不同溫度作用下草魚胚胎的發育及hsp70基因表達
2021-11-23 13:52:24邱裕明郭紅會李大鵬
水產科學 2021年6期
邱裕明,林 旺,郭紅會,張 策,李 莉,李大鵬
( 華中農業大學 水產學院,池塘健康養殖湖北省工程實驗室,水產養殖國家級實驗教學示范中心,湖北 武漢 430070 )
草魚(Ctenopharyngodonidella)作為我國四大家魚之一,在水產行業中占有重要的地位,根據2019年中國漁業統計年鑒數據,草魚以5.5043×106t的產量位居淡水養殖魚類產量第一[1]。相對來說,草魚與其他水產動物一樣,在個體發育的早期階段,其死亡率最高。然而,在自然條件或人工飼養條件下,常常會發生由天氣變化、給排水、活魚運輸等原因引起的溫度驟變,對魚類胚胎的發育和存活、幼魚生長等造成嚴重的影響。在研究水溫對異育銀鯽(Carassiusauratusgibelio)“中科3號”[2]胚胎發育的影響中發現,孵化溫度變化對于胚胎孵化過程影響較大,若水溫過低會減緩胚胎發育速度,當孵化溫度高于28 ℃,不僅會使孵化率顯著降低,胚胎畸形率也顯著升高,當水溫達到32 ℃時胚胎發育最差,孵化率僅為(2.3±1.0)%。亦有報道稱,胚胎不同發育階段對溫度的敏感程度不同。于歡歡等[3]認為,原腸中期可能是雜交子代胚胎發育過程中一個受溫度影響的重要敏感期。周鑫等[4]發現,短期內當水溫超過28 ℃時,草魚幼魚體內氧自由基增加,極大影響其免疫應答,最終將導致草魚幼魚的大規模死亡。可見,適宜的溫度對魚類維持正常的生理活動具有重要作用,其對胚胎發育的影響也會進一步影響到仔魚的孵化率和存活率。早在20世紀80年代就有一些有關水溫對魚類胚胎發育影響的研究[5-9]。如薛家驊[8]將胚胎置于14~15、24~27、32~33 ℃孵化,發現溫度可影響胚胎發育時間;郭永燦[9]分析認為,草魚胚胎發育最適溫度為23~28 ℃。然而上述研究大部分停留在表觀指標的統計上,如孵化率、畸形率、死亡率、胚胎各階段發育歷時、胚胎形態等,缺乏對不同溫度下魚類胚胎潛在的適應或影響機制的研究。
熱休克蛋白(hsp)家族具有協助變性蛋白復性以應對機體產生的不良影響的作用[10],是對溫度極為敏感的分子標記[11-12]。這其中,hsp70是最主要、最保守、細胞內含量最豐富的一類,它廣泛地參與了穩定新生多肽過程[13],具有幫助蛋白質進入線粒體和內質網[14]等功能。已有研究證明,熱應激會誘導機體內hsp70基因相對表達量的上升[15-17],不僅可以協助魚類抵御高溫應激,也有助于耐受其他應激源的刺激[18]。盡管生物體內存在調控機制以應對外界環境變化,但一旦超過其承受限度,將有可能會對機體造成不可逆的傷害。
