煙草細胞質雄性不育系SK01的胞質效應及特異性分子標記
2021-11-14 23:22:15劉艷芳,劉志文,孫玉合,楊愛國,李鳳霞
中國煙草科學 2021年5期
關鍵詞:煙草
劉艷芳,劉志文,孫玉合,楊愛國,李鳳霞
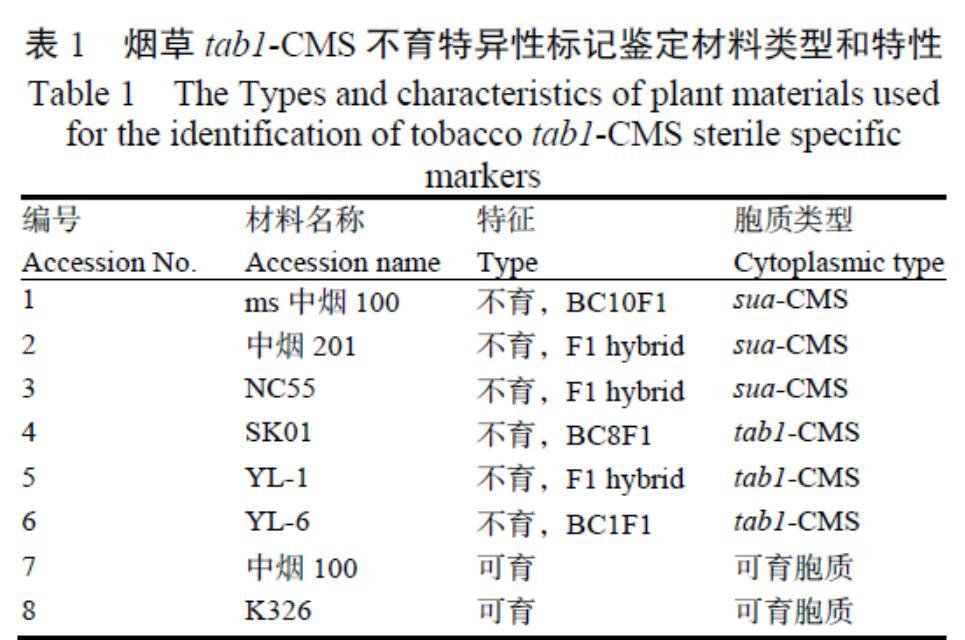

摘? 要:為了鑒定項目組發現的K326雄性不育自然突變株SK01與生產應用的不育胞質sua-CMS的異同,分析其在煙草育種和生產中的應用潛力,本研究以tab1-CMS不育系SK01和其同核異質系K326為母本,分別與4個烤煙品種雜交構建2組F1代雜交種,在兩個地點和人工氣候室中比較8個F1代以及SK01和K326之間的主要農藝性狀和抗病性差異,開展了tab1-CMS特異的DNA標記篩選。結果表明,SK01細胞質增強煙草雜交種F1代對CMV、黑脛病的抗性,對株高、莖圍等農藝性狀以及PVY、TMV抗性無顯著的細胞質效應;葉綠體基因組trnC-trnD基因區間,擴增到可有效區分煙草不育胞質類型的1個588 bp的特異性片段。本研究發現的不育胞質tab1-CMS與生產應用的sua-CMS類型不育系有明顯差異,對農藝性狀和抗病性無明顯的不利影響,是一種有一定應用潛力的不育細胞質新突變類型。
關鍵詞:煙草;細胞質雄性不育;農藝性狀;抗病性;特征標記
Cytoplasmic Effects and Specific Molecular Markers of Tobacco Cytoplasmic Male Sterile Line SK01
LIU Yanfang1,2, LIU Zhiwen3, SUN Yuhe1, YANG Aiguo1, LI Fengxia1*
(1. Tobacco Research Institute of Chinese Academy of Agricultural Sciences, Qingdao 266101, China; 2. Chinese Academy of Agricultural Sciences, Beijing 100081, China; 3. East China Normal University, Shanghai 201100, China)
Abstract: Cytoplasmic male sterile line (CMS line) is an effective genetic tool for tobacco hybrid breeding. At present, there is a single type of CMS used in tobacco hybrid breeding in China. In order to identify the similarities and differences between the male sterile natural mutant SK01 and the sterile cytoplasm used in production, and to analyze its application potential in tobacco breeding and production, tab1-CMS sterile line SK01 and its homokaryon heterogeneous line K326 were used as the female parents in crossing with four flue-cured tobacco varieties to construct two sets of F1 hybrids. Eight F1 lines, SK01 and K326 were compared in two locations in the field or in artificial climate rooms. The main agronomic traits and disease resistance differences between tab1-CMS-specific DNA marker screening were carried out. The results showed that the SK01 cytoplasm enhanced the resistance of tobacco hybrid F1 to CMV and black shank, and had no significant cytoplasmic effect on resistance to PVY and TMV, plant height, stem circumference or other agronomic traits. A 588 bp specific fragment was amplified from the trnC-trnD gene interval of the chloroplast genome, to that can effectively distinguish the sterile cytoplasm of tobacco. It shows that the sterile cytoplasmic tab1-CMS identified in this study is significantly different from the sua-CMS type sterile line used in production and has no obvious adverse effects on agronomic traits and disease resistance. It is a new type of cytoplasmicsterility with certain application potential.
