219; 生境Ⅲ值為0.255—0.401, 與v/m值相差約0.292—0.404; 說明接骨草生境Ⅱ葉片所捕獲的光能轉化能植株所需的化學能的轉化效率高于生境Ⅰ和生境Ⅲ, 生境Ⅲ的轉化效率最低。
v/o表示PSII植物葉片潛在的光化學活性,v/m和v/o被絕大多數研究者認為是反映葉片光合效率的重要依據, 兩者的值越高說明植物葉片所捕獲的光能可以更加有效地轉化為植物所需要的能量[19]。不同生境接骨草的v/o值具有極顯著差異(<0.01), 與v/m的變化趨勢一致, 生境Ⅱ和生境Ⅰ無顯著差異, 生境Ⅲ最低。
反映天線色素吸收的光能用于光化學電子傳遞比例, 反映PSII開放程度及原初電子受體(Q)的還原情況。研究結果表明, 生境Ⅱ的值顯著大于生境Ⅰ和生境Ⅲ(<0.05), 生境Ⅰ和生境Ⅲ間無顯著差異, 這說明生境Ⅱ利用光能效果較生境Ⅰ和生境Ⅲ高。
反映了植物熱耗散的能力, 植物葉片出現非光化學淬滅是對光合機構保護的機制, 不同生境的接骨草的熱耗散能力差異較顯著(<0.05), 其中生境Ⅲ的值較高, 表明具有較強的熱耗散能力, 說明生境Ⅲ的接骨草在光環境下PSII天線色素吸收的光能用于熱耗散的比例顯著高于生境Ⅰ和生境Ⅱ光環境下的對應部分, 而用于光合的比例對應的會低于生境Ⅰ和生境Ⅱ光環境下的部分, 這可能是由于生境Ⅲ的接骨草生長狀態不如生境Ⅱ和生境Ⅰ的重要原因之一。
表觀光合電子傳遞效率值反映了實際光強下的表觀電子傳遞速率,值越高說明植株能將有效量子傳遞到光反應的能力越強。不同生境的接骨草值具有顯著的差異(<0.05), 生境Ⅱ>生境Ⅰ>生境Ⅲ。
3 討論與結論
植物在不同的生境條件下的形態特征的變化情況是反映植物對環境適應能力的重要指標, 葉片作為營養器官同時也是植物光合作用的主要器官, 為適應異質性生境常表現出較大的可塑性, 其形態及光合生理特征被認為能很好地體現對異質生境的適應[20]。通過對不同生境的接骨草的株高增量, 葉片長、寬和葉面積增量的比較后發現, 隨著年份的增加接骨草的株高、葉片性狀隨之增長。不對同生境同一測定年份進行單因素方差分析表明, 生境Ⅱ更利于接骨草的生長, 株高、葉片長、葉面積均顯著高于生境Ⅰ和生境Ⅲ。相關性分析表明, 植株高度、葉片長和葉面積均與相對光照呈顯著負相關, 株高、葉片長和寬、葉面積均與相對濕度呈顯著正相關(圖2); 這說明光強越大則不利接骨草的生長, 濕度越大越利于接骨草的生長。前人對接骨草的光合-光響應曲線及光合日變化進行研究發現, 接骨草是一種喜陰性的草本植物, 對弱光利用能力較強[5,8]。這與本文的研究結果一致, 接骨草在相對光強較高的林緣生境, 其株高和葉片生長特性均最小。在本文的研究中接骨草在林窗生境中株高、葉片長和葉面積最大, 這也進一步說明接骨草耐陰, 同時也具有向陽生長的特征, 對光的適應能力比較強, 分布廣泛。黃雅等人的研究結果表明不同產地的接骨草主要藥用成分總多酚依次為葉>全草>莖>根[21], 在本研究中林窗生境中植株較高和葉片較大說明接骨草在高度和廣度兩方面都增加了對資源的獲取和利用, 植物在環境壓力或資源有限條件下將分配更多的生物量到葉, 這樣能更有效地捕捉光能, 有利于接骨草次生代謝產物的合成及積累。同時, 3個生境的接骨草的株高、葉片生長性狀的變異系數較低, 表明其有較穩定的遺傳性, 生境Ⅱ的接骨草生長更好, 可從生境Ⅱ中進行引種栽培。
