不同種質番茄材料抗番茄黃化曲葉病毒病特性研究
2021-11-27 13:33:46魏小紅朱曉林王寶強
浙江農業學報 2021年11期
王 賢,劉 放,魏小紅,朱曉林,王寶強
(甘肅農業大學 生命科學技術學院,甘肅省作物遺傳改良與種質創新重點實驗室,甘肅省干旱生境作物學重點實驗室,甘肅 蘭州 730070)
番茄(Solanumlycopersicum)在世界范圍內廣泛種植,其營養豐富、適應性廣、食用方式多樣,是全球總產量最高的30種農作物之一,在世界農業經濟和農業市場中占有重要的地位[1]。但長期以來,番茄的生產受到許多病害的影響,尤其是番茄黃化曲葉病毒病(tomato yellow leaf curl virus disease,TYLCVD)。番茄黃化曲葉病毒(tomato yellow leaf curl virus,TYLCV)傳播媒介為煙粉虱,因此也被稱為煙粉虱傳雙生病毒(whitefly-transmitted geminiviruses,WTG),可侵染茄科、豆科等20多種植物[2]。該病具有發病率高、危害大、傳播效率高等特點[3]。番茄感染TYLCV后,葉片變形黃化,無法正常開花結果,對果實顏色、口感及營養成分影響極大,使其綜合品質降低,減產甚至絕產。1940年TYLCV在以色列首次被發現,隨后開始大規模暴發,20世紀90年代末亞洲各國相繼發現該病毒病,1991年我國首次在廣西南寧發現該病毒,2006 年起該病毒傳播到北京、山西、江蘇、上海、廣東、廣西、云南等地,具有關部門統計,該病毒對我國番茄產業造成的經濟損失每年高達十多億元,嚴重威脅我國番茄產業的發展[4-5]。該病毒最有效的解決方法是通過育種手段培育新的抗病品種,因此篩選抗病性較強的材料迫在眉睫。
番茄抗性品種的選擇中,表型、抗氧化酶活性、總酚與類黃酮含量以及抗性基因的表達等作為主要的抗性指標被用來鑒定不同材料的抗病能力[6]。植物體感染病原菌后,體內活性氧過量積累,而超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)是普遍存在于植物體內一類重要的保護酶,可以清除體內過剩的活性氧[7],另外多種抗氧化酶相互協調作用,可以增強植物對病害的抵抗力[8-11]。鄒芳斌等[7]研究顯示,抗氧化酶活性的升高與黃瓜抗病能力呈極顯著正相關。Ji等[12]的研究顯示,SPA(sodium pheophorbide a)可以通過提高抗氧化酶活性來控制櫻桃番茄對灰霉病的抗性。因此,抗氧化酶含量的變化可以作為植物抗病性機制發生的重要指標之一。次生代謝產物在植物的許多生長代謝過程中發揮著不可或缺的作用,其中最主要的是酚類,酚類中最主要的是類黃酮。研究發現,酚類物質與植物的抗病過程密切相關[13],邱永祥等[14]認為,感病后期抗病品種中的次生代謝物含量高于感病品種。植物感病后體內的莽草酸或乙酸途徑會積累大量的酚類物質,抑制病原菌進而達到抗病的目的。類黃酮在植物抗病毒與抗菌中發揮重要作用,可以清除植物在非正常狀態下產生的自由基進而在一定程度上降低植物的損傷[15]。早期的栽培番茄品種中沒有抗TYLCV的基因,而在野生番茄中蘊含著豐富的抗病基因,如最初發現的醋栗番茄(Solanumpimpinellifolium)和秘魯番茄(Solanumperuvianum)等,目前已被報道的抗病基因主要有Ty-1、Ty-2、Ty-3、Ty-3a、Ty-4和Ty-5,帶有抗病基因的抗性植株被病毒侵染后,體內病毒含量很低,不表現明顯的感病癥狀[16-17]。因此,了解植物抗病基因在細胞內的表達程度對于研究植物抗病水平具有重要作用,植物抗病基因的表達分析已廣泛應用于生命科學的各個領域。