Keywords: tobacco; cytoplasmic male sterility; agronomic traits; disease resistance; characteristic markers
細胞質雄性不育(cytoplasmic male sterility, CMS)是植物因雄蕊退化、花粉敗育或功能不育等原因造成的雄蕊不能正常授粉而雌蕊功能正常的一種母性遺傳性狀。雜種優勢是作物提高單產、改進品質、增強抗逆的重要途經[1]。利用雄性不育系生產雜交種可降低種子生產成本并保證種子的純度,水稻、十字花科蔬菜等多種植物的雜交種在生產中具有重要地位。煙草(Nicotiana tabacum)雜種優勢明顯,雜種F1代在生長勢、生長量、抗逆性以及品質等方面都比雙親平均值有明顯提高[2]。雜交種逐漸成為煙草品種培育的趨勢。目前煙草不育系和雜交種的種植面積占我國烤煙總面積的70%,但只有胞質來源于N. suavelens的sua-CMS類型不育系被廣泛使用。作物長期使用單一不育胞質遺傳基礎脆弱,易受病害蟲侵害,在生產上具有很大風險性。上世紀70年代美國玉米雜交種的不育胞質幾乎全部為T-CMS,由于T-CMS易感小斑病菌(Elminthosporium Maydis Shoemaker),造成了全美范圍內玉米的嚴重減產,T胞質被停用[3]。因此,急需開展煙草不育胞質創制、鑒定和評價等研究,打破煙草雜交育種中長期、大面積使用單一不育胞質的局面。
由于核質互作的影響,不育細胞質對作物多種性狀有顯著的細胞質遺傳效應[4]。水稻h2s細胞質對結實率、穗重、產量等性狀具有顯著的正效應[5-6],棉花不育細胞質對產量性狀具有明顯負效應[7]。在抗病性方面,據水稻、小麥、油菜等作物的相關報道,不同細胞質對抗病性一般表現為負效應,但多個外源胞質在小麥抗病性上表現為正效應[8]。煙草不育細胞質對后代的影響研究表明,與對照相比,不育胞質來源于N. bigelovii、N. megalosiphon、N. plumbaginifolia的不育系有效葉數減少;不育胞質來源于N. undulata的不育系開花期推遲[2],sua-CMS細胞質對煙草的農藝性狀和品質性狀沒有顯著的負效應[9]。
鑒定胞質雄性不育系類型的方法有形態學、細胞學和分子標記,其中分子標記是鑒定胞質雄性不育類型的主要方法[10-13],包括線粒體特異基因[14-16]、線粒體/葉綠體簡單序列重復(simple sequence repeat, SSR)[17-19]、單核苷酸多態性(single nucleotide polymorphisms,SNP)[20-22]等。葉綠體是與CMS有關的一個細胞器,在葉綠體基因組的基因間區和內含子區有大量的SNP、InDel、易位、重復、點突變以及基因組結構變異,具有比核基因組更高的核苷酸替代率[23],可用于區分細胞質雄性不育系[24-25]。葉綠體trnC-trnD約為3000 bp,包括trnC、petN、psbM、trnD 4個基因。該區域在多個物種中具有豐富的多態性,研究人員利用trnC-trnD序列構建了人參[26]、山礬屬[27]等物種的系統發育樹[28]。
SK01(tab1-CMS)是2010年7月在K326的試驗田中發現的一株雄性不育自然突變體,表現為完全敗育。2011—2016年間在青島和西昌兩地連續8代對SK01進行K326回交,回交后代均表現雄性不育,且為細胞質雄性不育。
本研究以SK01(tab1-CMS)及其同核異質系K326為母本、4個烤煙品種為父本配制8個雜交組
合,由此產生5組成對的細胞核基因組相同而細胞質不同的雜交種,以此評價SK01不育細胞質在煙草主要農藝性狀和抗病性等方面的胞質效應;通過比較tab1-CMS不育系、sua-CMS不育系及其可育對照的葉綠體trnC-trnD多態性,鑒定tab1-CMS的特定分子標記。這將為豐富煙草不育胞質源及煙草優異不育胞質在育種和生產上使用奠定基礎。