光合作用是植物生物產量的基礎, 葉綠素是植物光合作用必不可少的光催化劑, 其含量和比值常作為植物適應環境的重要評價因子, 它可以將捕獲的光能轉化為化學能, 在植物的光合作用過程中起著重要的作用, 葉綠素含量的變化與植物的生長發育有著密切的關系[22]。本研究中, 林窗生境下的接骨草生長較好, 其葉綠素含量也相對較高, 隨著光照強度的減弱, 接骨草的葉綠素a, 葉綠素b, 總葉綠素含量增加, 表明光強的降低可促使植株更好的吸收光能, 適度的林窗光照可以降低強光對葉綠素的破壞, 從而使接骨草葉片更加有效的進行光合作用, 這與蔣運生的研究規律一致[23]。高溫、強光和干旱脅迫會影響葉綠素的合成, 促進葉綠素的分解, 使其含量下降[24]。本研究中接骨草不同生境葉綠素相對含量具有顯著的差異(表3), 林窗生境Ⅱ的變異系數的范圍低于9%, 葉綠素含量較高的植株變異系數較低。光照強度在一定范圍內, 接骨草的葉綠素含量能夠達到最大值, 光照過弱或過強均會使葉綠素含量呈下降的趨勢, 林窗生境Ⅱ的光照范圍能夠使其達到最大值。
葉綠素熒光與光合作用反應過程緊密相關, 葉綠素熒光技術是一種快速、靈敏、無損傷的技術, 是光合作用的探針, 任何環境因素對光合作用的影響都可以通過葉綠素熒光反映出來[25-26]。反映PSII反映中心利用所捕獲激發能的情況, 被認為是植物發生光抑制最明顯的特征[27]。在3個生境中, 林緣生境的值低于0.75, 說明其生境的接骨草在生長過程中受到了光抑制。光合量子產量是用于衡量植物光合電子傳遞的量子產量, 可作為植物葉片光合電子傳遞速率快慢的相對指標, 在本研究中不同生境的實際的光合效率、潛在的光化學活性、光化學淬滅系數均表現出顯著的差異性, 表明不同的生境的植株對環境的適應能力不同, 這與張其德和任士福等人對相同植物不同生境的葉綠素熒光特性的研究結果一致[28-29]。在眾多熒光參數中, 高的v/m、v/o和PSII值已基本被認為是葉片高光合效率的重要依據, 且不少研究指出v/m、v/o和PSII有很好的一致性[30], 這與本研究的結果一致。大多數研究者認為, 非化學猝滅系數低的植物光合性能較好[31]。本研究中, 林窗生境和林內生境的接骨草PSII值較高, 非化學猝滅系數較低, 林緣生境的非化學猝滅系數較高, 這說明林緣生境的接骨草吸收的光能主要用于熱耗散; 因此, 其實際的光化學效率相對較低。本研究中, 生境Ⅱ林窗生境的v/m、v/o和PSII值較高,值較低, 其生境更利于接骨草的生長。
林窗、林內及林緣的水、熱、光等環境因子存在著顯著差異, 致使生長在不同光環境中的植物光合生理特征各不相同, 從而影響接骨草的生長。本研究中不同生境的接骨草植株生長高度、葉片生長特性、葉綠素含量和葉綠素熒光參數具有顯著差異。在林窗生境中, 接骨草的生長情況較好, 葉片的光合作用能力強, 對光適應生態幅較寬既耐陰也有向陽生長的特點, 接骨草在林窗的環境中競爭能力較強, 具有較高的潛在光能利用能力, 具有較好的生物學特性和生態適應能力。在林內的條件下表現也較好, 能正常生長, 但光合能力不如林窗生境。結合3種環境條件下接骨草的葉片葉綠素含量及光合生理特征, 可知在林窗條件下生長的接骨草光合作用能力更強、長勢更好。在引種栽培接骨草時, 應模擬野外生境Ⅱ的生長條件, 提供適宜的光照條件和充足的水分。在實際的栽培種林隙光照條件(56.05±0.78%)和相對濕度條件(52.78±0.63%)可作為遮光種植和田間管理的參照。
[1] 李勝華, 李愛民, 伍賢進. 接骨草化學成分研究[J]. 中草藥, 2011, 42(8): 55–57.
[2] 黃雙全, 王孝民. 食果鳥白頭鵯與接骨草的分布[J]. 植物學報, 2000, 42(10): 1096–1100.