由于番茄黃化曲葉病對番茄產業的影響巨大,抗病性的鑒定與檢測大多集中在表型或某一單一的抗性指標,目前綜合多種抗性指標研究番茄抗黃化曲葉病的報道較少。本實驗通過選取自育的15份番茄種質材料。利用苗期農桿菌接種法接種TYLCV,從表型、帶毒率、抗氧化酶活性、次生代謝物含量、抗性基因表達多個方面綜合研究不同種質的抗病性,旨在全面鑒定不同種質的抗病能力并篩選出幾個抗病性較強的材料,為抗病育種材料的選擇及生產應用提供理論依據。
1 材料與方法
1.1 試驗材料
1.1.1 材料
編號分別為801、802、803、805、806、817、818、819、820、822、841、842、853、857、867,以及感病品種桃太郎Money Maker(MM)與抗病品種齊達利兩個對照品種,番茄種子由甘肅張掖益新泉蔬菜育種公司提供(齊達利由瑞士先正達種子公司提供,MM由中國農業科學院蔬菜花卉研究所鮮食番茄課題組提供),所攜帶抗性基因見下表1。用于苗期接種的 TY-DNA 侵染性克隆 pBin-PLUS-1.7A+2β(農桿菌)引自浙江大學。

表1 不同番茄材料的基因型
1.1.2 主要試劑
總RNA提取試劑盒,FastKing cDNA第一鏈合成試劑盒,熒光定量PCR試劑盒,植物基因組DNA提取試劑盒購于天根生化科技有限公司。
1.2 方法
1.2.1 番茄苗的培養
將17份番茄種子用10%次氯酸鈉溶液浸泡10 min后播種于50孔的育苗穴盤中,置于育苗基地培養,長出1~2片真葉后定植于花盆中繼續培養,每份種質各定植10株。
1.2.2 pBin-PLUS-1.7A+2β的培養
事先分別配置固體與液體的LB培養基,配置過程中加入抗生素Kan(50 mg·L-1)和 Rif(50 mg·L-1),先將含 pBin-PLUS-1.7A+2β的農桿菌菌液涂布于固體LB培養基上,于28 ℃培養箱倒置暗培養至長出單菌落,后挑選合適的單菌落轉接于LB液體培養基中,放于200 r·min-128 ℃的搖床過夜,于600 nm處測量吸光值且處于0.6~0.8時,方可達到接種要求。
1.2.3 接種病毒
參考張少麗等[18]的方法,采用農桿菌接種法接種TYLCV。選取長勢最佳的植株過量澆水,以便病毒侵入,采用1 mL注射器吸取菌液從葉背注射于植株體內,注射時以可觀察到菌液滲入為準,每株約0.2 mL。接種完成后分別于20、30、40 d調查發病情況并采樣,進行超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)的活性、總酚與類黃酮含量以及抗性基因表達測定,試驗設3次重復。
1.3 測定項目與方法
1.3.1 帶毒率的檢測