1? 材料與方法
1.1? ?試驗材料
1.1.1? 植物材料? SK01(K326的同核異質系,tab1-CMS),可育對照K326,以及以SK01(S)和K326(F)為母本,以中煙100(M1),NC82(M2),紅花大金元(M3)和Speight G-28(M4)為父本分別雜交的F1代。tab1-CMS組包括S以及S與M1-M4的4種雜交組合;對照組包括F以及F與M1-M4的雜交組合。
1.1.2? 供試毒株及菌種? 煙草花葉病毒(TMV)、黃瓜花葉病毒(CMV)、馬鈴薯Y病毒(PVY)、黑脛病菌種,均由中國農業科學院煙草研究所育種研究中心保存。
1.2? 試驗方法
1.2.1? 農藝性狀調查? 在山東省諸城市(北緯35°95',東經119°19',海拔65 m)和四川省西昌市(北緯27°84′,東經102°37′,海拔2100 m)兩個不同的煙草生態區分別種植SK01,K326和8個F1代雜交組合,采用完全隨機區組設計,設置2次重復,每個重復50株,在50%植株中心開放時,每個重復選取長勢相對均一的10個單株,按照煙草行業標準YC/T 142—2010測定株高、有效葉數、節距、莖圍、最大葉葉長和最大葉葉寬[29]。
1.2.2? 抗病性調查? 抗性鑒定在中國農業科學院煙草研究所人工氣候室進行,溫度為25 ℃,濕度為60%,每個材料設2次重復,每個重復25株。
TMV、CMV、PVY的抗性鑒定:將新鮮繁毒葉片研磨、過濾,濾液經純凈水5倍比例稀釋,待煙苗長至5~6片真葉時進行人工摩擦接種。接種30~35 d后,參照我國煙草病蟲害分級及調查方法B/T23222—2008進行抗病性調查。病情等級劃分為0、1、3、5、7、9六個等級。
黑脛病抗性鑒定:采用莖基部菌谷接種的方法,待煙苗長到4~5葉期時,在每株煙苗的莖基部用小刀輕輕劃一條大小相同的傷口,然后在根部接種3~5 g菌谷,立即覆土并灌水保濕。接種3周后調查每株發病情況,病害分級按國家行業標準YC/T39—1996規定執行,病情等級劃分為0、1、3、5、7、9六個等級。
1.2.3? 數據處理? (1)農藝性狀數據處理:使用SAS PROC GLM(版本9.3;SAS Institute Inc.)中的III型平方和固定模型和ANOVA進行數據分析。如果細胞質、細胞核、環境之間相互作用顯著,則采用固定效應的最小二乘法(LS-均值)分析環境和細胞核的細胞質效應。細胞質效應、細胞質與核及環境相互作用的多重比較采用Duncan多極差檢驗。tab1-CMS細胞質對農藝性狀的影響估計計算如下:[S/M(M1-M4)]F1-[異核同質系F/M(M1-M4)]F1[30]。
(2)抗病性數據處理:使用SAS 9.3軟件的協因素方差分析,對SK01,K326和8個F1代雜交組合進行病情指數分析(LSD分析p<0.05為有顯著差異)。
1.2.4? 特征標記擴增? 選用sua-CMS,tab1-CMS兩種不育類型的不育系以及可育煙草共8個材料(表1),用試劑盒提取葉片總DNA(TIANGEN Biotech,中國)。根據煙草葉綠體基因組序列(GenBank No:Z00044),設計兩對特異性引物(S1:5'-GAGTGGTAAGGCAGAGGAC-3'和5'-AAATCA ATGAAGGAAAAGC-3';S2:5'-TATCGCTTTTCC TTCATTG-3'和5'-ATTTTC TGCTAGATCCCGT-3')擴增葉綠體trnC-trnD基因間區域。PCR程序如下:94 ℃預變性5 min,PCR擴增30個循環,每循環94 ℃變性30 s,51 ℃退火30 s,72 ℃延伸45 s,最后72 ℃延伸7 min。用1%瓊脂糖凝膠電泳檢測擴增到的PCR產物。
