藥用植物鯊烯環氧酶基因研究進展△
2021-12-01 01:57:28李彤李劍超王晨曲雪潔戚文濤劉長利
中國現代中藥 2021年10期
李彤,李劍超,王晨,曲雪潔,戚文濤,劉長利
首都醫科大學 中醫藥學院,北京 100069
鯊烯環氧酶(squalene epoxidase,SE 或SQE)屬于單加氧酶,又稱鯊烯單加氧酶,廣泛存在于人參、雷公藤等多種藥用植物中。SE 催化角鯊烯發生環氧化反應生成2,3-氧化鯊烯,2,3-氧化鯊烯經過羥基化、糖基化、環化作用等修飾后得到三萜及植物甾醇,生物合成過程見圖1。SE 被認為是三萜皂苷、甾體皂苷等中藥活性成分生物合成途徑中重要的調控酶。近年來,次生代謝物質的生物合成途徑已有較為深入的研究,其中的關鍵酶基因受到廣泛關注,多種植物的SE基因被成功克隆挖掘,并進行功能研究[1]。本文對藥用植物鯊烯環氧酶基因的克隆提取材料、生物信息學分析、組織特異性、茉莉酸甲酯(MeJA)誘導調控、亞細胞定位、基因功能分析幾方面予以綜述,為藥用植物中鯊烯環氧酶基因的相關研究提供參考。
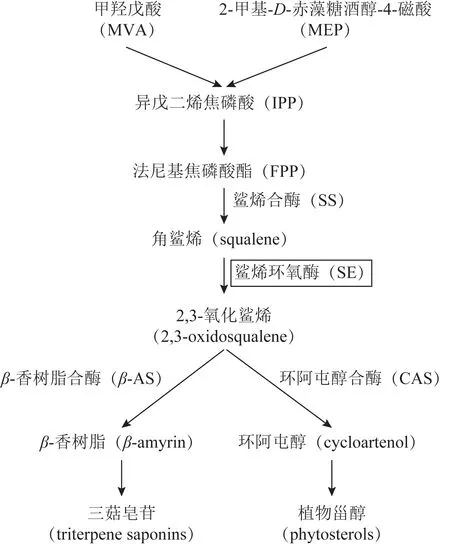
圖1 三萜皂苷與植物甾醇在植物體內的生物合成途徑
1 SE基因克隆提取材料與生物信息學分析
1.1 SE基因克隆提取材料
基因克隆的結果受提取材料影響。同一植物不同的組織器官,因其化學成分和含量的不同,克隆的難易與質量也會存在差異。通常,藥用部位是基因克隆首選的采樣部位。相關研究分別從滇重樓根、絞股藍葉片、羅漢果果實、桑黃、牛樟芝等菌類的菌絲克隆獲得相應的SE基因[2-7]。
另外,離體培養的植物細胞和器官同樣是理想的提取材料。Han等[8]從人參葉片中克隆獲得2條SE基因的全長cDNA 序列;祝傳書等[9]從雷公藤發狀根中克隆得到TwSE1、TwSE2;Liu 等[10]從雷公藤懸浮細胞中克隆得TwSQE6和TwSQE7;尤鵬升等[11]從刺五加葉片中克隆獲得刺五加SE 基因;Gao 等[12]從北柴胡毛狀根中克隆獲得BcSE1。這些部位作為采樣部位,與葉片、根組織等相比,易于提取和純化核糖核酸(RNA)。
1.2 SE基因生物信息學分析
生物體內,SE 常多于1 條[13],如擬南芥中已經克隆得到6條SE基因。SE氨基酸序列結構和理化性質有一定程度的差異與相似點,多具有跨膜結構域及功能結合域。編碼約500 個氨基酸,跨膜結構域數目0~6 個,具有黃素腺嘌呤二核苷酸(FAD)結合位點,多為堿性疏水蛋白[14]。表1是部分已報道的藥用植物SE基因克隆結果。

表1 藥用植物SE基因的生物信息學數據
N 端差異普遍存在于同一物種不同的SE 基因中。PgSE1與PgSE2具有83%同源性,N 端有很大的差異,前60 個氨基酸的同源性僅為23%[8]。龍月紅等[23]克隆獲得的2 條刺五加SE 基因cDNA,兩者間同源性高,N 端特異性較高。N 端差異可能是造成SE不同功能的原因之一。
2 SE基因組織特異性與MeJA誘導調控
SE 基因組織特異性主要分為2 種,即根中表達水平最高或葉片表達水平最高。綜合分析SE的組織表達特異性發現:人參[8]、北柴胡[12]、刺五加[23]、白樺[15]、藍莓[16]、盾葉薯蕷[17]、紫花苜蓿[18]的SE 基因在葉中表達量顯著高于根和莖;滇重樓[2]、雷公藤[10]、枸骨[19]、金鐵鎖[20]、遠志[21]、總序天冬[24]的SE基因在根中表達量顯著高于葉和莖;黃芪[25]和銀杏[26]的SE在莖中具有高表達量,橡膠草[22]的SE在乳膠中具有高表達量。SE 組織特異性可能與植物內部分泌組織在各個器官中的分布有關。人參在葉片中有豐富的樹脂道,可能是人參葉片SE表達量較高的原因。另外,對不同年份人參根中SE基因表達量分析發現[27],1~5 年生的人參中,5 年生人參根中表達水平最高,說明SE基因隨藥用植物生長年份累積表達水平提高。多數組織特異性分析僅選擇某一確切生長階段植物研究。然而,在植物不同生長時期分析各植物組織器官的SE表達量、皂苷含量,可以更清晰地闡明SE對皂苷合成、植物生長的影響,后續研究應予以關注。