筆者以草魚胚胎為研究對象,通過設置不同溫度梯度,探究溫度對草魚胚胎孵化耗時、孵化率、畸形率、初孵仔魚體長等生物學指標的影響,并測定初孵仔魚hsp70基因的表達動態,旨在為草魚的發育生物學提供基礎資料,同時為生產上草魚苗種繁育以及資源增殖提供一定的參考依據。
1 材料與方法
1.1 試驗材料
試驗在湖南省岳陽市華容縣田家湖水產場進行。選用基地馴化多年的草魚親本,人工催產后,采集干法人工授精的草魚胚胎作為試驗對象。水源為曝氣24 h以上的自來水,孵化容器為圓筒形塑料桶,每個桶內裝有1 L曝氣水,通過水浴控制桶內水溫。
1.2 試驗方案
設置22、25、28、31 ℃ 4個溫度組,每組3個平行,溫差不超過±0.5 ℃。每個孵化桶內置剛孵化的草魚卵150個,孵化過程中微曝氣,保證魚卵在水中翻動且卵膜不會被氣流沖破。為準確地統計胚胎受精率,每個溫度組另設3個孵化桶,各置100個草魚卵,待胚胎發育至原腸胚期后測定受精率。孵化過程中定期選擇不同溫度一定數目的胚胎在顯微鏡下觀察其發育情況,待魚苗全部孵化記錄孵化時長,并統計孵化率、畸形率,計算有效積溫和溫度系數Q10。每組隨機選取30尾初孵仔魚,記錄體長。收集不同溫度組的初孵仔魚樣品,立即凍存于液氮中直至檢測。
試驗各項指標按下式計算:
F=n1/n2×100%
(1)
H=n3/(n2×F)×100%
(2)
M=n4/n3×100%
(3)
K=N(T-C)
(4)
(5)
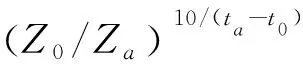
(6)
式中,F為受精率(%),H為孵化率(%),M為畸形率(%),n1為受精卵數(個),n2為觀察卵子總數(個),n3為孵出仔魚數(尾),n4為畸形仔魚數(尾);K為胚胎發育的有效積溫(℃·h),N為胚胎完成發育所需要的時間(h),T為胚胎發育的水溫(℃),C為胚胎發育的生物學零度(℃);n為試驗設計的溫度梯度個數,V為胚胎發育所需要時間的倒數(1/N),即發育速率;Q10[5]表示溫度改變10 ℃時發育速度加快的倍數,Z0和Za分別表示溫度t0和ta時胚胎孵化持續的時間(h)。
1.3 熒光定量PCR
使用TRIzol試劑盒提取胚胎樣品總RNA,利用NanoDrop ND-2000分光光度計(Thermo Scientific, Wilmington, DE, USA)和1.2%瓊脂糖凝膠電泳檢查總RNA濃度和完整性。取1 μg總RNA,用PrimeScriptTMRT Reagents反轉錄試劑盒合成cDNA(TaKaRa),并保存于-20 ℃冰箱直至檢測。