使用PCR純化試劑盒(TIANGEN Biotech,中國)回收DNA片段,并將其連接到pMD18-T載體(TAKARA,日本)中。每個片段挑選5個陽性克隆進行測序,測序結果一致。根據sua-CMS、tab1-CMS和可育煙草中1738位點和2321位點的序列特征,用Primer 5設計特異引物S3:5'-AGGGGGCAGCAACTACAAT-3'和5'-CGGAAA AAATGATAAATAAAT-3'以擴增tab1-CMS的特異性標記。PCR擴增程序為94 ℃預變性5 min,PCR擴增30個循環,每循環94 ℃變性30 s,50 ℃退火30 s,72 ℃延伸30 s,最后72 ℃延伸7 min。用1.5%瓊脂糖凝膠電泳檢測擴增到的PCR產物。
2? 結? 果
2.1? tab1-CMS胞質對煙草農藝性狀的影響
在諸城、西昌兩個煙草生態區種植SK01(tab1-CMS)和可育對照K326,以及8對雜交組合,中心花開放期對其株高、節距、有效葉數、莖圍、最大葉葉長、最大葉葉寬等農藝性狀進行調查(表2)。
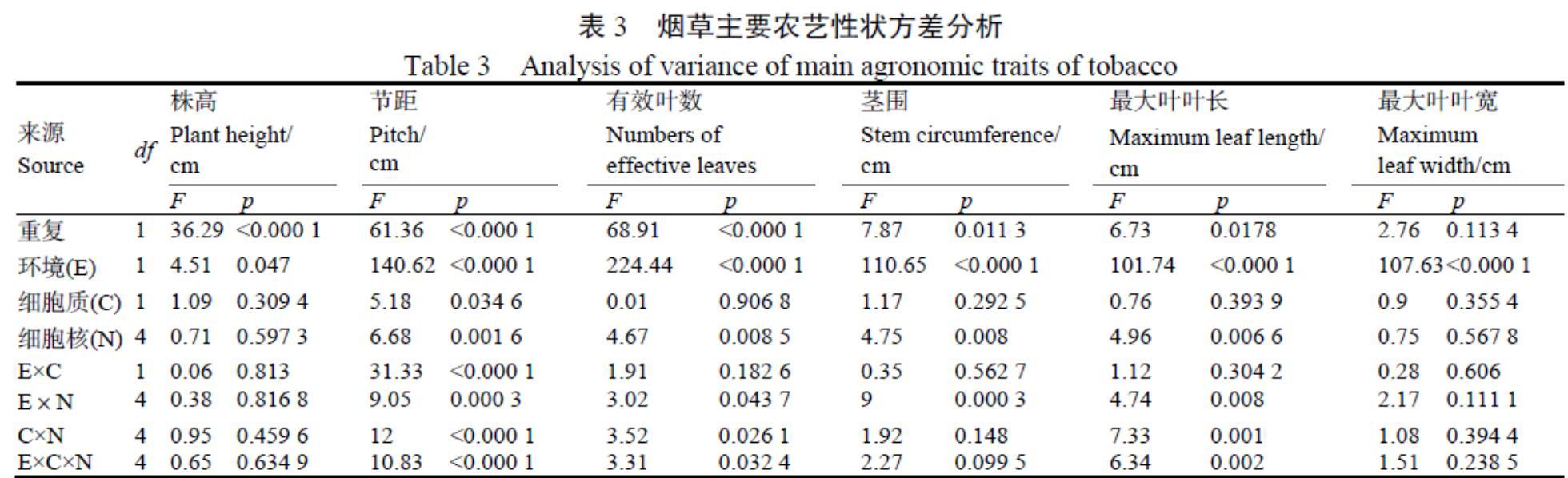
方差分析(表3)發現,tab1-CMS細胞質對所調查的農藝性狀除節距外無顯著影響,環境是影響農藝性狀的最重要因素,而核基因組對節距、有效葉數、莖圍和最大葉葉長等農藝性狀有顯著影響,對株高、最大葉葉寬無顯著影響。細胞質與環境互作對節距有顯著影響,細胞質-細胞核組互作、細胞質-細胞核-環境三因素互作均對節距、有效葉數、葉長等性狀有顯著影響。
2.2? tab1-CMS對產量性狀的胞質效應
對每一對具有相同父本的兩個雜交組合(例如S/M1-F/M1,S/M2-F/M2,S/M3-F/M3,S/M4-F/M4)以及S-F進行統計分析,可以估算tab1-CMS對煙草產量相關性狀的細胞質效應。結果表明(表4),在兩個煙草生態區,tab1-CMS的胞質效應不一致。在西昌生態區,tab1-CMS細胞質對產量相關性狀均無顯著的細胞質效應;在諸城生態區,tab1-CMS細胞質對大多數雜交組合的產量相關性狀均表現出細胞質效應,正效應和負效應的個數相同。對于有效葉數,tab1-CMS胞質在S/M1-F/M1、S/M2-F/M2、S/M4-F/M4三個組合上表現為正胞質效應,在組合S-F中表現為有負胞質效應,在S/M3-F/M3中表現為無顯著胞質效應。對于最大葉葉長,tab1-CMS細胞質在S/M1-F/M1、S/M2-F/M2兩個組合中表現為負胞質效應,在組合S-F中表現為有正效應,在其他兩個組合S/M3-F/M3、S/M4-F/M4中無顯著細胞質效應。對于最大葉寬,tab1-CMS細胞質在S/M1-F/M1、S/M3-F/M3兩個組合中表現為有負效應,在組合S-F中表現為正效應,在S/M2-F/M2、S/M4-F/M4中則無顯著細胞質效應。表明tab1-CMS的細胞質效應與其互作的核基因組有關。
2.3? tab1-CMS胞質對煙草抗病性的影響
抗病性是胞質評價的重要內容,對SK01和K326以及它們分別與M1、M2、M3、M4四個煙草品種的各雜交F1代的病情指數進行統計分析(表5),結果顯示,tab1-CMS細胞質對黑脛病抗性有顯著影響(p=0.0369),對CMV抗性有極顯著影響(p<0.0001),對TMV、PVY抗性無顯著影響。細胞質-細胞核互作對黑脛病、CMV有顯著影響,對TMV、PVY則無顯著影響。
比較父本相同、母本細胞核一致而細胞質不同的兩個F1代雜交種的病情指數差異(表6),可以看出當父本為中煙100(M1)、NC82(M2)時,F1代的黑脛病病情指數存在顯著差異,細胞質為tab1-CMS的F1代病情指數顯著低于可育胞質,表現出tab1-CMS胞質在黑脛病抗性上的正效應。在CMV抗性上,父本為中煙100(M1)時,tab1-CMS的F1代病情指數顯著低于可育胞質;SK01和K326兩個材料在黑脛病抗性和CMV抗性上均存在顯著差異。在TMV、CMV抗性上,細胞質不同、細胞核相同的成對雜交組合間病情指數的差異不顯著。
2.4? tab1-CMS特異的DNA標記