[3] 姚元枝, 伍賢進, 黎曉英, 等. 接骨草的化學成分與藥理活性研究進展[J]. 中成藥, 2015, 37(12): 160–166.
[4] 趙湘婷. 中藥接骨草化學成分研究[D]. 蘭州: 蘭州理工大學, 2014.
[5] 董磊磊, 黎云祥, 權秋梅, 等. 中藥接骨草光合生理特性研究[J]. 草地學報, 2013, 21(4): 816–820.
[6] 陳志澄, 毋福海, 黃麗玫, 等. 砷礦尾砂污染及其治理研究[J]. 環境污染與防治, 2006, 28(3): 180–182.
[7] 班芷樺, 王瓊. 喜旱蓮子草和接骨草競爭對模擬增溫的響應[J]. 植物生態學報, 2015, 39(1): 43–51.
[8] 王瓊. 不同生境中入侵植物喜旱蓮子草與本地種接骨草的光合特性[J]. 西華師范大學學報: 自然科學版, 2015, 36(3): 253.
[9] 李月靈, 金則新, 王強, 等. 不同生境華東野核桃光合生理特性及葉綠素熒光參數比較[J]. 浙江大學學報(理學版), 2013, 40(2): 221–229.
[10] 張秦徽, 王洪武, 姜國云, 等. 紅松半同胞家系變異分析及選擇研究[J]. 植物研究, 2019, 39(4): 557–567.
[11] 王瓊. 喜旱蓮子草和接骨草光合色素含量測定方法研究[J]. 西華師范大學學報(自然科學版), 2009, 30(4): 18–22.
[12] HUGHES A P. Plant growth and the aerial environment : vi. the apparent efficiency of conversion of light energy of different spectral compositions by impatiens parvielora[J].Phytologist, 2006, 64(1), 48–54.
[13] ORLANDO G, EVA R, YAJAIRA G, et al. Influence of low light intensity on growth and biomass allocation, leaf photosynthesis and canopy radiation interception and use in two forage species of(DC. ) Benth[J]. Grass and Forage Science, 2018, 73(4): 967–978.
[14] BAGOES M I, OKA P, RAHMA H, et al. Factors affecting biomass growth and production of essential oil from leaf and flower of[J]. Journal of Essential Oil Bearing Plants, 2018, 21(4): 1021–1029.
[15] 李鵬民, 高輝遠, RETO J S. 快速葉綠素熒光誘導動力學分析在光合作用研究中的應用[J]. 植物生理與分子生物學學報, 2005, 31(6), 559–566.
[16] 李佳, 江洪, 余樹全, 等. 模擬酸雨脅迫對青岡幼苗光合特性和葉綠素熒光參數的影響[J]. 應用生態學報, 2009, 20(9): 2092–2096.
[17] 李孟洋, 巢建國, 谷巍, 等. 高溫脅迫對不同產地茅蒼術開花前葉片葉綠素熒光特征的影響[J]. 植物生理學報, 2015, 51(11): 1861–1866.
[18] 徐德聰, 呂芳德, 栗彬, 等. 不同品種美國山核桃葉綠素熒光特性的比較[J]. 果樹學報, 2008, 25(5), 671–676.
[19] 張守仁. 葉綠素熒光動力學參數的意義及討論[J]. 植物學報, 1999, 16(4): 444–448.
[20] 吳濤, 耿云芬, 柴勇, 等. 三葉爬山虎葉片解剖結構和光合生理特性對3種生境的響應[J]. 生態環境學報. 2014, 23(10): 1586–1592.
[21] 黃雅, 陳國華, 周欣, 等. 黔產接骨草中總多酚的含量測定及抗氧化活性研究[J]. 天然產物研究與開發, 2017, 29(2): 255–263.
[22] AND G H K, WEIS E. Chlorophyll fluorescence and photosynthesis: the basics[J]. Annual Review of Plant Physiology, 2003, 42(42): 313–349.
[23] 蔣運生, 柴勝豐, 唐輝, 等. 光照強度對廣西地不容光合特性和生長的影響[J]. 廣西植物, 2009, 29(6): 792–796.
[24] 梁永富, 王康才, 薛啟, 等. 高溫強光脅迫下水楊酸對多花黃精生理及光合特性的影響[J]. 南京農業大學學報, 2018, 41(5): 839–847.
