禾花鯉攝食選擇與仔稚魚消化器官發育的研究
2021-11-23 13:52:22羅雨蕭蔣士宋梁藝馨龐琳星莫翠琴
水產科學 2021年6期
劉 婷,黃 凱,林 勇,羅雨蕭,蔣士宋,梁藝馨,龐琳星,莫翠琴
( 1.廣西大學 動物科學技術學院,廣西 南寧 530004; 2.廣西水產科學研究院,廣西 南寧 530021; 3.全州縣科學技術情報研究所,廣西 桂林 541500 )
禾花鯉(Cyprinuscarpio),屬鯉科溫水性魚類,原產于桂林市全州縣龍水鄉和融水縣苗山等地,是具有地方特色的土著養殖魚類,因主食稻田禾花并帶有特色風味而得名。禾花鯉食性廣,生長速度快,繁殖能力強,肉質鮮美,魚骨柔軟無泥腥味。有研究表明,禾花鯉具有4種呈味氨基酸,營養價值較高且礦物質含量豐富,證實其是適宜推廣養殖和加工利用的魚類品種[1-2]。隨著稻田種養模式產業化和規模化發展,禾花鯉已成為華南地區稻田養殖對象研究的熱點[3-4]。與普通鯉魚相比,禾花鯉上市規格較小,通常規格為30~50 g/尾,稻田中養殖的禾花鯉旺季價格可達30元/kg。然而,市場面臨著需求量大的問題,且在禾花鯉仔稚魚階段尚無較為理想的人工開口餌料。
消化器官的發育影響魚類對攝取食物的消化和吸收程度,且與魚類的生長發育有關。國內外關于魚類消化器官發育的組織學研究已有諸多報道,包括魚(Lizahaematocheila)[5]、泥鰍(Misgurnusanguillicaudatus)[6]、日本白姑魚(Argyrosomusjaponicus)[7]等,但對禾花鯉仔魚到稚魚消化器官發育的組織學變化研究尚未見報道。在早期生長發育過程中,隨著消化器官的逐步完善,消化酶活性也隨之改變。幼魚早期不同發育階段消化酶活性變化已經作為幼魚發育程度、成活率和人工飼料投喂初始時間的參考依據[8]。因此,筆者測定禾花鯉早期生長、攝食選擇與消化酶活性變化,并采用組織切片技術對消化器官組織結構進行觀察,了解飼養階段禾花鯉的食性和消化生理特性,為開發出適宜的禾花鯉仔稚魚開口飼料提供理論基礎和參考。
1 材料與方法
1.1 試驗材料
試驗魚為2018年5月2日于廣西農業良種海南南繁育種基地進行人工催產孵化獲得,親魚產卵池規格4.5 m×1.8 m×0.8 m,于200 m2的水池中孵化,孵化水溫為29 ℃,經36~48 h后魚苗孵出,自孵化后第2~10天向池內潑灑熟豆漿,豆漿投喂量為4 kg/666.7 m2,日投喂2次(9:00、16:00),自第11天起每隔2 d使用水蛛樂和益菌寶(廣東佛山市順德區活寶源生物科技有限公司)追肥1次,養殖周期為30 d。
1.2 試驗方法
1.2.1 口徑與生長測量
1.2.2 育苗池浮游生物采集定量
在育苗池的不同采樣點采集浮游植物:使用采水器采水1 L,倒入水樣瓶用15 mL魯哥氏液固定,沉淀后去掉上清液,濃縮至20~40 mL沉淀物保存于棕色廣口瓶內。浮游動物的采集:用采水器采水20 L,經浮游生物網過濾濃縮于100 mL廣口瓶中,加入5 mL福爾馬林固定。將沉淀物滴滿計數框置于顯微鏡下進行計數,方法參考《水生生物學》[10]。
1.2.3 消化道內含物采集與測定
由于仔魚較小,無法分離出腸道內含物,因此分別在孵化后第14、18、22、26、30天取20尾魚腸道,將腸道置于10%福爾馬林中固定,在體視解剖鏡(XTL-165-VT/MT,鳳凰)下解剖取出魚腸內含物,吸去水分后,分離食物團,置于顯微鏡(MODEL E100,尼康)下觀察。通過計算每份食物體積占腸內含物總體積的百分比,或每類食物出現的頻率來評估魚對食物的選擇[11]。計算仔稚魚對養殖環境中餌料生物的選擇性(E):