設計S1、S2兩對引物在sua-CMS(MS中煙100、中煙201、NC55)、tab1-CMS(SK01、YL-1、YL-6)兩種不育胞質類型的不育系和可育煙草(中煙100、K326)共8個煙草材料中擴增trnC-trnD的完整基因間區域,擴增結果(表7)顯示每個樣品的擴增產物單一,且同一引物在不同材料間的擴增長度一致,其中S1擴增長度為1492 bp,S2擴增長度為1756 bp。對每個片段的5個克隆測序結果表明每個PCR片段序列完全一致。而S1和S2的擴增區域在tab1-CMS不育系、sua-CMS不育系與可育煙草間存在25個多態性位點,含有相同細胞質的煙草材料序列具有100%的相似性(表7)。在可育煙草和tab1-CMS與sua-CMS之間存在16個差異位點,sua-CMS和可育煙草與tab1-CMS之間存在5個差異位點,tab1-CMS和sua-CMS與可育煙草之間存在3個差異位點。在3種細胞質中,trnC-trnD基因間區域的s1738位點各不相同。在s1738位點,sua-CMS不育系為TTTTATTCCAGG GGGG,tab1-CMS不育系為GGGGGCAA,可育煙草為GGGGGCAGCT TTATAA,而在s2321位點,sua-CMS不育系和可育煙草均為ATCATT,而tab1-CMS不育系該位點缺失。根據sua-CMS、tab1-CMS和可育煙草中trnC-trnD基因間區域中s1738和s2321位點的序列特征,設計特異性引物S3在tab1-CMS不育系中特異性地擴增到一個588 bp的片段,該片段在sua-CMS不育系和可育煙草中未得到擴增(圖1)。
3? 討? 論
煙草不育系或雜交種的種植面積占總種植面積的70%以上,但不育胞質來源單一,急需擴大不育胞質來源。本研究對一個新的不育胞質tab1-CMS在農藝性狀和抗病性上的胞質效應進行評價,結果顯示tab1-CMS細胞質對農藝性狀無顯著影響,對黑脛病、CMV抗性具有正效應,對TMV、PVY無顯著影響,表明tab1-CMS表現優異,有望用于煙草雜交育種和不育系轉育。
農藝性狀是品種選育的基礎。在本研究中,一年兩點的試驗結果表明tab1-CMS細胞質對煙草株高、節距、莖圍、最大葉葉長、最大葉葉寬,有效葉數等農藝性狀無顯著影響,但在西昌、諸城兩個煙葉生態區,不育細胞質對農藝性狀的影響各不相同:在西昌,tab1-CMS細胞質對所調查的6個農藝性狀均無顯著的細胞質效應;在諸城,tab1-CMS細胞質對62.9%(17/27)的性狀有顯著影響,有些表現為正效應,有些表現為負效應。以云煙87和G13-5為材料配制正反交組合F1代農藝性狀分析顯示,株高、有效葉數、腰葉長正反交存在顯著差異,而節距、莖圍、腰葉寬不存在正反交差異[31],說明細胞質在一定程度上會影響煙草F1代的農藝性狀。
環境是影響農藝性狀的重要因素,本研究中的諸城和西昌兩個煙草生態區海拔、緯度不同,環境因素對所調查的6個農藝性狀均有顯著或極顯著影響。其他研究者的研究也表明,環境因素對不同品種的煙草農藝性狀有一定的影響[32]。核基因組(品種)是影響農藝性狀的主要因素,在本研究中,核基因組對莖圍和最大葉葉長等部分農藝性狀有顯著影響,對株高、最大葉葉寬無顯著影響。黃鵬等[33]對畢納1號等7個煙草品種的農藝性狀比較也顯示,在株高等農藝性狀上不同品種間的表現不同,暗示品種間的農藝性狀差異與品種間的親緣關系有一定的關系。細胞質-環境、細胞質-細胞核、細胞質-細胞核-環境之間存在相互作用,互作對節距等部分性狀有顯著影響。總的來說,在配置雜交組合和不?育系轉育時,需要考慮父本的核背景以及品種的種植區域等因素。
在本研究中,tab1-CMS細胞質在黑脛病、CMV抗性上表現為正效應,在PVY和TMV抗性上與可育胞質無顯著差別。煙草其他不育胞質在抗病性上也觀察到顯著的胞質效應,孫玉合等[34]的研究表明,不育胞質來源于N. glauca的不育系86-6與其保持系革新一號相比,在黑脛病、赤星病的抗性上好于革新一號,而不育胞質來源于N. bigelovii、N. megalosiphon和 N. plumbaginifolia的不育系抗病性減弱[2]。因此,在煙草抗病育種中,除了發掘優異的抗病種質外,還需要考慮細胞質對抗病性的影響。