MeJA是與損傷相關的植物激素和信號分子,可以激發植物防御基因表達。經MeJA 處理后,SE 基因表達普遍出現不同程度的提高。MeJA 處理人參后,PgSQE1在根中的表達增強,但PgSQE2在根中的表達降低[8]。MeJA 處理北柴胡不定根后,BcSE1與β-香樹脂合成酶基因(β-AS)的表達量一起上調,而環阿屯醇合酶基因(CAS)表達幾乎沒有變化[12]。β-AS與三萜類物質合成相關,CAS與甾醇合成相關[28],提示BcSE1可能是催化北柴胡三萜皂苷生物合成的關鍵酶。利用MeJA 處理刺五加后[23],SE1的表達水平提升顯著高于SE2。茉莉酸甲酯誘導后基因表達量提高可以為基因功能提供參考,但不足以確定某條SE的表達對皂苷生物合成的調控功能,SE基因在皂苷生物合成中的具體功能和參與的次生代謝途徑還需進一步分析。
3 SE基因的亞細胞定位
在過往研究中,鯊烯環氧酶被認為是一種膜結合蛋白,多定位在內質網膜(ER)上。部分文獻報道,SE 定位于非ER 上。研究者構建紫花苜蓿MsSQE瞬時表達載體,在洋蔥表皮細胞中表達,融合蛋白只在細胞膜上有微弱的熒光[18]。研究者將橡膠草的TkSQE1與藍色熒光蛋白Cerulean 分別在N 端及C 端融合[22],觀察到C 端TkSQE1 融合蛋白(TkSQE1-Cerulean)定位于ER,N 端TkSQE1 融合蛋白(Cerulean-TkSQE1)未觀察到定位。同樣地,構建瞬時表達載體,并在洋蔥內皮細胞中表達,三七SE主要位于在細胞膜及其周圍[29]。SE定位情況與軟件預測結果不同,可能是事實如此,也可能是實驗方案所限。基因的連接方式,即選擇N 端或C 端與熒光蛋白基因融合,很有可能影響蛋白的狀態。研究表明,SE 高簡并性的N 端編碼序列可能影響蛋白質的半衰期;C 端被認為與蛋白跨膜有關[30]。西葫蘆SE 研究中,eGFP 與CpSE1 進行N 端融合時,檢測不到eGFP-CpSE1 在ER 的熒光,與橡膠草N 端融合蛋白Cerulean-TkSQE1一樣未觀察到定位;而C端融合蛋白(CpSE1-eGFP)檢測到明顯的ER 定位[31],在N 端的熒光蛋白基因融合可能干擾了蛋白質的表達。亞細胞定位實驗中基因融合位置影響定位結果,實驗應將其考慮在內,以明確實驗中無熒光定位、熒光定位過于微弱的原因。
4 SE基因功能分析
4.1 SE對皂苷合成的影響
SE 的轉錄水平影響皂苷的合成。植物SE 對皂苷合成影響的研究中,常構建基因干擾載體或過表達載體等,并進行遺傳轉化,得到轉基因株系,從而對皂苷含量進行分析。
靈芝中SE 過表達可增加靈芝酸的合成,含量是野生型的2 倍[32]。劉天巍等[33]構建人參SE 基因RNA干擾(RNAi)載體,農桿菌介導轉化人參愈傷組織,轉化的人參愈傷組織中Rg1、Rc、Re單體皂苷含量都大幅下降。Han 等[8]構建PgSQE1-RNAi 的人參植株,轉基因組人參皂苷積累明顯減少,證明PgSQE1作用于人參皂苷生物合成。劉天巍與Han的研究均證明了SE 對人參皂苷生物合成具有調控作用。康俊梅等[18]在對紫花苜蓿的功能研究中,構建過表達MsSQE1轉基因植株,過表達MsSQE1苜蓿與對照植株相比MsSQE1表達水平上調,總皂苷含量也顯著增加。在西葫蘆研究中用農桿菌侵染法,獲得CpSE2轉基因毛狀根,過表達CpSE2的毛狀根與對照組相比,葫蘆素E 增加了1.7 倍,提示CpSE2過表達可增加三萜合成[31]。SE 遺傳轉化研究日益增多,對解析SE功能、皂苷生物合成途徑是非常有必要的。
