纖維素酶和木聚糖酶對象草青貯發酵品質及體外消化率的影響
2021-12-09 09:05:52吳永杰代童童尹雪敬賈玉山李君風
草地學報 2021年11期
丁 浩, 吳永杰, 邵 濤, 趙 杰, 代童童, 董 東, 尹雪敬, 賈玉山,李君風*
(1.南京農業大學草業學院飼草調制加工與貯藏研究所, 江蘇 南京 210095; 2.內蒙古農業大學農業農村部飼草栽培加工與 高效利用重點實驗室, 內蒙古 呼和浩特 010018; 3.內蒙古農業大學教育部草地資源重點實驗室, 內蒙古 呼和浩特 010018)
象草(PennisetumpurpureumSchumach)屬于禾本科狼尾草屬多年生草本植物,具有生物產量高、再生性強、生長速度快等優點,廣泛種植于熱帶、亞熱帶地區,是反芻動物重要的粗飼料來源[1]。熱帶、亞熱帶地區雨熱同期、高溫高濕,牧草很難調制成干草進行全年儲藏利用。而青貯不受氣候條件的限制,青貯飼料可全年長期有效地保存,調節飼草季節性供給不平衡[2-3]。
象草水溶性碳水化合物含量相對較低、粗纖維含量高,表現出多莖、多空、節間粗硬的物理特性。在青貯調制過程中難以壓實,殘余的空氣較多,造成植物呼吸作用及好氧性微生物活動時間延長[4]。好氧腐敗菌代謝易使水溶性碳水化合物大量損失,乳酸菌發酵底物不足,乳酸生成量少,pH很難快速降到4.2以下,往往導致青貯發酵品質不佳[5]。
酶制劑常用于改善青貯發酵品質[6]。添加纖維素酶能使纖維水解為葡萄糖,增加乳酸菌發酵底物;木聚糖酶能將木聚糖類半纖維素降解為低聚糖和木糖[7]。同時纖維素酶與木聚糖酶能打破青貯原料細胞壁表面結構,使纖維素網狀結構變得松散,比表面積增大。因此反芻動物采食酶制劑處理后的青貯飼料,更利于瘤胃微生物的附著與分解,提高飼料的消化率[8]。Desta等人[9]在象草青貯中添加木聚糖酶,發酵30 d后,青貯過程中水溶性碳水化合物和乳酸含量增加,半纖維素含量降低。李茂[10]報道王草青貯時添加纖維素酶,顯著提高了乳酸含量、降低了pH值、酸性洗滌纖維和中性洗滌纖維含量,提高了發酵品質。酶制劑添加劑在暖季型牧草青貯中研究較多,但在生產運用時仍面臨作用不穩定、普適性低等問題。本試驗以象草為材料,旨在探討纖維素酶、木聚糖酶單獨以及組合添加對象草青貯發酵品質和體外消化率的影響,為優質象草青貯飼料的生產和利用提供理論依據與技術支持。