在接種20 d后對各材料進行 PCR 檢測,以此判斷該品種對 TYLCV的易感程度。參考李佳蔚[19]的研究合成 TYLCV特異引物V2(F: 5′-GGGCCACGATTTAATTAGGGATCTTAT-3′; R: 5′-TACATTCTGTATATTCTGGGCTTCCG-3′),其擴增長度為229 bp。PCR體系為20 μL,其中模板DNA 1 μL;Master Mix 10 μL、上游引物1 μL、下游引物1 μL、ddH2O 7 μL。95 ℃變性5 min后開始循環,95 ℃變性30 s,61 ℃退火30 s,72 ℃延伸1 min,30個循環結束后,72 ℃繼續延伸7 min,4 ℃保存。PCR產物于1.0%瓊脂糖凝膠電泳25 min(150 V),最后用Goldview染色,凝膠成像系統拍照。
1.3.2 抗氧化酶活性的測定
參照Hu等[20]的方法,采用氮藍四唑(NBT)光化還原法測定超氧化物歧化酶(SOD)活性;參照裴斌等[21]的過氧化氫還原法測定過氧化氫酶(CAT)活性;采用紫外分光光度法測定抗壞血酸過氧化物酶(APX)活性。
1.3.3 次級代謝物的測定
總酚和類黃酮的測定分別參照唐巧玉等[22]和Pastrana-Bonilla等[23]。分別以沒食子酸與蘆丁繪制標準曲線并計算含量。
1.3.4 抗性基因的表達分析
采用天根生物公司RNA試劑盒分別于接種20、30、40 d提取番茄葉片中的總RNA,并用微量紫外分光光度計檢測其濃度和純度,然后用1%的瓊脂糖凝膠電泳檢測其完整性。根據FastKing cDNA試劑盒說明書進行反轉錄合成cDNA。參考天根公司的SYBR System操作說明進行實時熒光定量PCR反應。反應程序為95 ℃預變性15 min,95 ℃變性10 s,60 ℃退火20 s,72 ℃延伸20 s,40個循環。每種材料設3個重復。根據已知序列設計Ty-1、Ty-2、Ty-3基因的引物,以actin基因為內參,見表2。合成由陜西楊凌天潤奧科生物科技有限公司完成。
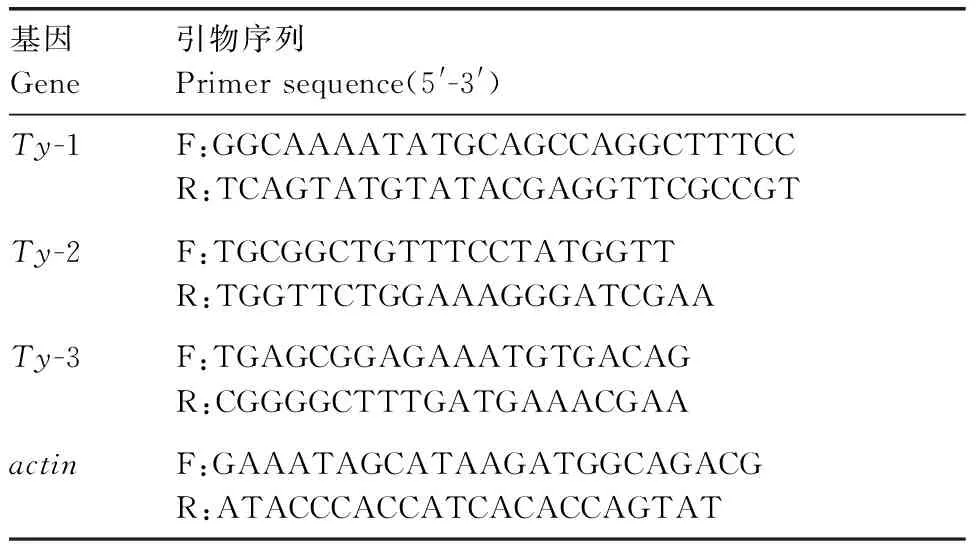
表2 抗病基因及引物序列
1.4 數據處理
采用 Microsoft Excel整理與分析實驗所得的數據,用SPSS 22.0 軟件對數據進行差異顯著性分析(字母法標記)與相關分析。
2 結果與分析
2.1 不同種質番茄材料的表型及帶毒率分析
2.1.1 表型特征分析
如圖1所示,所有苗齡為30 d的試驗植株接種TYLCV病毒后,選取各抗性水平下具有代表性的材料802、803、820、842、853、齊達利。與未感病植株相對比,802、803在感病20 d出現葉片卷曲、黃化、變形、變小并隨時間推移加重,至40 d時變形皺縮嚴重,表現為易感病;820在感病20 d與30 d癥狀并不明顯,而在40 d出現葉片黃化、卷曲、皺縮,842在感病20 d和30 d時癥狀不明顯,在40 d出現葉片輕微黃化,表現為不易感病;853與齊達利感病后在各時期無任何感病癥狀,表現為免疫。

A,對照;B,感病后表型。A, The control; B, The phenotype after infection.圖1 部分品種感病后的表型Fig.1 The phenotypes of some varieties after virus infection
2.1.2 帶毒率分析
檢測結果如圖2所示,17份番茄材料全部擴增出約229 bp的條帶,說明在接種后20 d各番茄材料中均含黃化曲葉病毒的DNA序列,病毒成功侵入植株體內,帶毒率為100%,說明各試驗材料成功接種TYLCV,可以進行后續的實驗與分析。該結果也表明了農桿菌接種法接種該病毒的可行性。