E=(r-p)/(r+p)
式中,r為某種餌料在腸道食物中所占的比例,p為某種餌料在育苗池中所占的比例。E值為-1.0 (完全不選食)~1.0(總是選食)[12]。
1.2.4 消化器官的組織切片觀察
組織切片的制作參考文獻[13]的方法,對出膜1~30 d的魚消化器官(包括口咽腔、食道、腸道、肝胰臟)進行組織結構觀察。隔日每次取樣20尾。采用10%福爾馬林固定,石蠟包埋,使用德國徠卡切片機(RM 2126)進行縱、橫方向連續切片,厚度5 μm,蘇木精—伊紅染色,中性樹膠封片,在尼康顯微鏡下觀察拍照。
1.2.5 消化酶活性測定
取樣前將魚置于清水中空腹過夜,前10 d隔天取樣1次,后10 d每隔4 d取樣1次,1~15 d的仔魚取全魚,16~20 d的仔稚魚剪取腹部進行消化酶活性測定。每次取樣2.5 g,每組3個平行樣。其中的2.0 g樣品在10倍體積冰凍雙蒸水里進行組織勻漿,于-80 ℃冰箱冷凍保存,使用南京建成生物工程研究所的酶試劑盒測定酸性蛋白酶、堿性蛋白酶、淀粉酶和堿性磷酸酶的活性。使用生工生物工程(上海)股份有限公司的改良型BCA蛋白濃度試劑盒測定酶蛋白濃度。0.5 g樣品保存于-80 ℃冰箱,使用索萊寶的酶試劑盒測定脂肪酶活性。
1.3 數據處理
數據使用Excel軟件進行統計,采用軟件SPSS 21.0進行單因素方差分析,所得數據用平均值±標準差表示。
2 結 果
2.1 禾花鯉仔稚魚口徑的變化與仔魚生長形態
2.1.1 禾花鯉仔稚魚生長與口徑變化
魚孵出后第3天測量到口徑大小,3 d后口徑隨著日齡的增加而變化,口徑長(y,mm)與日齡(x,d)的關系式為:y=0.0033x2+0.0421x+0.3774,r2=0.9754(圖1a)。在前10 d生長速度較為緩慢,從12 d開始生長速度明顯加快,體長和全長均隨著日齡的增加而增長(圖1b)。體長(y,mm)與日齡(x,d)的關系式為:y=0.1076x2-0.75x+6.6221,r2=0.9874;全長(y,mm)與日齡(x,d)的關系式為:y=0.1399x2-0.904x+7.3907,r2=0.9884。
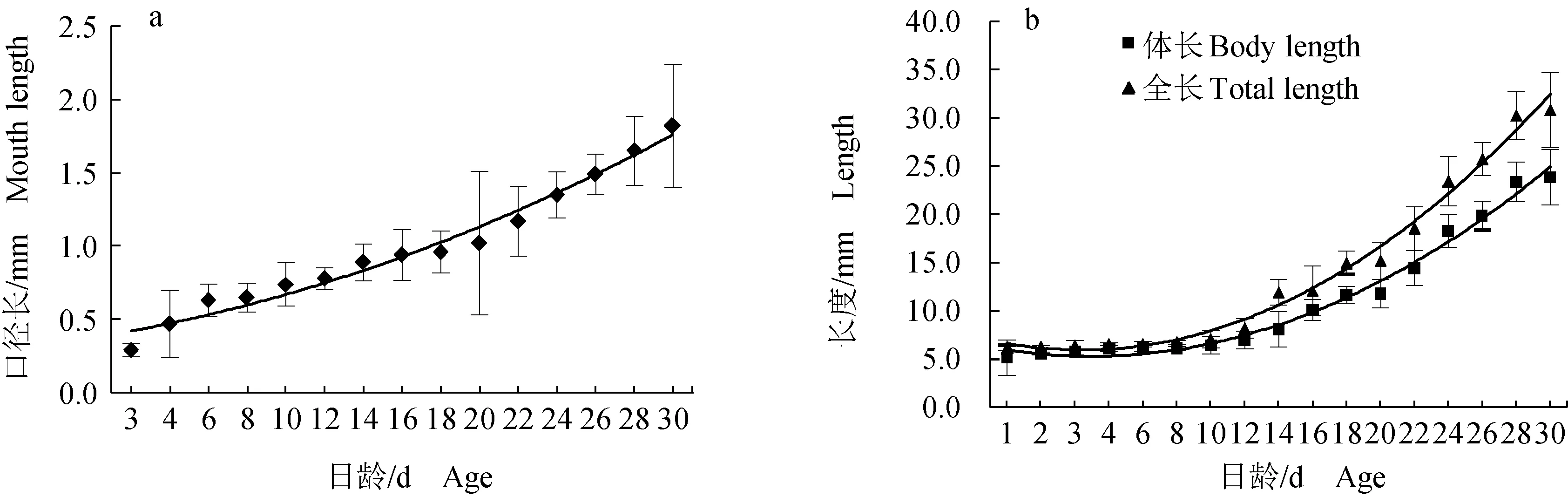
圖1 禾花鯉仔稚魚口徑長(a)與體長、全長(b)變化Fig.1 Changes in mouth length (a), body length, and total length (b) of rice flower carp P. merus larvae and juveniles
2.1.2 禾花鯉仔魚的形態變化
1 d仔魚在孵化初期尚未開口,營養主要來源于卵黃囊(圖2a);2 d仔魚卵黃囊尚未完全消失,但已出現口裂、鰓弓(圖2b);發育至第3 d,仔魚開口攝食(圖2c)。10 d首次觀察到鱗片,鱗片的出現標志著仔魚期向稚魚期的轉變;生長至17 d,所有鱗片基本長齊,為稚魚過渡到幼魚的階段。
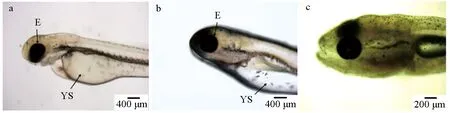
圖2 禾花鯉仔魚的形態特征Fig.2 Morphological characteristics of rice flower carp P. merus larvaea.1 d仔魚; b.2 d仔魚; c.3 d仔魚; YS.卵黃; E.眼.a.1 day old larva; b.2 day old larva; c.3 day old larva; YS.yolk sac; E.eye.