1 材料與方法
1.1 試驗材料
供試象草種植于南京農業大學試驗地(32°01′19″ N,118°51′08″ E,海拔25 m),于2020年8月27日刈割,此時象草處于營養生長期,其化學成分如表1所示。試驗所用酶為青貯專用酶制劑,購于中國南京奧德美生物技術有限公司。纖維素酶、木聚糖酶的酶活為5 400 U·g-1,添加量參考使用說明。實驗室青貯窖采用容積為2 L有內外蓋的聚乙烯容器。
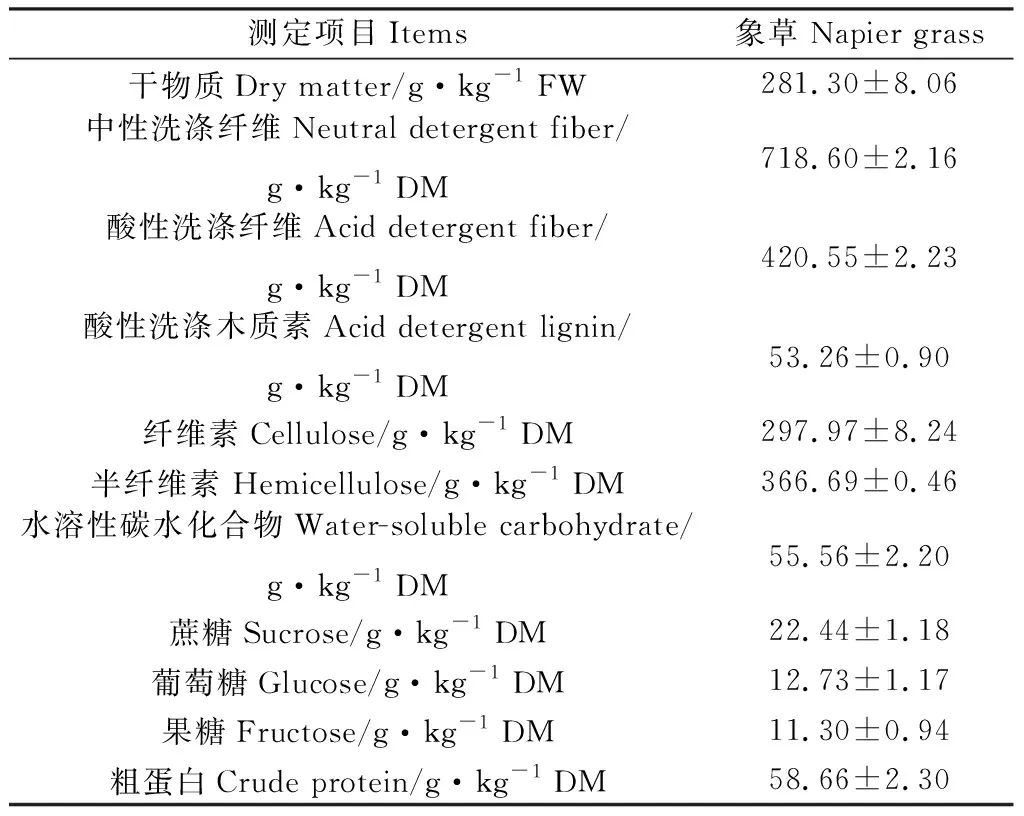
表1 象草的化學成分Table 1 The chemical compositions of napier grass
1.2 試驗設計
試驗采用完全隨機區組設計,設對照組(CK),添加纖維素酶組(CE),添加木聚糖酶組(XE),組合添加酶組(CX,纖維素酶+木聚糖酶)。添加量均以青貯原料鮮重(Fresh weight,FW)為基礎,其中纖維素酶、木聚糖酶添加量為0.3%(酶活為16.8 U·g-1FW),組合添加酶組添加量為0.15%纖維素酶+0.15%木聚糖酶。分別在青貯7,30,60,90 d后打開實驗室青貯窖取樣,測定相關發酵指標。每個處理每個青貯天數設3個重復,總共48個實驗室青貯窖。
1.3 試驗方法
1.3.1青貯飼料調制 象草切碎至2~3 cm,莖葉充分混合均勻后,按試驗設計添加量將溶解于滅菌水中的纖維素酶、木聚糖酶溶液噴灑在切碎的象草上,對照組加等量滅菌水,再次充分混勻,裝入2 L實驗室青貯窖中,每個青貯窖中裝1.36 kg,壓實后蓋上內外蓋,用膠帶密封,置于室溫(25℃±5℃)下保存。
1.3.2樣品處理 青貯窖打開后,將全部青貯飼料取出混勻,采用四分法精準稱量樣品20 g放入100 mL廣口錐形瓶中,加入60 g蒸餾水,密封后于4℃冰箱靜置24 h,然后通過4層紗布和定性濾紙過濾,得到青貯飼料的浸提液,立即測定pH后,于—20℃冰箱冷凍保存待測。浸提液用于測定氨態氮、乳酸、揮發性脂肪酸含量。將剩余青貯飼料收集,于65℃烘箱中烘60 h以上至恒重,粉碎過0.5 mm篩保存于自封袋中,用于水溶性碳水化合物、中性洗滌纖維、酸性洗滌纖維和體外消化率的測定。
