大白菜BrSPS1Fb基因剪接受體位點變異及其對剪接的影響
2021-11-28 13:05:16岳智臣趙彥婷雷娟利李必元
浙江農業學報 2021年11期
陶 鵬, 岳智臣, 趙彥婷, 雷娟利, 李必元
(浙江省農業科學院 蔬菜研究所,浙江 杭州 310021)
在真核生物中,大部分基因是不連續的,包含一個或多個內含子,這些基因在轉錄成pre-mRNA后,需要剪切掉內含子才能形成連續的成熟mRNA[1]。Pre-mRNA剪切過程中遵守GU-AG法則(又稱為Chambon法則),在內含子5′端邊界序列GU(對應于DNA上的剪接供體位點GT)和內含子3′端邊界序列AG(對應于DNA上的剪接受體位點AG)進行剪切。真核生物的pre-mRNA中包含多個潛在的剪切位點,可通過選擇性剪接方式對pre-mRNA進行剪切以產生不同的成熟mRNA,導致其轉錄本與蛋白產物的多樣性[2]。選擇性剪接在植物生長發育和非生物脅迫響應中發揮著重要作用[3-4]。
基因在DNA水平上的剪切位點主要包括剪接供體位點(GT)、剪接受體位點(AG)和分支位點(A)。剪接復合體(spliceosome)可識別剪切位點,并將內含子準確切除[5]。保守的剪接位點可保證pre-mRNA加工的準確性。白菜FLC1基因的剪接供體位點從GT變異成AT后,導致其在pre-mRNA加工過程中發生了選擇性剪接,并導致白菜極易抽薹[6]。當基因的剪接受體位點(AG)變異后,將會如何影響pre-mRNA的剪接方式和剪接效率?大白菜He2是本課題組的核心材料,具有生長勢小、生長緩慢的特點。SPS1F編碼了一種蔗糖磷酸合成酶,其可與蔗糖磷酸酶(sucrose phosphate phosphatase)共同作用,促進植物的生長和生物量的積累[7]。研究大白菜He2的SPS1F基因變異情況,有可能從DNA水平說明大白菜He2生長勢小的原因。本研究通過重測序發現大白菜He2的BrSPS1Fb-He2基因第6個內含子的剪接受體位點上存在一個單核苷酸多態性(single nucleotide polymorphisms,SNP)變異。以該位點未變異的BrSPS1Fa-He2和大白菜Chiifu401的BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401為對照,分析了BrSPS1Fb-He2剪接供體位點變異后對其剪接方式和剪接效率的影響。
1 材料與方法
1.1 大白菜He2的重測序和轉錄組測序
大白菜He2生長勢小、配合力強、半直立、葉色綠、葉面平展、光澤度好、葉脈不顯、短筒球形,其高代自交系是由浙江省農業科學院蔬菜研究所大白菜甘藍組選育而成的骨干親本材料。以大白菜He2為材料進行研究可進一步從分子水平了解骨干親本,以更好應用于育種。在大白菜蓮座期提取其葉片DNA并建庫,采用Illumina測序平臺進行高通量測序。大白菜He2開花期,對花瓣取樣3次,提取樣品總RNA并進行檢測,構建文庫,使用qPCR方法對文庫的有效濃度進行準確定量,不同文庫按照目標下機數據量進行pooling,在Illumina平臺進行轉錄組測序。重測序與轉錄組測序工作由北京百邁客生物科技有限公司完成。
1.2 SPS1F基因鑒定和選擇性剪接分析