2.2 禾花鯉育苗池的浮游生物組成
14~30 d育苗池中浮游動物的組成以輪蟲數量最多,隨著養殖天數的增加,輪蟲數量逐漸上升,但在26 d明顯減少,30 d又增加(圖3a)。枝角類和橈足類的數量呈現規律性上下浮動。浮游植物組成以藍藻門和硅藻門為主,在22 d后藍藻門數量逐漸降低(圖3b)。硅藻門數量呈現先降低22 d后逐漸增加的趨勢。綠藻門自18 d開始出現,26 d開始可觀察到甲藻門和黃藻門的少量個體。
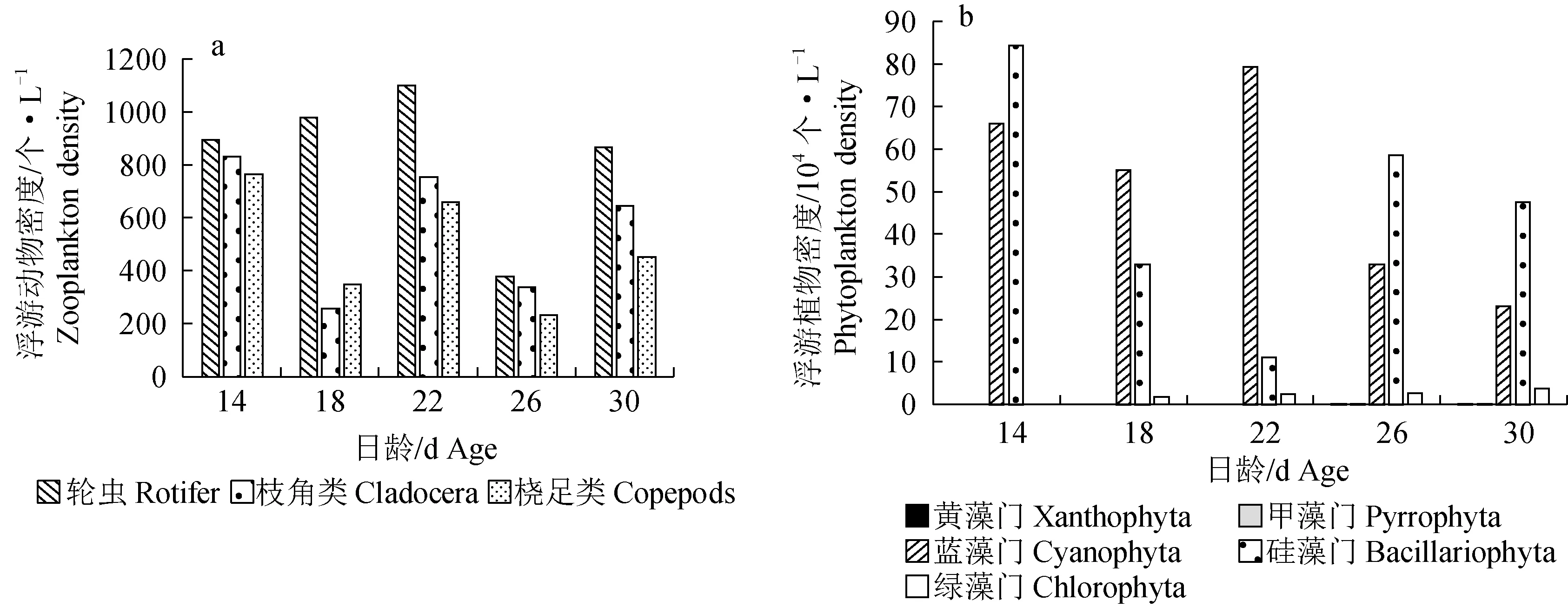
圖3 禾花鯉育苗池浮游生物餌料組成Fig.3 Composition of plankton food in the seedling pond of rice flower carp P. merus
2.3 禾花鯉稚魚腸道內含物的餌料組成
14~30 d稚魚腸道內含物的浮游植物組成主要以藍藻門和硅藻門為主,綠藻門、甲藻門和黃藻門較少或幾乎觀察不到;22 d前,藍藻門和硅藻門共約占當天總體組成比例的40%~60%,22 d后藍藻門和硅藻門的比例有所下降(圖4)。枝角類和橈足類組成比例隨著日齡增加逐漸提高,22 d后,腸道內含物中生物總體組成轉變為以枝角類和橈足類為主,共約占當天總體組成的40%~50%;輪蟲數量逐漸減少,26 d未觀察到輪蟲,30 d又觀察到少量輪蟲,約占1%。在22 d首次觀察到搖蚊幼蟲,且比例隨日齡逐漸增大。腸道中部分已消化成糊狀的食物占比相對較大,歸類為其他。
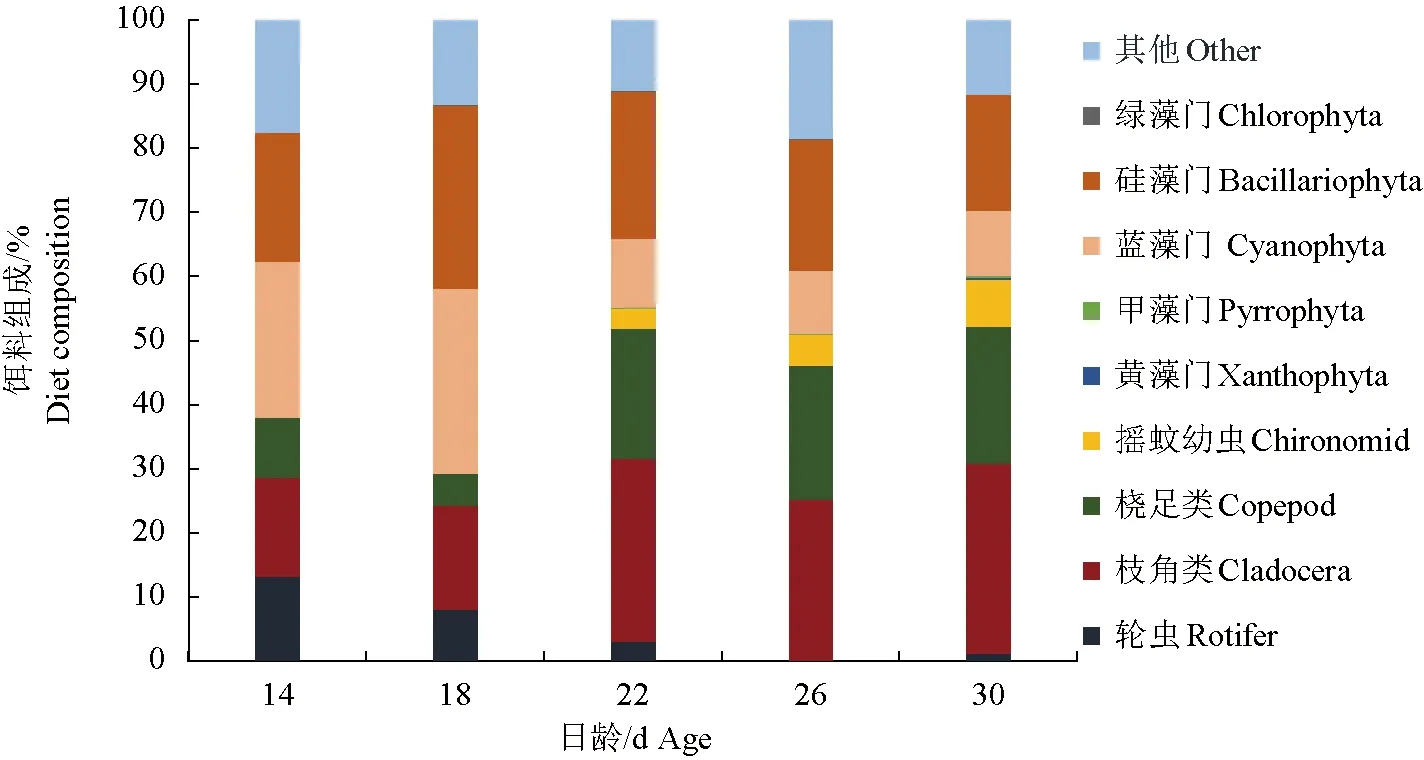
圖4 不同日齡的禾花鯉腸道內含物組成Fig.4 Food item composition of intestinal contents of rice flower carp P. merus with different daily age
2.4 禾花鯉稚魚的攝食選擇性指數
14~30 d稚魚對枝角類和橈足類的偏好性強,攝食選擇性指數值均接近1,成為其重要的食物來源(圖5)。對于藍藻門、綠藻門的偏好性極差,攝食選擇性指數為0或負數,不會對其主動選食,為被動攝食。甲藻門攝食選擇性指數時正時負,攝食選擇不穩定。在22 d,禾花鯉對黃藻門、甲藻門和硅藻門有明顯的選擇性,攝食選擇性指數為正。在26 d前,對輪蟲的偏好性強,攝食選擇性指數在0.9以上;在26 d,對輪蟲的攝食選擇性指數為-1,完全回避攝食,26 d后逐漸恢復攝食。
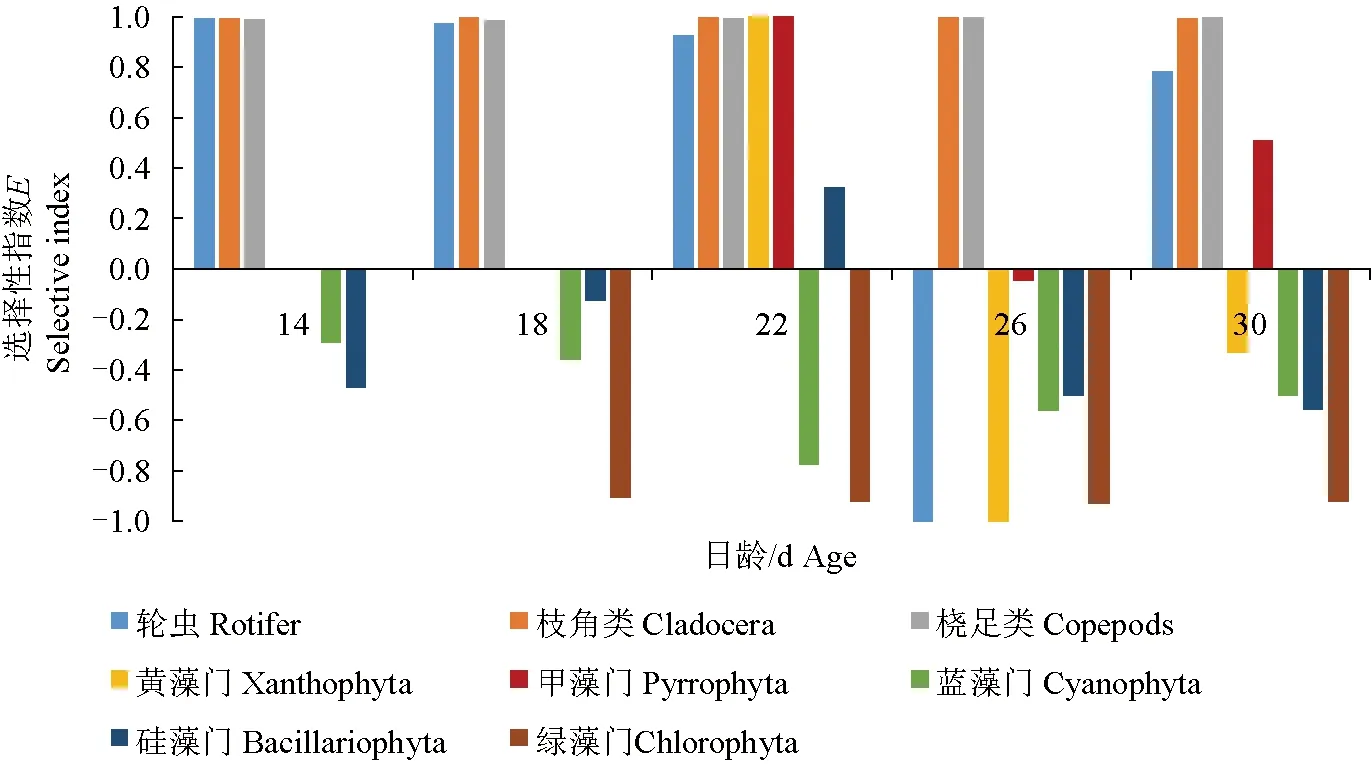
圖5 禾花鯉對不同餌料的選擇性指數Fig.5 The selectivity index E of food items in rice flower carp P. merus
2.5 禾花鯉仔稚魚消化器官的組織結構觀察
2.5.1 口咽腔