使用Primer Premier 5.0軟件設計基因特異性引物(表1)。在LighrCycler?480實時PCR檢測系統(Roche, Basel, Switzerland)上檢測hsp70基因相對表達量。PCR反應體系總量20 μL:2 μL cDNA模板,10 μL iQTMSYBR?Green Supermix,上下游引物(0.25 μmol/L)各0.8 μL,6.4 μL雙蒸水。PCR反應條件為:95 ℃預變性30 s,95 ℃變性10 s,56 ℃退火20 s,72 ℃延伸40 s,進行40個循環。本次試驗以β-actin基因為內參基因,采用2-△△Ct法計算目的基因的相對表達量比(R)。qPCR分析中使用6個生物學重復。
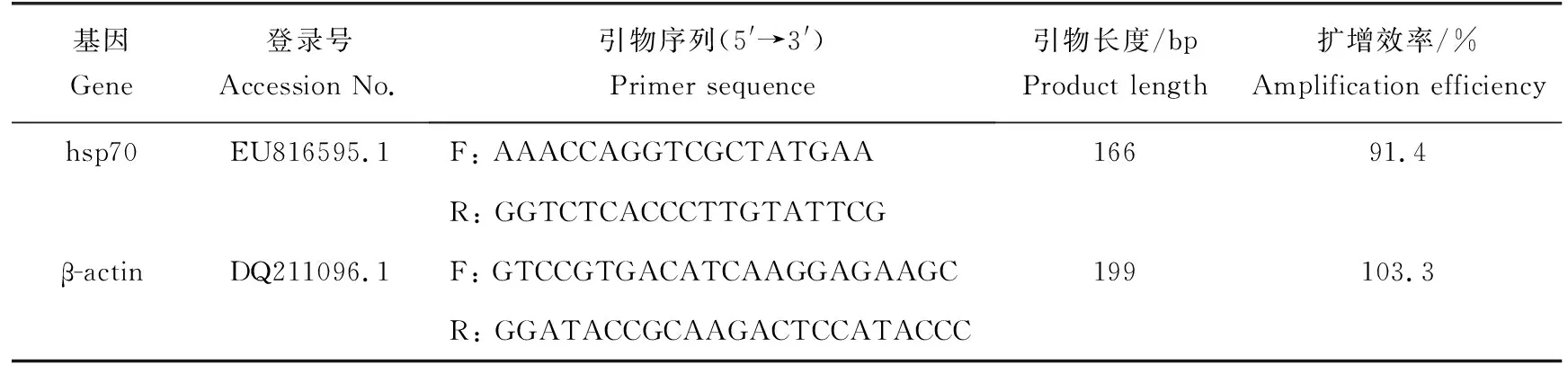
表1 熒光定量PCR所用引物序列
1.4 統計分析
利用SPSS 20.0對所有數據進行單因素方差分析和LSD多重比較,并數據表示為平均值±標準誤(n=6)。采用字母標記法表示每個數值之間的差異顯著性。
2 結 果
2.1 溫度對草魚孵化率和畸形率的影響
草魚胚胎的孵化率隨著溫度升高先上升,在28 ℃達到最高[(97.00±1.81)%],而31 ℃條件下孵化率顯著降低,僅為(31.51±1.84)%(P<0.05)(圖1)。溫度對畸形率的影響更為明顯,隨著溫度升高畸形率顯著降低(P<0.05),在28 ℃達到最低值[(2.08±1.06)%],31 ℃,大部分不能正常發育,畸形率達(90.00±5.79)%。通過回歸分析,得到孵化率(y)同溫度(x)之間的回歸方程:y=-0.0192x2+0.9695x-11.246(r2=0.872);畸形率(y)同溫度(x)之間的回歸方程:y=0.0271x2-1.3608x+17.035(r2=0.931)。
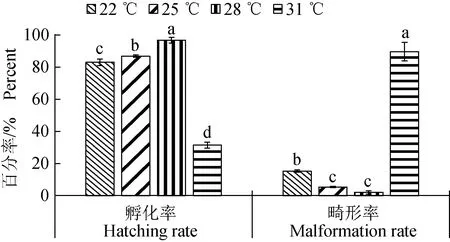
圖1 不同溫度條件下草魚胚胎孵化率和畸形率Fig.1 The hatching rate and malformation rate of grass carp C. idella embryos at different temperatures數據用平均值±標準誤表示(n=6),不同小寫字母表示有顯著性差異(P<0.05);下同.Data are expressed as mean±standard error (n=6), and means with different letters are significant difference (P<0.05);et sequentia.
2.2 溫度對胚胎孵化耗時和有效積溫的影響