使用擬南芥SPS1F(AT5G20280)基因序列在大白菜參考基因組(Chiifu401)中檢索,并進行共線性分析。在大白菜LF和MF2亞基因組中篩選到2個SPS1F基因,分別命名為BrSPS1Fa(Bra002289)和BrSPS1Fb(Bra020096)。為區分大白菜He2和Chiifu401中的SPS1F,對He2中SPS1F命名為BrSPS1Fa-He2和BrSPS1Fb-He2,而Chiifu401中的SPS1F命名為BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401。
分別使用PA和PB序列在大白菜He2花瓣的轉錄組測序read文庫中檢索,可獲得BrSPS1Fa-He2和BrSPS1Fb-He2轉錄本read。基于大白菜參考基因組分析所獲得的read序列,將不同類型的read進行分類,并分別命名為S1、S2、S3、S4、S5,并分析這些類型剪接異構體的剪接方式。分別使用PA和PB序列在大白菜Chiifu401花的轉錄組測序read文庫(NCBI SRA登錄號:SRR643622)中檢索,可獲得BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401轉錄本read,參考上述S1-S5分類模式對read進行分類并記錄剪接方式。所使用的檢索序列見表1。

表1 所使用的檢索序列
1.3 SPS1F基因的剪接頻率分析
在大白菜He2花瓣的轉錄組測序read文庫中使用PB1-PB5及其反向互補序列進行檢索,可獲得BrSPS1Fb-He2基因的S1~S5類型剪接異構體的數量,再基于read數量計算S1~S5各種剪接異構體出現的比例。基于上述方法,使用PA1~PA5及其反向互補序列在He2轉錄組測序文庫進行檢索,并計算BrSPS1Fa-He2基因的S1~S5各種剪接異構體出現的比例。采用上述檢索序列和方法,在大白菜Chiifu401花的轉錄組測序read文庫(SRR643622)中檢索對應的read,計算BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401中各種剪接異構體的百分比。使用Excel作圖。
1.4 剪接異構體、剪接方式鑒定和蛋白產物預測
使用primer5將大白菜He2的BrSPS1Fb基因4種類型轉錄本(S1、S2、S3和S4)和大白菜Chiifu401的S5轉錄本(BrSPS1Fb-Chiifu401)轉換成蛋白。使用Clustal Omega將上述蛋白質氨基酸序列進行比對,以BrSPS1Fb-Chiifu401蛋白質氨基酸序列作為參考,標注大白菜He2 4種蛋白(BrSPS1Fb-He2-S1、BrSPS1Fb-He2-S2、BrSPS1Fb-He2-S3和BrSPS1Fb-He2-S4)的變異情況。
2 結果與分析
2.1 SPS1F的鑒定與變異
使用擬南芥AT5G20280序列在大白菜參考基因組(Chiifu401)中進行比對,發現大白菜中包含2個SPS1F基因,分別命名為BrSPS1Fa-Chiifu401(Bra002289)和BrSPS1Fb-Chiifu401(Bra020096),這2個基因分別定位于LF和MF2亞基因組中,互為共線性基因。BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401均包含13個外顯子和12個內含子(圖1)。通過對大白菜材料He2進行重測序,分析BrSPS1Fa-He2和BrSPS1Fb-He2的變異,結果顯示,BrSPS1Fb-He2在6號內含子(I6)的剪接受體位點從AG突變為AC,而且此位點為純合的SNP變異,BrSPS1Fa-He2所有剪接受體位點未發生變異。