第3天仔魚開口,口腔表面的黏膜層發育出復層扁平上皮及少量杯狀細胞,但未發現味蕾,黏膜層較薄,咽部出現角質墊(圖6a)。發育至第5天,黏膜層加厚,可見黏膜下層、肌肉層、漿膜層,杯狀細胞逐漸增多,味蕾開始出現,黏膜上皮由直線型變為“凹”型,隨后逐漸發育出齒囊(圖6b、c)。隨著日齡增加,黏膜層逐漸加厚,黏液細胞增多(圖6d);第21天,口咽腔已基本發育完全(圖6e)。
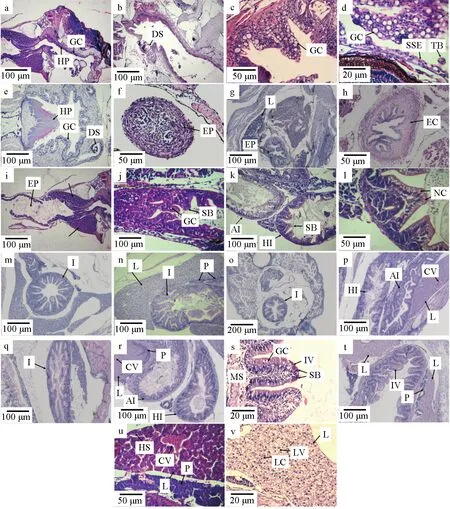
圖6 禾花鯉仔稚魚消化器官的組織學變化Fig.6 Histologic changes in digestive organs of rice flower carp P. merus larvae and juvenilea.3 d口咽腔縱切; b.6 d口咽腔縱切; c.13 d口咽腔橫切; d.13 d口咽腔縱切; e.21 d口咽腔橫切; f.8 d食道橫切; g.19 d食道橫切; h.30 d食道橫切; i.3 d腸道縱切; j.6 d腸道縱切; k.12 d腸道縱切; l.12 d腸道縱切; m.15 d 腸道橫切; n.15 d腸道及肝胰臟縱切; o.17 d腸道及肝胰臟橫切; p.17 d腸道縱切; q.19 d 腸道縱切; r.23 d腸道及肝胰臟縱切; s.29 d腸道橫切; t.30 d腸道及肝胰臟橫切; u.8 d肝胰臟縱切; v.29 d肝胰臟橫切. HP.角質墊; GC.杯狀細胞; DS.齒囊; SSE.復層扁平上皮; TB.味蕾; EP.食道; EC.食物團; L.肝臟; SB.紋狀緣; I.腸; AI.前腸; HI.后腸; NC.核上內容物; MS.肌肉層; CV.中央靜脈; P.胰臟; IV.腸絨毛; HS.肝血竇; LC.肝臟細胞; LV.肝臟空泡.a.longitudinal section of oropharyngeal cavity in 3 d; b.longitudinal section of oropharyngeal cavity in 6 d; c.cross section of oropharyngeal cavity in 13 d; d.longitudinal section of oropharyngeal cavity in 13 d; e.cross section of oropharyngeal cavity in 21 d; f.cross section of esophagus in 8 d; g.cross section of esophagus in 19 d; h.cross section of esophagus in 30 d; i.longitudinal section of intestine in 3 d; j.longitudinal section of intestine in 6 