隨著孵化溫度的升高,孵化耗時顯著降低(P<0.05)(表2)。相對的,草魚胚胎孵化的有效積溫在22~28 ℃隨溫度的升高而顯著增加(P<0.05),28 ℃時達到最大值,為(351.48±4.04) ℃·h,隨后在31 ℃顯著下降(P<0.05)。
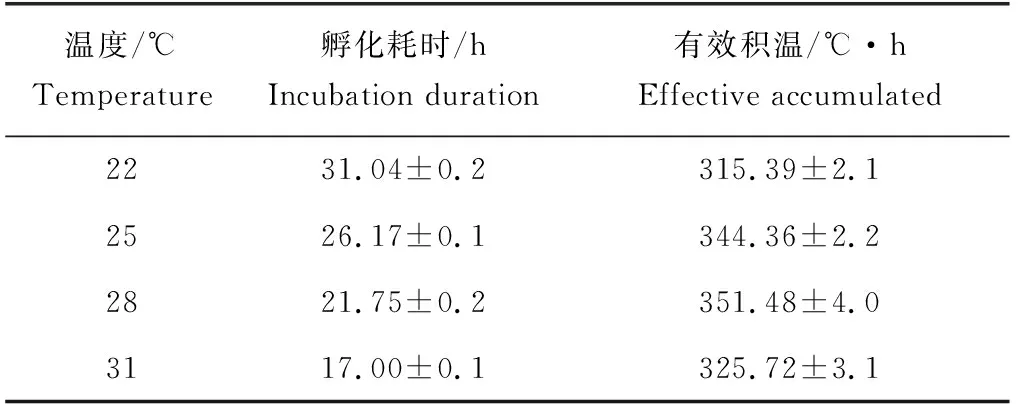
表2 不同溫度條件下草魚胚胎發育時間和有效積溫
試驗溫度為22~31 ℃,Q10變化為1.77~2.27,其中25~31 ℃溫度帶內的Q10值為2.05,接近2(表3)。
2.3 溫度對初孵仔魚體長的影響
隨著孵化溫度的上升,初孵仔魚的體長呈現先增后降的趨勢,其中22 ℃組體長最小,28 ℃體長最大,25 ℃和28 ℃之間無顯著差異,其他組之間差異顯著(P<0.05)(圖2)。通過回歸分析,得到初孵仔魚體長(y)同溫度(x)之間的回歸方程:y=-0.0357x2+1.9394x-21.138(r2=0.999)。
2.4 溫度對初孵仔魚hsp70基因相對表達水平的影響
對初孵仔魚體內hsp70基因的檢測發現,隨著溫度的上升,其相對表達水平顯著上升(P<0.05),且在28 ℃達到最高,28 ℃和31 ℃之間差異不顯著(圖3)。
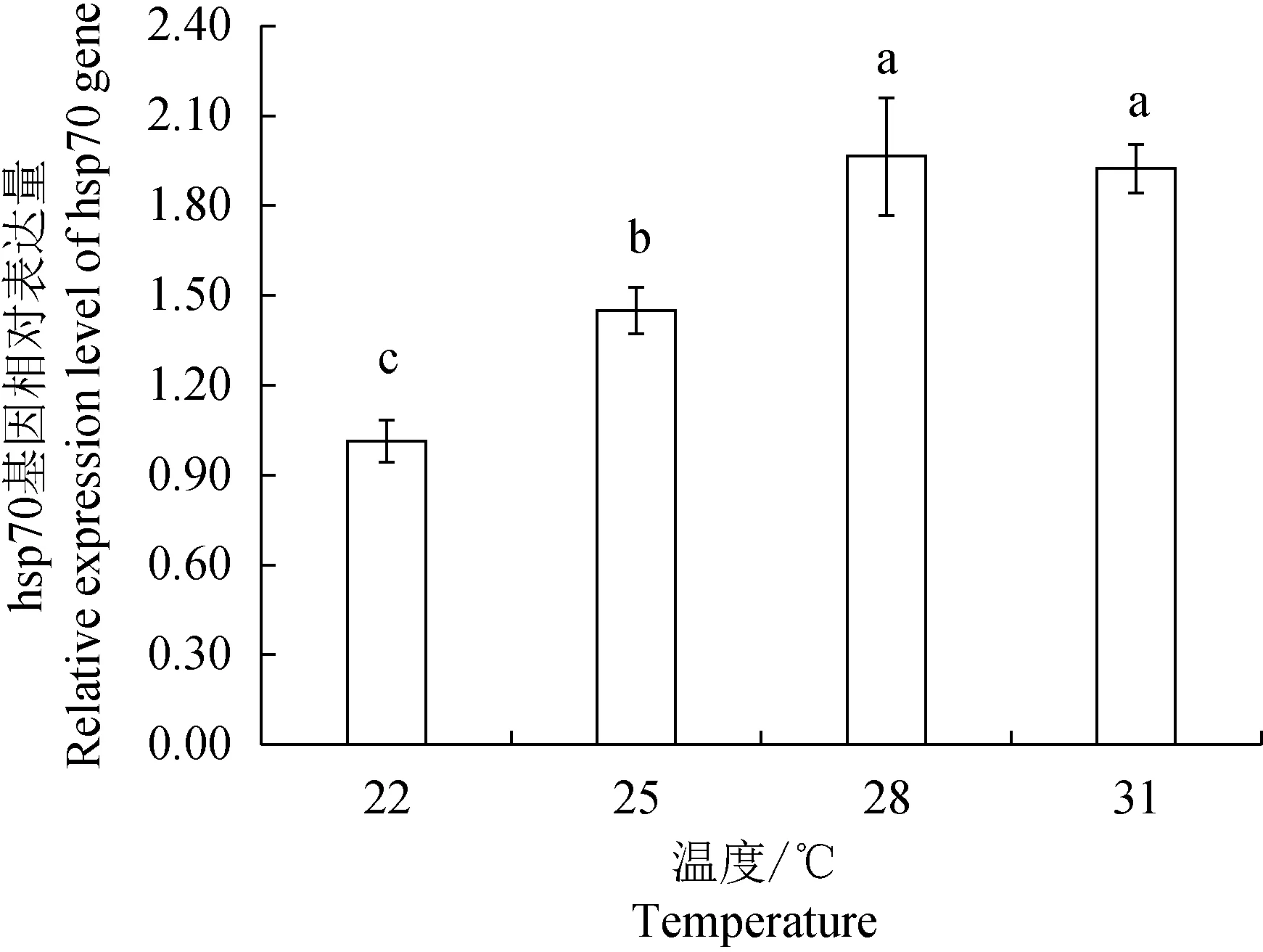
圖3 不同溫度條件下草魚仔魚hsp70基因相對表達量Fig.3 Relative expression level of hsp70 gene of grass carp C. idella larvae at different temperatures
3 討 論
3.1 不同溫度對草魚胚胎發育的影響
魚類胚胎在發育過程中對水溫的變化較為敏感,高溫或低溫的刺激往往會導致胚胎發育畸形、滯育,甚至死亡[5]。本試驗結果顯示,隨著孵化溫度的上升,草魚胚胎的孵化率和初孵仔魚的體長先上升,而后在31 ℃組顯著下降;相反的,畸形率在22~28 ℃先下降,在31 ℃下顯著上升。姜禮燔等[19]將草魚卵在36 ℃下急性暴露1、5、10、40 min,或在10 ℃暴露5、10、15、30 min后,再將魚卵移至19~21 ℃水體內孵化,發現高溫組內4種處理時間均使孵化率降低,而在低溫下暴露超過30 min也會使胚胎發育遲緩或畸形。在對黑莓鱸(Pomoxisnigromaculatus)[5]、淇河鯽(Carassiusauratus)[6]、尖裸鯉(Oxygymnocyprisstewartii)[20]、四川華鳊(Sinibramataeniatus)[21]等魚類的研究中也有類似的結果,即在適宜的溫度范圍內,隨著溫度升高,胚胎發育的孵化率增加,但是溫度過高或過低反而會影響孵化率,并導致胚胎發育畸形。