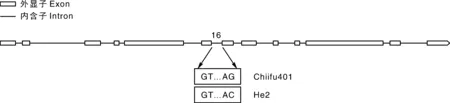
圖1 大白菜BrSPS1Fb-Chiifu401內含子-外顯子結構與大白菜He2 BrSPS1Fb剪接受體位點的SNP變異Fig.1 Intron-exon structure of BrSPS1Fb-Chiifu401 of Chinese cabbage and SNP variation of acceptor splicing site of BrSPS1Fb in the inbred line He2 of Chinese cabbage
2.2 大白菜He2的BrSPS1Fb剪接異構體鑒定與剪接方式
為鑒定大白菜BrSPS1Fb-He2 SNP變異后的剪接方式,選擇緊鄰6號內含子的PB序列(表1)作為檢索序列,在大白菜花瓣轉錄組測序文庫中進行檢索,獲得了4種類型剪接異構體,分別命名為S1~S4(圖2-A)。分析大白菜BrSPS1Fb-He2的剪接異構體序列,在大白菜He2BrSPS1Fb中鑒定了3個剪接受體位點(A1~A3),其中,A1和A2定位在BrSPS1Fb的I6內含子內部,而A3定位在外顯子E7上(圖2-B),剪接受體位點A1和A3均為AG,而A2則是非典型的AT。I6內含子中的GT(D1)是唯一的剪切位點供體。S1、S2、S3剪接異構體分別是選擇D1和A1、D1和A2、D1和A3進行剪接產生的。S4剪接異構體保留了整個I6內含子。

A,大白菜He2的BrSPS1Fb-He2基因4種剪接異構體及其相對應的剪接供體位點和剪接受體位點。B,BrSPS1Fb-He2基因的剪接供體位點和剪接受體位點的位置。黑色背景和灰色背景分別代表外顯子和內含子。A, Four splicing isoforms of BrSPS1Fb-He2 of Chinese cabbage He2 and their corresponding donor splicing sites and acceptor splicing sites. B, Location of donor splicing sites and acceptor splicing sites of the BrSPS1Fb-He2 gene. Black and gray background represented the sequence of exon and intron, respectively.圖2 BrSPS1Fb-He2基因的剪接異構體與剪接位點位置Fig.2 Splicing isoforms and location of splicing sites of BrSPS1Fb-He2
2.3 大白菜He2 BrSPS1Fa和BrSPS1Fb的剪接頻率