d; k.longitudinal section of intestine in 12 d; l.longitudinal section of intestine in 12 d; m.cross section of intestine in 15 d; n.longitudinal section of intestine and hepatopancreas in 15 d; o.cross section of intestine and liver in 17 d; p.longitudinal section of intestine in 17 d; q.longitudinal section of intestine in 19 d; r.longitudinal section of intestine and hepatopancreas in 23 d; s.cross section of hepatopancreas in 29 d; t.cross section of intestine and hepatopancreas in 30 d; u.longitudinal section of hepatopancreas in 8 d; v.cross section of liver in 29 d. HP.horny pad; GC.goblet cell; DS.dental sac; SSE.stratified squamous epithelium; TB.taste buds; EP.esophagus; EC.esophageal contents; L.liver; SB.striated border; I.intestine; AI.forgut; HI.hindgut; NC.supran uclear content; MS.muscle layer; CV.central vein; P.pancreas; IV.intestinal villus; HS.hepatic sinusoid; LC.liver cell; LV.liver vacuole.
2.5.2 食道
食管初始為半封閉狀態,食道短且狹窄,由復層鱗狀上皮構成黏膜層,發育至第8天,食道仍未完全貫通(圖6f),后期逐漸出現褶皺及杯狀細胞,黏膜層出現橫紋肌纖維。發育至第19天食道可見大量杯狀細胞,橫紋肌纖維逐漸增厚,但未見黏膜下層及肌層(圖6g),至第30天肌層相繼出現(圖6h)。
2.5.3 腸道
仔魚在剛孵出時消化管位于卵黃囊與脊索之間,呈直管狀,出膜后第2天卵黃囊變小(圖2)。第3天逐漸消失,仔魚開口后進入混合營養期,腸腔內可見食物團,出現褶皺及單層柱狀細胞,但褶皺不明顯(圖6i)。第6天出現較薄的環肌,首次觀察到杯狀細胞和紋狀緣(圖6j)。隨著日齡增加,腸腔褶皺越來越明顯,高度不斷增加,杯狀細胞逐漸增多,發育至第12天,出現前腸和后腸,最先出現核上內容物的是前腸(圖6k、l)。后期腸道不斷增加,形成復雜的彎曲形狀(圖6p、r),發育至第30天,紋狀緣已經非常明顯,杯狀細胞豐富,腸道的形態發育和細胞分化基本完善(圖6s)。