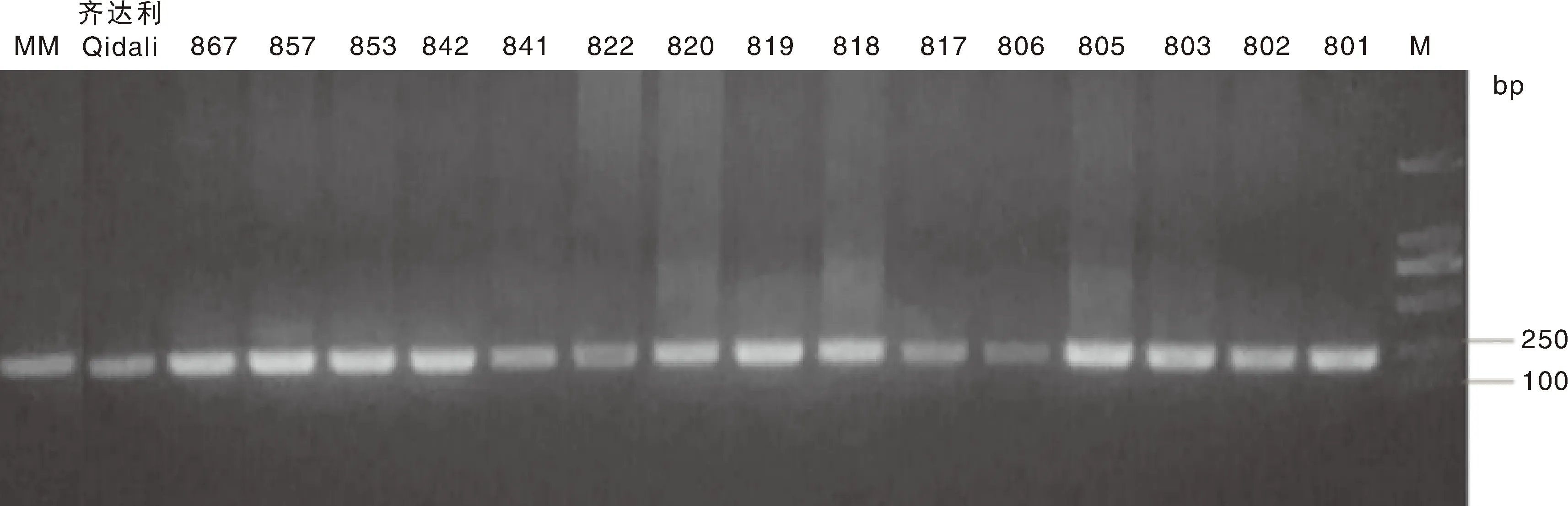
圖2 不同番茄材料接種20 d后的PCR擴增結果Fig.2 PCR amplification results of different tomato materials after virus inoculation for 20 days
2.2 不同種質番茄幼苗葉片抗氧化酶的分析
2.2.1 SOD活性分析
如圖3所示,感病后,805、817、819、820、841、842、853、857、867的SOD活性在各時期下均顯著高于陰性對照MM(P<0.05);853、867材料的SOD活性在各時期下甚至高于陽性對照齊達利,其中867具有顯著性差異(P<0.05);801的SOD活性在各時期下均低于陰性對照MM;在20 d時,867的SOD活性是陰性對照MM的1.80倍,801的SOD活性是陽性對照齊達利的0.55倍,在30 d時,853的SOD活性是陰性對照MM的1.50倍,806的SOD活性是陽性對照齊達利的0.70倍,在40 d時,867的SOD活性是陰性對照MM的1.56倍,801的SOD活性是陽性對照齊達利的0.69倍。

柱上沒有相同小寫字母表示差異顯著(P<0.05)。下同。The bars without the same lowercase letters showed significant difference(P<0.05). The same as below.圖3 不同番茄材料接種病毒后超氧化物歧化酶(SOD)活性的變化情況Fig.3 Changes of superoxide dismutase (SOD) activity in different tomato materials after virus inoculation
2.2.2 CAT活性分析
如圖4所示,感病后,817、842、853、857、867的CAT活性在各時期下均高于陰性對照MM,其中817、853、867具有差異顯著性(P<0.05);853材料的CAT活性在各時期下甚至高于陽性對照齊達利;在20 d時,857的CAT活性是陰性對照MM的2.46倍,819的CAT活性是陽性對照齊達利的0.44倍,在30 d時,867的CAT活性的是陰性對照MM的2倍,820的CAT活性是陽性對照齊達利的0.62倍,在40 d時,867的CAT活性是陰性對照MM的2.12倍,822的CAT活性是陽性對照齊達利的0.53倍。

圖4 不同番茄材料接種病毒后過氧化氫酶(CAT)活性的變化情況Fig.4 Changes of catalase (CAT) activity in different tomato materials after virus inoculation
2.2.3 APX活性分析