1.3.3測定指標及分析方法 采用烘干恒重法測定青貯飼料干物質含量[11];pH值使用211型pH計(意大利哈納科技有限公司)測定[12];氨態氮含量采用苯酚-次氯酸鈉比色法測定[13];緩沖能采用鹽酸、氫氧化鈉滴定法測定[14];水溶性碳水化合物采用硫酸-蒽酮比色法測定[15];采用范氏纖維分析法測定青貯飼料中中性洗滌纖維(Neutral detergent fiber,NDF)、酸性洗滌纖維(Acid detergent fiber,ADF)和酸性洗滌木質素(Acid detergent lignin,ADL)含量,NDF測定時加入α淀粉酶、亞硫酸鈉[16]。
采用高效液相色譜儀(1260 HPLC;德國安捷倫有限公司)測定乳酸及揮發性脂肪酸含量,色譜柱為Carbomix?H-NP5,流動相為2.5 mmol·L-1H2SO4,流速設定0.5 mL·min-1,溫度設定55℃[14]。
采用高效液相色譜儀(1260 HPLC;德國安捷倫有限公司)測定青貯飼料中葡萄糖、果糖、蔗糖含量,色譜柱為Agilent InfinityLab Poroshell 120 HILIC-Z,流動相0.3%的乙腈氨水溶液,流速設定0.4 mL·min-1,溫度設定35℃[12]。
1.3.4體外消化率測定方法 瘤胃液取自4只體況相近,裝有永久性瘺管的去勢波爾雜交山羊(平均體重為35 kg)。飼喂水平及日糧組成為:苜蓿干草45 g,羊草干草350 g,混合精料240 g(10%麩皮+25%豆粕+65%玉米粉),維生素和礦物質。于晨飼前采集瘤胃液,四層紗布過濾,置于39℃保溫瓶中,并立即帶回試驗室,通入CO2維持厭氧條件。
稱取1 g青貯飼料樣品(65℃烘干后粉碎,過0.5 mm篩)于濾袋中,每個處理設3個重復,密封后置于血清瓶中,再向血清瓶中加入20 mL瘤胃液和40 mL McDougal緩沖液,置于搖床中培養(39℃,150 r·min-1),進行空白校正后,使用壓力傳感器技術記錄4,8,12,24,48,72 h的產氣量。試驗結束后,從血清瓶中取出濾袋,洗滌后在65℃烘箱中干燥48 h至恒重,測定DM,NDF和ADF殘留量,計算體外干物質消化率(InvitroDM digestibility,IVDMD),體外中性洗滌纖維消化率(InvitroNDF digestibility,IVNDFD)和體外酸性洗滌纖維消化率(InvitroADF digestibility,IVNDFD)[17]。