4.2 SE對甾醇合成的影響
甾醇是植物體中不可缺且廣泛存在的一類物質,合成過程復雜。SE 普遍存在于植物中,催化甾醇的前體——環氧化鯊烯合成。萊茵綠藻的研究中,使用不同溶度的特比萘芬(1 種SE 抑制劑)處理綠藻細胞,甾醇及葉綠素含量顯著降低,特比萘芬劑量增大至150 mg·L-1,細胞漂白褪色,說明SE 在此種藻類甾體合成和細胞存活不可缺少;然而敲除萊茵綠藻CrSQE后,轉基因株系的甾醇含量沒有顯著降低[34]。在橡膠草研究中,研究者構建TkSQE1-RNAi轉基因植株,角鯊烯水平增加了27 倍,但2,3-氧化角鯊烯、甾醇和五環三萜的含量均未顯著降低[22]。有報道指出,酵母內SE表達存在反饋機制,當終產物耗盡時,SE 將再被合成[35]。亦有報道指出,1 種硅藻Phaeodactylum tricornutum缺乏SE 基因,其甾醇生物合成途徑嵌合了真菌與植物的甾體合成途徑[36]。殘留的SE 活性或基因調控可能使甾醇水平不受SE基因表達影響,也可能存在未知途徑進行催化反應,SE對甾醇合成的影響仍需要深入探究。
4.3 SE與其他酶之間的相互作用
SE 催化角鯊烯氧化過程受其他酶的影響。細胞色素P450 氧化還原酶(CPR)是1 種還原型煙酰胺腺嘌呤二核苷酸磷酸(NADPH)依賴的細胞色素P450 還原酶,有提供電子的作用,參與多種氧化還原反應。在對盾葉薯蕷研究中,研究者以角鯊烯為底物,進行體外酶促實驗,色譜圖中出現2,3-氧化鯊烯的峰;當反應物中無盾葉薯蕷細胞色素P450 氧化還原酶(DzCPR)或無CPR 蛋白時,即使增加角鯊烯,均無2,3-氧化鯊烯合成[17]。SE和CPR都直接且共同參與了2,3-氧化角鯊烯的催化反應,為SE功能研究展開了新視角,提示次生代謝產物合成中的電子轉移反應值得關注。
5 討論
基因表達是動態過程,存在著皂苷的轉移和植物不同生長階段帶來的變化。有學者提出,內質網負責植物油腺細胞中萜類化合物的運輸和(或)合成[37]。組織特異性分析中,SE 在葉片中常表現較高的轉錄水平,表明可能葉片是合成2,3-氧化角鯊烯的主要器官。有報道指出,根莖是皂苷積累的主要器官[38]。可以推測皂苷及其前體主要在葉片中合成,并轉移到根莖中儲存。而在橡膠草的SE組織特異性表達研究中,12 周和16 周齡植物中出現SE 表達高峰;短角蒲公英研究中也呈現在16 周多種酶基因表達高峰這一情況[39]。可能是由于膠質細胞的快速生長或根體積的快速擴張所致,這一時期植物快速生長對膜固醇的需求增加,SE基因表達量也會增加。
SE 表達與三萜合成之間呈正相關關系。另外,SE 與植物生長也有密切聯系,如SE 促進種子萌發[24],沉默SE 將影響根的伸長及生長[40]。隨著基因克隆技術日漸成熟,克隆基因的難度降低。確定基因是否對三萜的生物合成具有調控作用,進行三萜皂苷合成中SE的功能分析,常用基因沉默技術篩選出與三萜合成相關的基因。然而植物中SE為多基因家族,SE 基因催化作用存在多樣性。為提高三萜皂苷合成量,沉默非目標產物合成途徑上的基因可能對植株的正常生長造成影響,造成根系發育不良等情況,研究應予以關注。
隨著科技的發展和人類認識的進步,各個學科領域之間交融互進,分子生物學應用也越來越廣泛。深入了解次生代謝物質生物合成的各個步驟,解析有效成分次生代謝途徑,對緩解藥用植物資源緊張,促進藥用植物經濟的發展有重要作用。本文概述藥用植物SE 基因的研究進展,為其他藥用植物SE 基因的克隆與表達分析研究提供參考。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