事實上,作為胚胎發育過程中的主要營養來源,卵黃物質一方面提供能量,一方面也為胚胎組織、器官的構建提供物質基礎,而這其中需要大量酶的參與[22]。在溫度較低的情況下,酶的活性被抑制,阻礙胚胎細胞的代謝,使發育時間延長,如在凡納濱對蝦(Litopenaeusvannamei)的研究中發現,水溫的降低會延長受精第一、第二極體的釋放時間,最終影響孵化耗時[23]。而在溫度較高時,酶活性亦有可能被抑制,甚至失活,導致胚胎細胞膜的滲透性發生變化,分化過程紊亂,致使受精卵的正常發育嚴重受阻[24]。這可能是本試驗中31 ℃下草魚胚胎幾乎全部畸形的原因之一。除此之外,也有研究認為,胚胎某些結構可能對高溫更加敏感[25]。
本試驗結果顯示,草魚胚胎孵化耗時隨著溫度的升高顯著下降,但有效積溫隨著溫度的升高顯著上升,在28 ℃達到最大值,隨后在31 ℃下顯著下降。有效積溫代表胚胎發育過程中累計所必需的熱量總數,理論上是個常數,但在異育銀鯽[2]、扁吻魚(Aspiorhynchuslaticeps)[26]、玫瑰無須鲃(Puntiusconchonius)[27]等研究中均發現,當孵化溫度變化時,有效積溫會隨之出現波動。前期研究也證明,胚胎發育的不同階段對溫度變化的敏感性可能不同[5,26-28]。這或許說明溫度對胚胎的作用機制并不僅限于量的積累,還可通過參與調控胚胎發育過程中復雜的分子途徑,影響其代謝及器官分化等過程,因此有效積溫在一定范圍內波動。一般認為,溫度系數Q10越接近2,該溫度帶內水溫變化對胚胎發育的影響越小,即該溫度范圍為所研究對象的最適溫度范圍。本試驗結果發現,25~31 ℃溫度帶內的草魚胚胎發育Q10值為2.05,接近2,結合此溫度范圍內孵化率和畸形率的變化,可進一步認為草魚胚胎發育最適溫度應為25~28 ℃。
3.2 不同溫度對草魚胚胎hsp70基因表達的影響
溫度脅迫會導致機體活性氧增加,進而引起蛋白質的錯誤折疊,使蛋白質失活,影響機體生理過程。此時熱休克蛋白的大量轉錄翻譯可重折疊變性蛋白,阻止更多蛋白聚集,以維持機體穩態[29-31]。不僅如此,胚胎在發育過程中有大量蛋白質合成,熱休克蛋白對新合成蛋白質也具有一定的保護作用[13]。已有研究發現,高溫可誘導雞[32]、牛[33]、蜥蜴[34]等動物體內hsp70基因mRNA水平上升。有關鮭魚的研究表明,水溫由15 ℃升至22 ℃時,鮭魚體內hsp70基因水平急劇增加[35]。Mottola等[36]將斑馬魚(Daniorerio)在高于正常飼養水溫5 ℃或7 ℃ 暴露30 min后,發現斑馬魚體內hsp70基因的相對表達量顯著上升,且在+7 ℃組內hsp70水平也出現上調。