如圖5所示,感病后,801、805、817、818、819、820、822、841、842、853、857、867的APX活性在各時期下均高于陰性對照MM,其中817、819、820、841、842、853、857、867具有差異顯著性(P<0.05);842、853、857、867材料的APX活性在各時期下甚至高于陽性對照齊達利,其中853、867具有顯著性差異(P<0.05)。在20 d時,853的APX活性是陰性對照MM的3.12倍,803的APX活性是陽性對照齊達利的0.29倍,在30 d時,867的APX活性是陰性對照MM的2.81倍,803的APX活性是陽性對照齊達利的0.36倍,在40 d時,857的APX活性是陰性對照MM的2.1倍,803的APX活性是陽性對照齊達利的0.4倍。
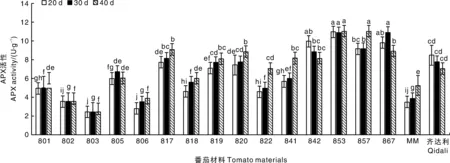
圖5 不同番茄材料接種病毒后抗壞血酸過氧化物酶(APX)活性的變化情況Fig.5 Changes of ascorbate peroxidase (APX) activity in different tomato materials after virus inoculation
2.3 不同種質番茄幼苗葉片總酚及類黃酮分析
2.3.1 總酚含量分析
如圖6所示,感病后,802、805、806、817、818、819、820、841、842、853、857、867的總酚含量在各時期下均高于陰性對照MM,其中805、806、817、819、820、841、842、853、857、867具有差異顯著性(P<0.05);853材料的總酚含量在各時期下甚至高于陽性對照齊達利;在20 d時,853的總酚含量是陰性對照MM的1.68倍,803的總酚含量是陽性對照齊達利的0.56倍,在30 d時,853的總酚含量是陰性對照MM的1.72倍,801的總酚含量是陽性對照齊達利的0.57倍,在40 d時,857的總酚含量是陰性對照MM的1.82倍,801的總酚含量是陽性對照齊達利的0.62倍。
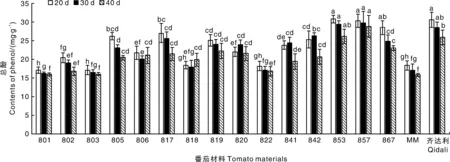
圖6 不同番茄材料接種病毒后總酚含量的變化情況Fig.6 Changes of total phenol content in different tomato materials after virus inoculation
2.3.2 類黃酮含量分析
如圖7所示,感病后,817、818、820、841、842、853、857、867的類黃酮含量在各時期下均高于陰性對照MM,其中817、853、857、867具有差異顯著性(P<0.05);857、867材料的類黃酮含量在各時期下甚至高于陽性對照齊達利,其中857具有顯著性差異(P<0.05);在20 d時,857的類黃酮含量是陰性對照MM的4.02倍,803的類黃酮含量是陽性對照齊達利的0.32倍,在30 d時,857的類黃酮含量是陰性對照MM的4.06倍,806的類黃酮含量是陽性對照齊達利的0.36倍,在40 d時,857的類黃酮含量是陰性對照MM的4.28倍,802的類黃酮含量是陽性對照齊達利的0.211倍。

圖7 不同番茄材料接種病毒后類黃酮含量的變化情況Fig.7 Changes in the content of flavonoids in different tomato materials after virus inoculation
2.4 不同種質番茄幼苗葉片抗病基因表達分析
2.4.1Ty-1基因表達分析