[25] MAXWEL K., JOHNSON G N. Chlorophyll fluorescence —a practical guide[J]. Journal of Experimental Botany, 2000, 51(345): 659–668.
[26] 段龍飛, 尚愛芹, 楊敏生, 等. 中華金葉榆不同無性系光合特性和葉綠素熒光參數日變化研究[J]. 中國農業科技導報, 2014, 16(6): 21–27.
[27] GENTY B, BRIANYAIS J M, BAKER N R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J]. Biochimica et Biophysica Acta (BBA) - General Subjects, 1989, 990(1): 87–92.
[28] 王建華, 任士福, 史寶勝, 等. 遮蔭對連翹光合特性和葉綠素熒光參數的影響[J]. 生態學報2011, 31(7): 1811–1817.
[29] 張其德. 外加因子對生長在不同光強下的小麥葉綠體熒光特性的影響[J]. 生物物理學報, 1988, 4(3): 192–196.
[30] WANG Y, NII N. Changes in chlorophyll, ribulose bisphosphate carboxylase-oxygenase, glycine betaine content, photosynthesis and transpiration in leaves during salt stress[J]. Journal of Horticultural Science & Biotechnology, 2015, 75(6): 623–627.
[31] 徐曉昀, 郁繼華, 頡建明, 等. 2, 4-表油菜素內酯對亞適溫弱光下黃瓜幼苗光合特性和抗氧化系統的影響[J]. 核農學報, 2017, 31(5): 979–986.
Effects of different habitats on the growth, chlorophyll content and chlorophyll fluorescence characteristics of medicinal and edible plantsLind.
CHEN Lanying1, XU Hang2, XU Zhenggang2, XIAO Juan2, *
1. College of life science, China West Normal University, Nanchong 637000, China 2. College of Environmental Science and Engineering, China West Normal University, Nanchong 637000, China
Lind. is a traditional medicinal and edible plant; its health effects and ecological benefit values have been paid more and more attention. In order to explore the adaptive response mechanism ofin different habitats and make use of the wild resources, the plant height, leaf morphology, chlorophyll content and chlorophyll content and chlorophyll fluorescence in canopy gap, forest edge and understory were measured and compared in the hill region of northeast of Sichuan. The results were as follows: (1) The plant height, leaf length and area ofin different habitats were significantly different. In the canopy gap habitat was the highest and had significant negative correlations with relative light intensity, positively significantly correlated with humidity. (2) The chlorophyll content had significant differences, which was consistent with the growth of plant morphology. The canopy gap habitat was significantly higher than forest edge and understory. (3) The difference of chlorophyll fluorescence parameters between different habitats were significant different. The actual photochemical efficiency (Φ) in canopy gap habitat was higher, while the chemical quenching coefficient () was lower, and a higher apparent electron transport rate () indicating thathad high photorespiration.had the characteristics of combining shade plant and heliophyte, the canopy gap habitat was better than forest edge and understory, and the plant morphology and photosynthetic characteristics ofwere considered to greatly reflect the adaptation of plants to different environmental regimes. When introducing and cultivating, the relative light (56.05%±0.78%) and relative humidity (52.78%±0.63%) can be used as reference for shading cultivation and field management.
; plant height; leaf traits; chlorophyll content; chlorophyll fluorescence
10.14108/j.cnki.1008-8873.2021.05.009
Q948
A
1008-8873(2021)05-069-09
2020-04-03;
2020-05-15基金項目:國家青年科學基金(31700387); 西華師范大學青年資助專項(18D049)
陳蘭英(1987—), 女, 四川南充人, 碩士, 助教, 主要從事植物生態學研究, E-mail: 284626264@qq.com
通信作者:肖娟, 女, 博士, 副教授, 主要從事植物學、環境生態學研究, E-mail: xiaojuanhj@163.com
陳蘭英, 許航, 徐正剛, 等. 生境對藥食兩用植物接骨草的生長、葉綠素含量與葉綠素熒光特性的影響[J]. 生態科學, 2021, 40(5): 69–77.
CHEN Lanying, XU Hang, XU Zhenggang, et al. Effects of different habitats on the growth, chlorophyll content and chlorophyll fluorescence characteristics of medicinal and edible plantsLind.[J]. Ecological Science, 2021, 40(5): 69–77.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