Hallare等[37]在對斑馬魚胚胎所做的溫度—鎘聯合暴露試驗中發現,相對于低溫組(21 ℃),對照組(26 ℃)和高溫組(33 ℃)胚胎中hsp70基因相對表達量上升,胚胎抵御鎘刺激的能力增強,從而使得死亡率降低、平均心率升高。類似的,低溫脅迫也可誘導hsp70基因的表達,如虎皮魚(Puntiustetrazona)腦、鰓和肌肉組織中hsp70基因mRNA水平隨溫度降低而逐漸升高,表現為溫度依賴性表達[38]。本試驗結果顯示,孵化溫度的上升促使草魚初孵仔魚體內hsp70基因相對表達量的上升,表明草魚可通過調節熱休克蛋白hsp70基因的表達量以適應溫度的變化,保護胚胎的正常發育。
然而,熱休克蛋白僅能在一定水平的應激強度范圍內保護機體[4,39,40],如將文蛤(Meretrixmeretrix)長時間暴露于pH過高或過低的環境中,均會導致其機體內hsp70基因表達量的下降[41]。Cardoso等[39]將北美鯧鲹(Trachinotuscarolinus)幼魚暴露于高溫環境下,檢測到魚體內hsp70基因和p53基因相對表達量隨溫度升高而上升,在36 ℃組則顯著降低,并伴隨著行為失常,死亡率升高。周鑫等[4]在草魚幼魚急性溫度變化暴露試驗中也發現,鰓和肌肉中的hsp70基因和hsp90基因的相對表達量隨溫度升高而升高,繼而在34 ℃組降低,死亡率也迅速提高。通過整合基因相對表達量、耗氧率及半致死溫度數據發現,草魚hsp基因的表達過程需要能量的支持,且同機體代謝顯著相關,并進一步證實,草魚的生理臨界溫度為28 ℃[4]。事實上,由于hsp基因家族大多數成員沒有內含子,在應激條件下,hsp70基因的mRNA會優先轉錄翻譯,而其他蛋白的mRNA轉錄翻譯則會受到抑制[11,41]。本試驗中,28 ℃組hsp70基因相對表達量達到最高值后,31 ℃組沒有進一步升高,這可能是由于胚胎發育過程中并沒有外源性營養,不斷合成熱休克蛋白勢必會影響其他蛋白的合成,導致機體組織細胞膜結構和蛋白質組成異常,最終也會使胚胎發育受阻。本試驗hsp70基因的mRNA水平的變化一方面說明28 ℃可能已達到草魚維持胚胎正常發育的極限溫度,另一方面,可能正是因為31 ℃組熱休克蛋白合成量無法繼續升高,不足以抵御高溫侵襲,最終導致了孵化率的降低和畸形率的升高。
4 結 論
本試驗結果顯示,水溫25~28 ℃時,草魚胚胎的受精率、孵化率、有效積溫和初孵仔魚體長均較高,而孵化耗時,畸形率較低,結合Q10數據分析,建議草魚胚胎孵化的最適溫度為25~28 ℃。同時,hsp70基因相對表達水平在25~31 ℃隨溫度升高先升后降,表明草魚可通過調節hsp70基因的表達量以適應溫度的變化,保護胚胎的正常發育,且28 ℃可能是胚胎正常發育的極限溫度。