2.5.4 肝胰臟
在仔魚生長至第3天時,可見腸道兩側出現疑似肝臟細胞團,肝細胞形態不明顯,細胞核顏色較深(圖6i)。隨后體積不斷增大,發育至第8天可觀察到明顯的中央靜脈及肝細胞索,隨著營養積累,肝臟開始出現空泡(圖6u),第29天肝臟空泡增多(圖6v)。胰臟在第8天觀察到,且分布較復雜散亂,呈彌散型(圖6n,r,t,u)。
2.6 禾花鯉仔稚魚消化酶活性變化
仔魚開口前就具有消化酶活性(圖7)。8~10 d脂肪酶活性升至最大值,接著出現下降,14~26 d基本穩定,26 d后急劇增加(圖7a)。淀粉酶活性從開口后第4 d逐漸下降,最低值出現在第8天,隨后逐漸上升再下降,直到第30天達到最大值(圖7b)。堿性蛋白酶活性在開口后有明顯上升;酸性蛋白酶在第14天和第26天出現峰值;且酸性蛋白酶和堿性蛋白酶的活性變化呈互補關系,當酸性蛋白酶活性升高時,堿性蛋白酶活性降低,反之亦然(圖7c)。第4天堿性磷酸酶活性出現峰值;第8天出現最高值,活性為0.21(圖7d)。
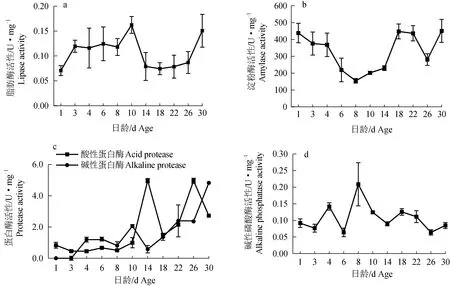
圖7 禾花鯉仔稚魚消化酶活性變化Fig.7 Changes in digestive enzyme activities of rice flower carp P. merus larvae and juvenile
3 討 論
3.1 禾花鯉的生長與稚魚后期攝食選擇
第3天仔魚即開口攝食,混合營養期持續不到1 d。混合營養階段是仔魚培育過程的關鍵時期,其組織形態和生理機能都將發生改變[14],此時需要給予仔魚營養豐富的適口餌料以提高其成活率。第10天部分禾花鯉已經長出鱗片,且10 d前出現生長緩慢的情況。有研究表明,在鱗片出現前后魚全長的日增長會下降,是因為用于生長的部分能量被用于了鱗片形成[15]。
仔魚口徑長短與攝食的餌料規格密切相關。有研究報道,鯉魚開口攝食的體長為6.6 mm,口徑為0.57 mm,能攝食285~425 μm的餌料,不能攝食水蚤和裸腹溞(Moina)成體,但可將其幼體作為攝食對象[9]。禾花鯉開口攝食的體長和口徑長相對較小,分別為5.74、0.29 mm,仔魚從開口第3天至第30天,口徑增大逾6倍,為1.82 mm。參考李云蘭等[16]的方法(飼料粒徑=口徑長×75%×25%),計算得到禾花鯉的適宜飼料粒徑,孵化后3~14 d為50~170 μm,14~30 d為170~350 μm。推測14 d前可能主要以小型輪蟲幼體(40~150 μm)和小型藻類(<100 μm)為食,隨著口徑長增大而主動選擇攝食體型更大的枝角類和橈足類,14 d后對浮游動物的攝食量也逐漸加大。至第22天時,首次從禾花鯉稚魚體內觀察到搖蚊幼蟲,搖蚊幼蟲屬于底棲生物,體現禾花鯉雜食的特性。