為分析BrSPS1Fb-He2剪接受體位點變異對其剪接的影響,分別使用PA1~PA5在大白菜He2的花瓣轉錄組測序read文庫中進行檢索。分析結果顯示,MF2亞基因組上的剪接受體位點未變異的BrSPS1Fa-He2(對照)只有S5類型的read(圖3),BrSPS1Fa-He2轉錄后可正常剪切I6內含子,未發生選擇性剪接。大白菜BrSPS1Fb-He2的I6剪接受體位點發生了變異,BrSPS1Fb-He2轉錄后不能選擇A4作為剪接受體進行剪接,不能產生正常剪接的S5類型剪接異構體。BrSPS1Fb-He2在剪接后可形成4種剪接異構體(S1~S4),其中,S3為主要的剪接產物,占BrSPS1Fb-He2所有轉錄本的81.73%。S1、S2、S4為次要剪接產物,分別占BrSPS1Fb-He2所有轉錄本數量的8.37%、3.40%和6.50%。
大白菜Chiifu401是白菜的參考基因組,其多個器官的轉錄組測序read文庫也可從NCBI中下載獲得。大白菜Chiifu401的BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401基因的剪接受體位點均未發生變異,分析這2個基因的選擇性剪接情況可進一步揭示剪接受體位點變異與選擇性剪接之間的關系。本研究從NCBI下載了大白菜Chiiffu401的花轉錄組測序read文庫(NCBI登錄號:SRR643622)。分別使用PA1~PA5和PB1~PB5在上述文庫中進行檢索,結果顯示,BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401在剪接過程中均選擇D1和A4進行剪接,只形成S5類型的剪接異構體。S1、S2、S3和S4類型的read均未檢測到(圖3)。大白菜Chiifu401的BrSPS1Fa-Chiifu401和BrSPS1Fb-Chiifu401在剪接過程中均正常,未出現選擇性剪接情況。
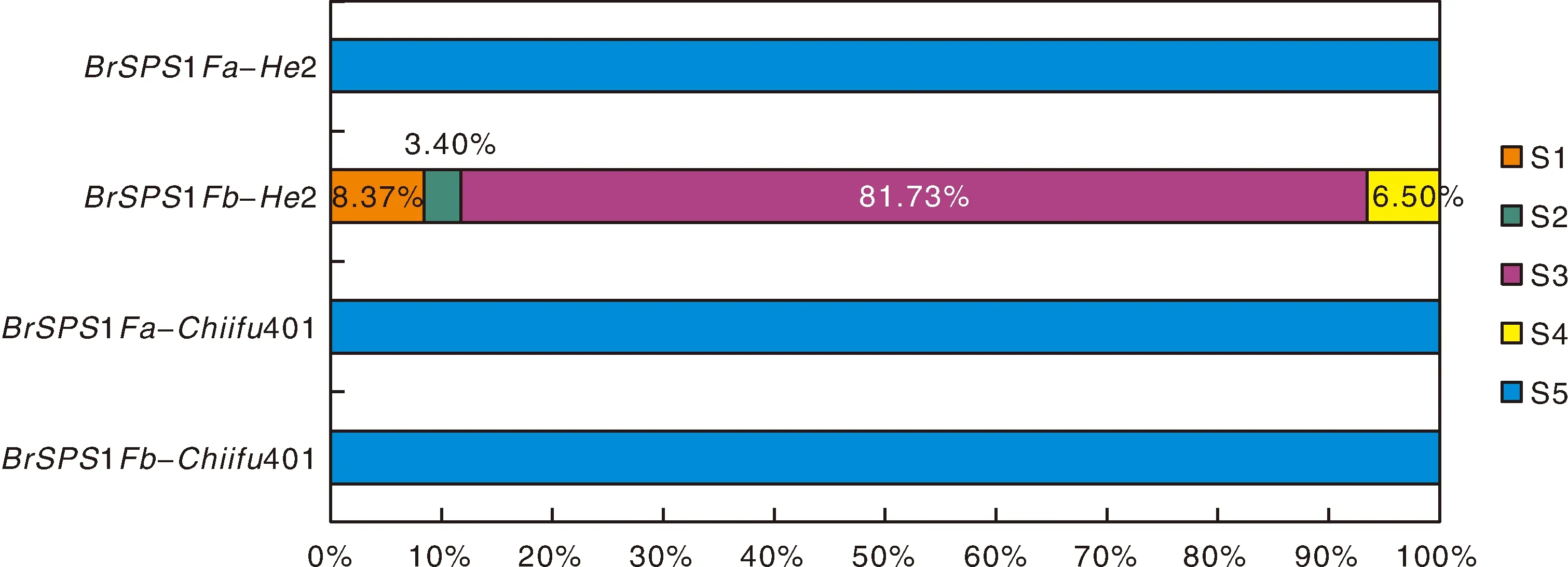
圖3 大白菜BrSPS1Fa和BrSPS1Fb基因的剪接異構體的比例Fig.3 Proportion of splicing isoforms of BrSPS1Fa and BrSPS1Fb of Chinese cabbage
2.4 大白菜He2 BrSPS1Fb轉錄本翻譯預測
由圖4可知,大白菜Chiifu-401的BrSPS1Fb基因只能形成1種I6完全剪切的S5類型轉錄本產物,其對應的蛋白質產物包含完整的E6和E7所對應的氨基酸序列。大白菜He2的BrSPS1Fb基因的剪接受體位點變異后,可主要形成S3類型的轉錄本,少部分形成S1、S2和S4類型的轉錄本,沒有檢測到S5類型的轉錄本。S1和S2類型的轉錄本中包含了I6部分內含子,對應的蛋白質產物分別插入了一段“SLLDLLIQHVMH”和“VMH”的氨基酸片段。S3類型轉錄本對應的蛋白質相對于BrSPS1Fb-Chiifu401缺少一段“IMRFFSNSRK”氨基酸序列。S4類型保留的整個I6內含子中包含1個提前終止密碼子,產生縮短蛋白。BrSPS1Fb-He2選擇性剪接會導致BrSPS1Fb-He2出現氨基酸片段的插入與缺少,甚至產生縮短蛋白。
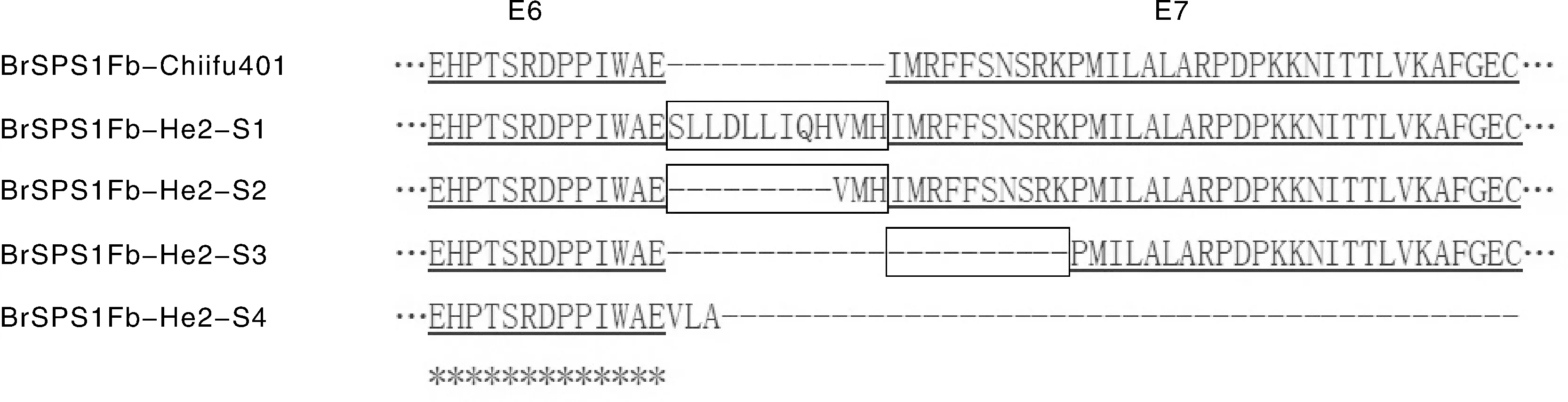