高等植物葉綠體基因組在結構和組成上高度保守,但在葉綠體基因組的非編碼區(如內含子或基因間區)存在大量的SNP、InDel和基因組結構變異[35-36],葉綠體中這些快速進化的序列片段為快速準確地識別物種提供了大量的分子標記信息[37-38]。本研究中,通過比較tab1-CMS,sua-CMS和可育煙草中trnC-trnD基因間區域的序列,獲得了25個變異位點。變異頻率為0.7%,顯示了在trnC-trnD的基因間區域變異豐富。
4? 結? 論
本研究通過比較tab1-CMS不育系YK01和同核異質系K326與4個烤煙品種雜交F1代在農藝性狀和抗病性上的差異,認為tab1-CMS細胞質對后代的農藝性狀和抗病性上沒有顯著的負效應,有的性狀還表現為正效應,可作為煙草育種中的優異不育胞質來源。但是煙草農藝性狀和抗病性的影響因素很多且存在著復雜的相互作用,尚需更多研究明確tab1-CMS細胞質在煙草遺傳改良中的利用規律,以對其胞質效應進行更全面的評價。
參考文獻
[1] KUMAR B V, VENUGOPAL M, BARWALE Z U, et al. Heterosis: emerging ideas about hybrid vigour[J]. Journal of Experimental Botany, 2012, 63(18): 6309-6314.
[2] 佟道儒. 煙草育種學[M]. 北京:中國農業出版社,1997:279-294.
TONG D R. Tobacco breeding[M]. Beijing: China Agricultural Press, 1997: 279-294.
[3] GRACEN, GROGAN. Diversity and Suitability for Hybrid Production of Different Sources of Cytoplasmic Male Sterility in Maize[J]. Agronomy Journal, 1974, 66(5): 654-657.
[4] TRAVEN A, WONG J M S, XU D, et al. Interorganellar Communication[J]. Journal of Biological Chemistry, 2001, 276(6): 4020-4027.
[5] QIN P, WANG Y, LI Y Y, et al. Analysis of cytoplasmic effects and fine-Mapping of a genic male sterile line in rice[J]. Plos One, 2013, 8(4): e61719.
[6] YAO S, CHEN T, ZHANG Y D, et al. Transferring translucent endosperm mutant gene wx-mq and rice stripe disease resistance gene stv-bi by marker-assisted selection in rice (Oryza sativa)[J]. Rice Science. 2011, 18(2): 102-109.
[7] 王學德. 棉花細胞質雄性不育的研究與利用[J]. 中國農業科學,2019,52(8):1341-1354.
WANG X D. Overview of the study and application of cytoplasmic male sterility in cotton[J]. Scientia Agricultura Sinica, 2019, 52(8): 1341-1354.