如圖8所示,857、867材料感病后Ty-1基因表達量在各時期下均高于陽性對照齊達利,其中867材料具有顯著性差異(P<0.05),801、802、803、805、806、817、818、819、820、841、842材料感病后Ty-1基因表達量在各時期下均低于陽性對照齊達利,其中除802、817、842外其他材料均具有顯著性差異(P<0.05);在20 d時,867的Ty-1基因表達量是陽性對照齊達利的 2.2倍,802的Ty-1基因表達量是陽性對照齊達利的0.53倍,在30 d時,867的Ty-1基因表達量是陽性對照齊達利的 1.4倍,801的Ty-1基因表達量是陽性對照齊達利的0.41倍,在40 d時,822的Ty-1基因表達量是陽性對照齊達利的1.4倍,818的Ty-1基因表達量是陽性對照齊達利的0.42倍,另外,總體來看,番茄植株接種病毒后Ty-1基因表達量在30 d、40 d時有明顯升高。
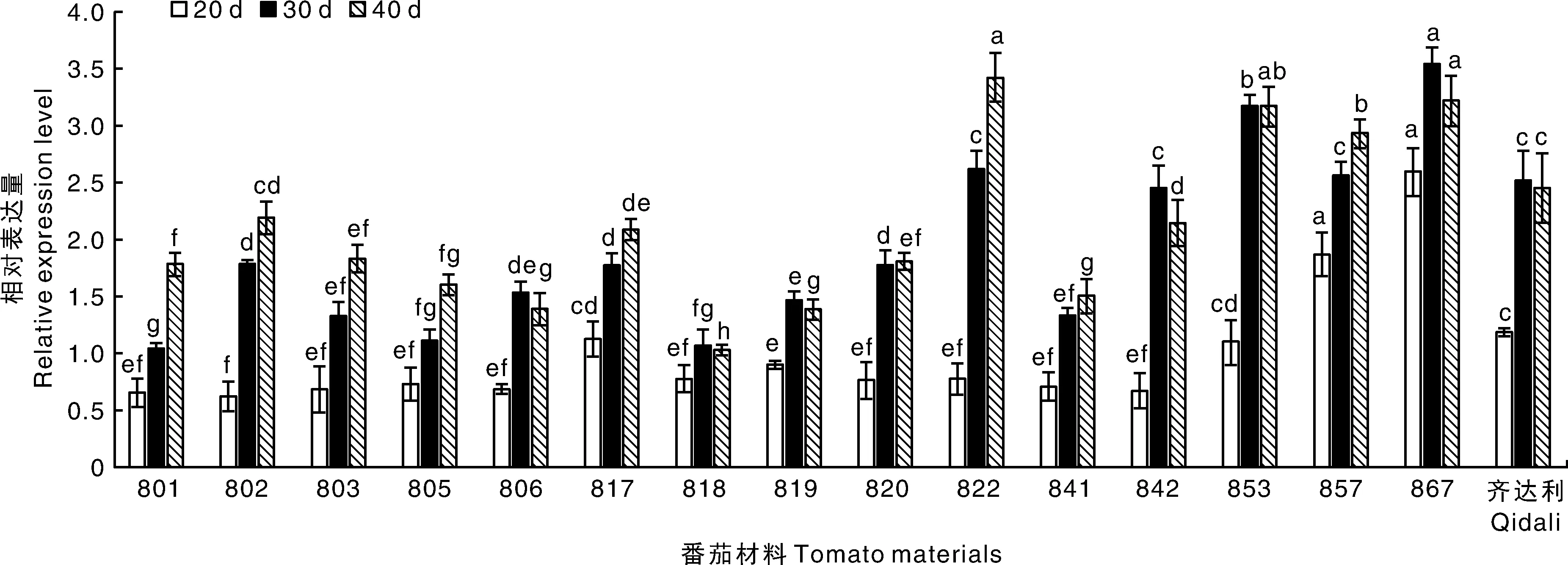
圖8 不同番茄材料接種病毒后Ty-1基因的表達情況Fig.8 Expression of Ty-1 gene in different tomato materials after virus inoculation
2.4.2Ty-2基因表達分析
如圖9所示,除853材料外,其他供試材料感病后Ty-2基因表達量在各時期下均低于陽性對照齊達利,且具有顯著性差異(P<0.05);在20 d時,853的Ty-1基因表達量是陽性對照齊達利的0.93倍,820的Ty-2基因表達量是陽性對照齊達利的0.31倍,在30 d時,853的Ty-2基因表達量是陽性對照齊達利的1.03倍,803的基因表達量是陽性對照齊達利的0.42倍,在40 d時,853的Ty-2基因表達量是陽性對照齊達利的1.03倍,801的Ty-2基因表達量是陽性對照齊達利的0.48倍。

圖9 不同番茄材料接種病毒后Ty-2基因的表達情況Fig.9 Expression of Ty-2 gene in different tomato materials after virus inoculation
2.4.3Ty-3基因表達分析
如圖10所示,842、857、867材料感病后Ty-3基因表達量在各時期下均高于陽性對照齊達利,其中857、867材料具有顯著性差異(P<0.05),801、802、803、805、806、817、818、819、820、841材料感病后Ty-3基因表達量在各時期下均低于陽性對照齊達利,均具有顯著性差異(P<0.05);在20 d時,857的Ty-3基因表達量是陽性對照齊達利的1.38倍,820的Ty-3基因表達量是陽性對照齊達利的0.31倍,在30 d時,867的Ty-3基因表達量是陽性對照齊達利的1.17倍,820的基因表達量是陽性對照齊達利的0.26倍,在40 d時,857的Ty-3基因表達量是陽性對照齊達利的1.53倍,820的Ty-3基因表達量是陽性對照齊達利的0.38倍。另外,842、853、857、867的Ty-3基因表達量高于其他材料,且存在顯著性差異(P<0.05)。

圖10 不同番茄材料接種病毒后Ty-3基因的表達情況Fig.10 Expression of Ty-3 gene in different tomato materials after virus inoculation