禾花鯉攝食選擇與育苗池中浮游生物組成有一定關聯。在14~30 d,育苗池中浮游植物主要以藍藻門和硅藻門為主,優勢種分別為微囊藻和直鏈藻,但攝食選擇性結果顯示,禾花鯉對其無明顯的主動選擇性,這可能與養殖環境中它們占主要比例有關,導致禾花鯉被動選擇食物。藍藻門中藻類大多含有微囊藻毒素,魚類攝食大量藍藻會對其組織發育造成損傷。但有研究表明,鳙魚(Aristichthysnobilis)可長期耐受高含量的微囊藻毒素[17]。而禾花鯉屬于被動攝食藍藻,是否對藍藻有很強的消化能力和耐受力,還需深入研究。育苗池中藍藻數量較多,表明應及時調整水質環境,以達到提高苗種成活率的目的。
26 d前育苗池中浮游動物以輪蟲數量最多,稚魚開始主動選擇攝食輪蟲。26~30 d禾花鯉對輪蟲攝食出現從無到恢復攝食的情況。推測一方面與育苗池中輪蟲數量出現先減后增有關,另一方面與消化器官發育完善而需攝食高營養的動物餌料有關。因此,在14~30 d,禾花鯉對浮游動物的攝食量逐漸升高,且主要以大型蛋白動物性餌料為食,說明此時需要攝入大量營養(蛋白質和脂肪)才能滿足稚魚的生長發育,應給予營養均衡的配合飼料并混合浮游動物投喂。其他藻類數量較少,攝食量也少。
3.2 禾花鯉消化器官發育與消化酶變化
仔魚開口后消化酶活性會受到消化器官生長發育的影響。3 d后仔魚各部分消化器官已開始分化,第6天首次從腸道內觀察到杯狀細胞,腸道中的杯狀細胞可分泌黏液和消化酶,促進食物的消化吸收。此時也出現紋狀緣,可增大腸道吸收表面積,它是魚類幼體發育過程中獲得成體消化模式的重要標志和關鍵步驟[18]。第8天堿性磷酸酶出現最大值,說明腸道功能發育完善[19]。第4天堿性磷酸酶活性出現峰值,可能受仔魚開口攝食的影響和與腸道功能開始發育有關。8 d后淀粉酶生產的中心器官肝胰臟逐漸發育完善,酶活性隨之增強。脂肪酶活性在8~10 d突然增加達到最大值,可能與第8天肝臟逐漸發育出中央靜脈和肝細胞索,并開始貯存糖原進行營養積累有關;后期脂肪酶活性下降可能與仔魚組織蛋白的增加有關[20]。本試驗結果顯示,禾花鯉仔魚在開口前已具有消化酶活性,與劉亞秋等[21-22]研究結果一致,其認為仔魚開口前消化酶活性受基因決定。
3.3 禾花鯉消化器官發育與攝食消化
3 d仔魚開口攝食,口腔內出現少量杯狀細胞,對堅硬食物起到潤滑作用。5 d觀察到咽齒齒囊,6~10 d長出咽齒,禾花鯉的咀嚼能力得到加強。10 d前,仔魚食道未完全打開,無法完全適應外源性營養攝入,生長較緩慢。根據討論3.1節可知,前期主要以微小的藻類和輪蟲幼體為食。因此,在育苗池培育小型浮游動物的同時也要培育一定量的浮游植物,為仔魚提供豐富的食物來源。