[8] 黃得潤,沈君輝,劉光杰,等. 作物抗病蟲性的細胞質遺傳效應研究進展[J]. 中國農學通報,2006(9):333-337.
HUANG D R, SHEN J H, LIU G J, et al. Recent advances in inheritance of cytoplasmic effects on crop resistance to diseases and insect pests[J]. Chinese Agricultural Science Bulletin, 2006(9): 333- 337.
[9] 馬文廣,張恒,白永富,等. 烤煙雄性不育系與其同型可育品種性狀比較[J]. 湖南農業大學學報(自然科學版),2005(5):38-41.
MA W G, ZHANG H, BAI Y F, et al. Comparison of traits of the mail sterile lines and their maintainer lines of flue-cured tobacco[J]. Journal of Hunan Agricultural University (Natural Sciences), 2005(5): 38-41.
[10] KIM S, PLAGNOL V, HU T T, et al. Recombination and linkage disequilibrium in Arabidopsis thaliana[J]. Nature Genetics, 2007, 39(9): 1151.
[11] YU X L, LIU Y P, LV Y X, et al. Development of molecular markers specific to petaloid-type cytoplasmic male sterility in tuber mustard (Brassica juncea var. tumida Tsen et Lee)[J]. Mol.Biol. Rep, 2014, 41(2): 769-778.
[12] TAN Y P, XU X, WANG C T, et al. Molecular characterization and application of a novel cytoplasmic male sterility-associated mitochondrial sequence in rice. BMC Genet 16, 45 (2015). https://doi.org/10.1186/s12863-015-0205-0.
[13] HENG S, LIU S S, XIA C X, et al. Morphological and genetic characterization of a new cytoplasmic male sterility system (oxa CMS) in stem mustard (Brassica juncea)[J]. Theoretical and applied genetics. Theoretische und angewandte Genetik, 2018, 131(1): 59-66.
[14] HENNG S P, SHI D Y, HU Z H, et al. Characterization and classification of one new cytoplasmic male sterility (CMS) line based on morphological, cytological and molecular markers in non-heading [J]. Chinese cabbage (Brassica rapa L.), 2015, 34(9): 1529-1537.
[15] SHU J, LIU Y, LI Z, et al. Detection of the diversity of cytoplasmic male sterility sources in Broccoli (Brassica Oleracea var. Italica) using mitochondrial markers[J]. Frontiers in Plant Science, 2016, 7: 927-927.
[16] LI N, ZHENG Y Q, DING H M, et al. Development and validation of SSR markers based on transcriptome sequencing of Casuarina equisetifolia[J]. Trees, 2018, 32(1): 41-49.
[17] 李鳳霞,楊愛國,崔萌萌,等. 四種細胞質來源的煙草不育系線粒體SSR位點差異[J]. 作物學報,2011,37(12):2285-2285.
LI F X, YANG A G, CUI M M, et al. Mitochondrial microsatellite variability of tobacco cms lines with four different cytoplasms[J]. Acta Agronomica Sinica, 2011, 37(12): 2285-2285.
[18] 孫炳蕊,范芝蘭,李晨,等. 含不同地區野生稻胞質的水稻雄性不育材料線粒體SSR標記分析[J]. 分子植物育種,2016,14(9):2392-2404.
SUN B R, FAN Z L, LI C, et al. The analysis of rice cytoplasmic male sterility lines with different wild rice cytoplasm using the mitochondrial SSR markers[J]. Molecular Plant Breeding, 2016, 14 (9): 2392-2404.
[19] 袁凱,張婷,史曉芳,等. K、V、T型小麥細胞質雄性不育系葉綠體DNA的SSR分析及RuBP羧化酶活性比較[J]. 西北植物學報,2019,39(3):97-104.
YUAN K, ZHANG T, SHI X F, et al. Comparison of chloroplast DNA and RuBP carboxylase (rubisco) activity with K, V, T-type cytoplasmic male-sterile wheat lines[J]. Acta Botanica Boreali- Occidentalia Sinica, 2019, 39(3): 97-104.
[20] 劉光照. 辣椒細胞質雄性不育系細胞學觀察和mtDNA-AFLP分析[D]. 咸陽市:西北農林科技大學,2010.
LIU G Z. Cytological observation of pepper CMS and mtDNA-AFLP analysis[D]. Xianyang: Northwest A & F University, 2010.
[21] CHRISTOPHER V, AGNIESZKA K, MICHAEL H. Sequencing and annotation of the chloroplast DNAs and identification of polymorphisms distinguishing normal male-fertile and male-sterile cytoplasms of onion[J]. Genome, 2013, 56(12): 737-742.