2.5 抗氧化酶、總酚、類黃酮以及抗病基因表達的相關性分析
由表3可以看出,番茄植株感染黃化曲葉病毒后,其體內的抗氧化酶活性的強弱、總酚與類黃酮的含量高低以及抗病基因表達量之間存在密切相關的關系,SOD活性、CAT活性、APX活性、總酚含量、類黃酮含量、Ty-1、Ty-2和Ty-3基因的表達量相互之間均呈顯著正相關(P<0.05),其中除總酚含量與Ty-1基因表達量之間呈顯著正相關外,其余各指標相互之間均呈極顯著正相關(P<0.01)。另外APX活性與SOD活性之間相關性最大,總酚含量與Ty-1基因的表達量之間相關性最小。

表3 各抗性指標的相關性分析
3 討論
黃化曲葉病毒病是目前影響番茄產量的主要病害之一,目前解決這一病害最有效的方法是通過育種手段培育新的抗病品種,但存在對抗性水平研究較少,了解不夠全面的問題。為了解帶有抗性基因的番茄材料對番茄黃化曲葉病毒病的抗性,對15份番茄以及兩份對照材料進行帶毒率 、抗氧化酶活性、總酚與類黃酮含量的檢測,并利用實時定量PCR方法分析抗性基因的表達水平。植物受到病毒侵害后,體內活性氧會大量積累,而抗氧化酶則可以清除多余的活性氧,從而減輕植物在病害下的損傷。因此,抗氧化酶活性的高低可反映出植物對病毒抗性的強弱[8,10],植物產生的次生代謝物與植物的抗病也密切相關,研究表明,酚類物質的積累可以抑制多數病原菌,植物受到侵害時酚類物質會快速積累,而感病品種則積累較慢,類黃酮物質可以提供質子供體而清除自由基,達到抗逆抗病的效果。因此,次級代謝物的含量高低也可反映出植物對病毒抗性的強弱[24-27]。
目前有關植物抗病性的研究中,多將抗氧化酶活性作為一個重要指標,王玲平[28]對黃瓜不同抗性品種的研究顯示,隨時間推移抗氧化酶活性均有不同程度升高且抗病品種中的SOD酶活性高于感病品種。丁九敏等[29]研究顯示,黃瓜接種霜霉病后,CAT、SOD活性均升高。但對APX的研究較少。賈雙雙等[30]研究顯示,高抗材料的總酚與類黃酮含量均顯著高于高感材料。本課題組之前的病情分析表明,供試材料中齊達利、853、857、867對TYLCV表現為免疫,805、817、819、820、841、842對TYLCV表現為高抗,MM、802、806、818、822對TYLCV表現為輕微抗病,801和803的抗病能力最弱。本研究結果表明,所有供試材料均成功感染TYLCV,從表型看,853沒有任何感病特征,802與803感病最為嚴重;867、853、857的抗氧化酶活性最高,其中853、867的SOD活性、853的CAT活性和842、853、857、867的APX活性在各時期下甚至高于陽性對照,另外在20d時853的APX活性是陰性對照的3.12倍;857、853、867 的次生代謝物含量最高,其中853的總酚含量和857、867的類黃酮含量在各時期甚至高于陽性對照,另外在40 d時,857的類黃酮含量是陰性對照的4.28倍;感病植株體內的各抗性指標相互之間均呈極顯著正相關(總酚與Ty-1之間為顯著正相關)。這與趙秀娟等[31]、齊紹武等[32]、賈雙雙等及本課題組之前的研究結論相符合。在15種供試材料中,857、853、867的抗氧化酶活性與次生代謝物含量均最高,初步推測抗氧化酶與次生代謝物之間存在因果關系。不同的供試品種對病毒的抵抗力表現不同,分析原因是不同的品種其基因型、遺傳物質以及體內防御系統存在差異。進而導致體內的抗氧化酶合成水平以及次生代謝物積累水平存在差異。