10 d后腸道功能逐漸加強,發育至第12天出現了核上內容物和空泡結構,說明腸道細胞已可進行胞飲作用和細胞內消化吸收蛋白質大分子[18]。第14天和第26天酸性蛋白酶出現峰值,預示著腸道消化蛋白機能的逐漸完善,14 d后食性轉變為動物性,生長速度明顯加快。因此,稚魚期的禾花鯉更傾向于攝食體格較大的浮游動物(枝角類和橈足類) 來為機體提供能量,進而導致第26天時對輪蟲偏好性降低,攝食由主動變為被動。30 d消化器官基本發育完全,更需攝食營養豐富餌料進行能量補充。
3.4 禾花鯉消化酶活性變化與餌料消化利用
由本試驗結果看,2~4 d是仔魚內源性營養轉換為外源性營養的轉換期;10~17 d是仔魚轉變為稚魚的轉變期。仔魚在轉變期間攝食不足時,機體會通過水解體內貯存的脂肪來補充能量,此時可適量增加餌料脂肪的占比。轉換期前仔魚具有較高的淀粉酶活性,這可能與本身卵黃囊中含有較豐富的糖原有關[19]。轉換期后淀粉酶活性逐漸降低,可能是卵黃囊消失使糖原下降所致。這反映了8 d前仔魚發育階段的碳水化合物被用作能量消耗,不是作為營養被吸收[21]。8 d后淀粉酶活性隨之增強,說明此時仔魚可能開始適應對碳水化合物的利用[23];22 d后淀粉酶活性先降后升,可能是消化器官發育與糖代謝調節不同步所致。高活性淀粉酶可促進藻類的消化,第22天禾花鯉對黃藻、甲藻和硅藻有明顯的攝食,刺激淀粉酶的合成和分泌。有研究表示,鯉魚可有效地利用碳水化合物作為能量來源[24]。因此,仔魚后期時可在餌料中添加糖的占比為機體提供能量。
禾花鯉仔魚開口前堿性蛋白酶活性較低,開口攝食后活性增加,可能在一定程度上受到外源食物刺激酶分泌增加的影響。禾花鯉仔稚魚前期(4~10 d)時堿性蛋白酶活性較酸性蛋白酶高,14 d后稚魚主要以動物蛋白餌料為食,且轉變為以酸性蛋白酶消化蛋白為主。吉紅等[8]認為,由于消化器官和組織發育的完善,對蛋白消化起主導作用的酶種類會發生轉變;常青等[25]表示,在仔稚魚后期酸性蛋白酶在蛋白消化過程中更為重要,且在仔魚發育早期較高活性的堿性蛋白酶可以補償酸性蛋白酶的缺乏。說明兩者在早期發育階段可以起到互補作用。
4 結 論
綜上,禾花鯉屬雜食性魚類,仔稚魚的攝食與消化器官發育、消化酶活性、養殖環境中餌料組成有關;仔魚開口后消化酶活性會受消化器官生長發育的影響;隨著消化器官發育的完善,禾花鯉需要攝食營養豐富餌料進行能量補充。因此,為促進禾花鯉仔稚魚的生長和提高育苗成活率,在苗種培育過程可根據不同發育時期消化系統發育、攝食選擇以及消化酶變化的綜合體現來相應調整飼料配方。本試驗結果表明,3~14 d禾花鯉飼料適宜粒徑為50~170 μm,此時可在餌料中增加糖的占比,或在育苗池中同時培養浮游動物和植物;在轉變期10~14 d可適量增加餌料中脂肪的占比;14~30 d禾花鯉食性為雜食偏動物型,此時需營養均衡的配合飼料(蛋白和脂肪)并混合浮游動物投喂,飼料適宜粒徑為170~350 μm。