[22] LIN H, HONG X, ZHOU P, et al. Combination of microRNA expression profiling with genome-wide SNP genotyping to construct a coronary artery disease-related miRNA-miRNA synergistic network[J]. Bioscience trends, 2014, 8(6): 297-307.
[23] DANIELL H, LIN C S, YU M, et al. Chloroplast genomes: diversity, evolution, and applications in genetic engineering[J]. Genome Biology, 2016, 17(1): 1-29.
[24] LIN D, GONG X, JIANG Q, et al. The rice ALS3 encoding a novel pentatricopeptide repeat protein is required for chloroplast development and seedling growth[J]. Rice, 2015, 8(1): 17.
[25] KOHN C V, KIELKOWSKA A, HAVEY M J. Sequencing and annotation of the chloroplast DNAs and identification of polymorphisms distinguishing normal male-fertile and male-sterile cytoplasms of onion[J]. Genome 2013, 56: 737-742.
[26] LEE C, WEN J. Phylogeny of Panax using chloroplast trnC-trnD intergenic region and the utility of trnC-trnD in interspecific studies of plants[J]. Molecular Phylogenetics & Evolution, 2004, 31(3): 894-903.
[27] FRITSCH, PETER W, CRUZ, et al. Phylogeny of Symplocos Based on DNA Sequences of the Chloroplast trnC-trnD Intergenic Region[J]. Systematic Botany, 2006. 31(1): 181-192.
[28] TALEBI, MAJID, AKBARI, et al. Molecular polymorphism in Pistacia vera L. using non-coding regions of chloroplast DNA[J]. Genetic Engineering and Biotechnology Journal, 2016, 14(1): 31-37.
[29] 國家煙草專賣局. 煙草農藝性狀調查方法:YC/T 142—2010[S]. 北京:中國標準出版社,2003:227-231.
State Tobacco Monopoly Administration. Investigation method of tobacco agronomic traits: YC/T 142—2010[S]. Beijing: China Standards Press, 2003: 227-231.
[30] 張維軍,袁漢民,王小亮,等. 寧夏春小麥抗旱性的遺傳多樣性分析[J]. 干旱地區農業研究,2017,9(6):95-103.
ZHANG W J, YUAN H M, WANG X L, et al. Genetic diversity of spring wheat germplasm resources based on drought resistance in Ningxia[J]. Agricultural Research in the Arid Areas, 2017,9(6):95-103.
[31] QIN P, WANG Y, LI Y, et al. Analysis of cytoplasmic effects and fine-mapping of a genic male sterile line in rice[J]. PLoS One, 2013, 8: e61719.
[32] 宋鵬飛,陳興位,馬迅,等. 緯度和海拔二維因素對云南烤煙農藝性狀的影響[J]. 西南農業學報,2017,30(10):2345-2351.
SONG P F, CHEN X W, MA X, et al. Effects of latitude and altitude on agronomic traits of flue-cured tobacco in Yunnan[J]. Southwest China Journal of Agricultural Sciences, 2017, 30(10): 2345-2351.
[33] 黃鵬,徐慶國. 不同烤煙品種農藝性狀與經濟性狀的差異研究[J]. 作物研究,2016,2(2):126-130.
HUANG P, XU Q G. The Differences of agronomic and economic characters in different flue-cured tobacco cultivars[J]. Crop Research, 2016, 2(2): 126-130.
[34] 孫玉合,丁昌敏,張歷歷,等. 煙草新胞質雄性不育系86-6的創造及其利用[J]. 中國煙草學報,1999(1):3-5.
SUN Y H, DING C M, ZHANG L L, et al. Creation and utilization of new cytoplasmic male sterile line 86-6 in tobacco[J]. Acta Tabacaria Sinica, 1999(1): 3-5.
[35] NIELL H, LIN C S, YU M, et al. Chloroplast genomes: diversity, evolution, and applications in genetic engineering[J]. Genome Biology, 2016, 17(1): 1-29.
[36] BAO H, XIONG Y, GUO H, et al. MapNext: a software tool for spliced and unspliced alignments and SNP detection of short sequence reads[J]. BMC Genomics, 2009, 10(16): 1851-1858.
[37] JANSEN RK, CAI Z, RAUBESON LA, et al. Analysis of 81 genes from 64 plastid genomes resolves relationships in angiosperms and identifies genome-scale evolutionary patterns[J]. Proc Natl Acad Sci, 2007, 104: 19369-19374.
[38] HAN Y W, DUAN D, MA X F, et al. Efficient identification of the forest tree species in Aceraceae using DNA barcodes[J]. Front Plant Sci, 2016, 7: 1707.
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