目前報道的番茄黃化曲葉病毒的抗性基因有Ty-1、Ty-2、Ty-3、Ty-3a、Ty-4和Ty-5等[33-34],這些基因在一定條件下可以抑制番茄黃化曲葉病毒的擴增、表達以及移動,從而從根本上達到抗病毒的目的[19],但不同抗性基因在番茄 TYLCV 病害防御中的機理報道較少。不同抗性基因對 TYLCV 病原的抗性水平不同,相關抗性基因在 TYLCV 病原侵染下的表達水平及差異鮮有報道[35]。 田兆豐等[36]用TYLCV感染不同基因型的番茄品種,采用qRT-PCR法分析基因表達,結果顯示,隨發病時間的延長,抗病基因Ty-1、Ty-3的基因表達量均升高,在不同品種中存在差異。本研究結果表明,867、857、853、842的抗病基因的表達量最高,857、867的Ty-1基因表達量、842、857、867的Ty-3基因表達量在各時期下均高于陽性對照,另外在20 d時867的Ty-1基因表達量是陽性對照的2.2倍。總體來看,植物接種病毒后Ty-1基因表達量在30、40 d時有明顯升高。這暗示出番茄感染病毒后,其抗病基因Ty-1大量表達的時間較其他抗病基因較遲,說明Ty-1基因表達具有延遲性,具體的機制有待進一步研究。抗病基因表達量越高的品種,抗氧化酶含量與次生代謝物含量也越高,其抗病能力也越強,表現出正相關性。分析原因可能是抗性基因的表達,促進了抗氧化酶的合成,也促進了次生代謝物的積累,也可能是抗性基因的表達與植株的抗病信號通路存在聯系,抗病基因的產物調控了信號分子,通過信號傳遞在生理水平上產生防御反應。另外發現802與803攜帶的抗性基因多于822,但抗病性卻弱于822,分析原因822可能存在其他的抗病途徑, 802與803在病毒引起的一系列逆境中的抵抗力較弱。綜合分析,植物的抗病系統是一個非常復雜的動態過程,受多種物質與因素的調節,眾所周知,基因的表達可以調節植物對逆境的應答,而植物抗病的應答又離不開酶這個 “員工”,植物的許多抗逆過程都是通過酶的介導完成的。本實驗中抗病基因相對表達量的升高與酶活性的升高、總酚與類黃酮含量的升高相互聯系,共同參與番茄植物的抗病過程,但具體如何聯系還有待于進一步研究。
4 結論
對不同番茄材料進行病毒檢測和抗性評價發現,所有研究材料均成功攜帶TYLCV,帶毒率為100%;不同材料的抗病能力表現如下, 853無任何的感病特征,802與803感病最為嚴重; 867、853、857的抗氧化酶活性較高,842與819次之,801與803較低; 857、853、867的總酚與類黃酮含量較高,817與842次之,822與803較低; 867、857、853、842的抗性基因表達水平較高,817與822次之,805與820較低。對各方面指標進行綜合分析,結果顯示,不同番茄材料中的抗病能力較高的為867、857、853三個材料,817與842次之, 803最低,可將867、857、853用作高抗性的育種材料或生產應用中,817與842作為替代材料。植株感病后體內的SOD、CAT、APX活性、總酚、類黃酮含量、Ty-1、Ty-2和Ty-3基因的表達量相互之間均呈極顯著正相關(總酚與Ty-1之間為顯著正相關),另外番茄植株感病后體內Ty-1基因的表達具有延遲性。