圖4 大白菜BrSPS1Fb-He2的蛋白產物與BrSPS1Fb-Chiifu401的氨基酸序列比對Fig.4 Alignment of amino acid sequences of protein products of BrSPS1Fb-He2 of Chinese cabbage and BrSPS1Fb-Chiifu401 protein
3 結論與討論
以大白菜He2的BrSPS1Fa和Chiifu401的BrSPS1Fa、BrSPS1Fb為參考序列,結果顯示,BrSPS1Fb-He2的I6剪接受體位點突變成AC之后,導致BrSPS1Fb-He2在pre-mRNA加工過程中I6發生選擇性剪接。白菜中FLC1的剪接供體位點由GT突變成AT后,導致其在pre-mRNA加工中發生了選擇性剪接,并產生了多種剪接異構體[6]。上述選擇性剪接是由剪接受體位點變異引起的。除了依賴剪接位點變異的選擇性剪接,薺菜和甘藍的KIN基因剪接位點均未發生突變,但它們在pre-mRNA加工中仍發生選擇性剪接[8-11]。選擇性剪接的類型很多,但由剪接位點SNP變異引起的選擇性剪接很可能會導致表型變異。
BrSPS1Fb-He2剪接受體位點變異成AC后,完全阻礙了I6選擇A4作為剪接受體進行剪切。因此,AC不能作為受體參與剪接過程,BrSPS1Fb-He2只能選擇其他潛在的剪接受體進行剪接(如A1、A2、A3)。A1~A4作為受體的剪接效率是不同的,未變異的A4受體(AG)剪接效率最高,其次是A3、A1、A2,以及變異的A4受體(AC)(圖3)。通常,真核生物基因內含子剪接過程中遵守GU-AG法則,并由剪接體介導進行[4,12-13]。本研究發現,BrSPS1Fb-He2在pre-mRNA加工中,AU(對應于A2位點)在某種程度上可被剪接體識別為剪接受體,以GU-AU法則進行剪接,但AU作為剪接受體進行剪接的效率不高(3.40%)。這種非典型的GU-AU剪接方式豐富了以GU-AG剪接方式為主的經典理論[14],拓寬了對基因剪接方式的新認識。
BrSPS1Fb-He2的選擇性剪接可產生4種剪接異構體,其中,S1和S2對應的蛋白分別插入了12個和3個氨基酸的片段,而S3則缺失了10個氨基酸的片段(圖4)。剪接受體位點變異后,BrSPS1Fb-He2的選擇性剪接導致出現了4種新蛋白。選擇性剪接廣泛存在于真核生物中,會造成同一個基因產生功能和結構上不同的mRNA和蛋白,或者由于影響基因表達水平而導致出現不同的表型[14-16]。BrSPS1Fb-He2基因的S4剪接異構體保留了整個I6內含子,導致在其I6中存在1個提前終止密碼子。在哺乳動物細胞中,如果1個mRNA提前終止密碼子距離外顯子-外顯子連接處上游超過50個核苷酸時,這個mRNA將經歷無義介導的mRNA降解(nonsense-mediated mRNA decay,NMD)[17]。BrSPS1Fb-He2基因S4類型轉錄本的提前終止密碼子距離外顯子-外顯子連接處上游遠遠超過50個核苷酸,暗示S4轉錄本將被NMD機制降解。BrSPS1Fb-He2編碼了一種蔗糖磷酸合成酶,擬南芥SPS1F突變體葉片中蔗糖磷酸合成酶的活性顯著降低,淀粉合成量增加。AtSPS1F在葉片光合蔗糖的合成中發揮重要作用[18-19],并促進植物生物量的積累[7]。大白菜He2材料具有生長勢小、生長緩慢的特點,這一性狀與生物量的積累密切相關,上述表型是否由BrSPS1Fb-He2剪接受體位點變異引起還需深入研究與探討。