利用GraphPad 6.01軟件,根據France產氣模型計算產氣動力學參數,模型:GPt=B×(1—exp(-c×(t—Lag)))[18],其中GPt是在t時間的累計產氣量,B表示理論最大產氣量(mL);c表示產氣速率常數(mL·h-1),Lag表示體外發酵產氣延滯時間(h),t表示產氣時間點(h)。
動力有限元模型的離散參數需要根據相應地層的剪切波波速及激勵的頻率范圍來確定[16]。為了簡化模型,同時又能反映地層的動力特性,根據地層剪切波速的變化范圍,將復雜的真實地層簡化為5層水平構造,每層代表一個主要的剪切波速域[17]。剪切波速cs的計算公式[18]為:
1.4 數據處理與分析
采用Excel 2019進行基礎數據收集歸納。采用統計分析系統(SAS 8.0)GLM程序對數據進行雙因素方差分析(ANVOA),采用Tukey’s HSD方法對處理間和青貯天數間平均數進行多重比較(P<0.05)。
2 結果與分析
2.1 添加酶制劑對象草青貯過程中pH、干物質損失、干物質含量和氨態氮的影響
如表2所示,酶制劑處理、青貯天數及其互作對青貯過程中pH、干物質損失、干物質含量和氨態氮有顯著影響(P<0.05)。整個青貯過程中,各組pH逐漸降低,30 d降至最低,隨后略有提高。CE,XE,CX組pH低于CK組;青貯30 d后CX組pH顯著低于其它三組(P<0.05),CE與XE組無顯著差異。
青貯過程中,干物質含量逐漸降低,青貯前7 d各組差異不顯著,之后CE,XE,CX組干物質含量顯著高于CK組(P<0.05);CE,XE組青貯60 d,90 d后干物質含量顯著高于CX組(P<0.05),CE和XE組無顯著差異。氨態氮含量隨著青貯的進行顯著提高(P<0.05),CE,XE,CX低于CK組;青貯30 d后CX組氨態氮含量顯著低于CE和XE組(P<0.05),CE和XE組無顯著差異。
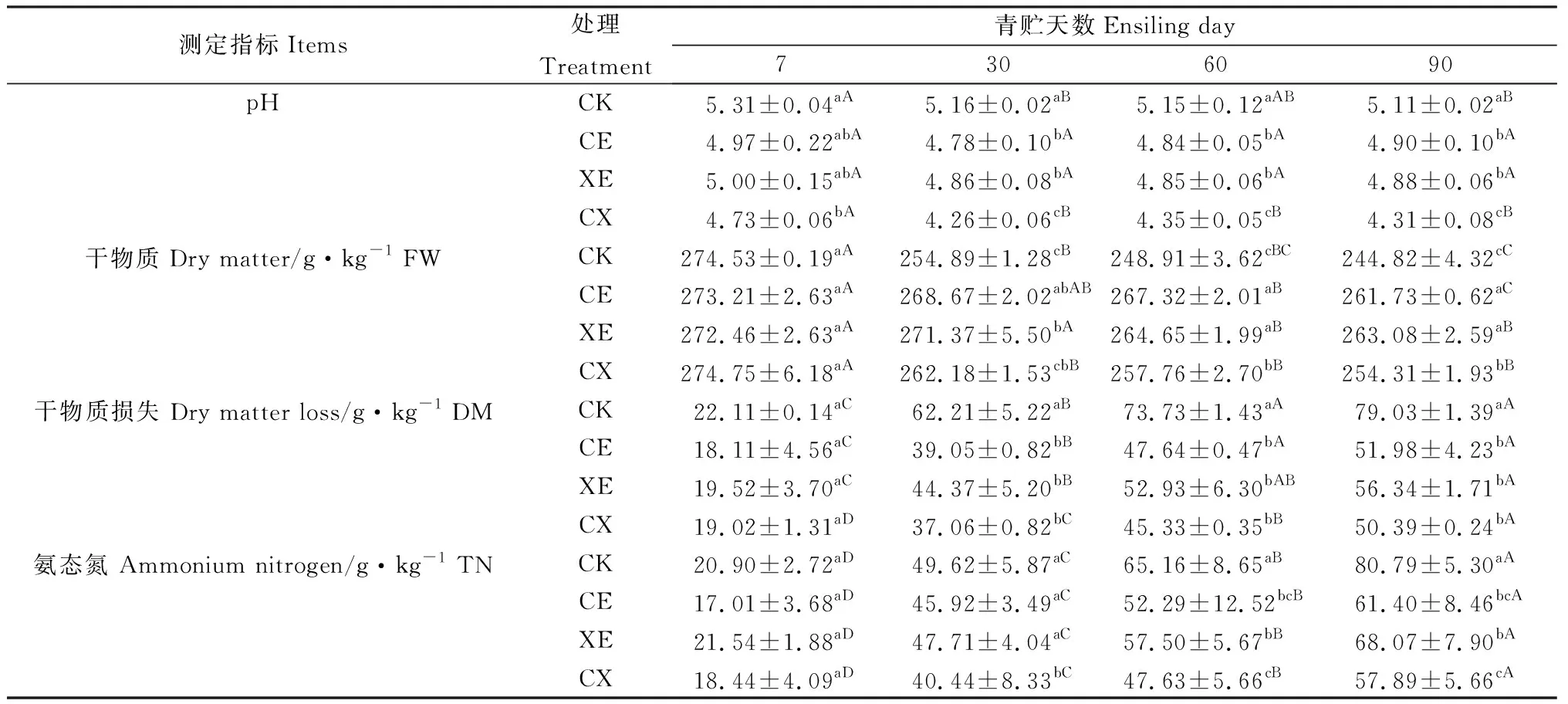
表2 添加酶制劑對象草青貯過程中pH、干物質損失、干物質和氨態氮含量的影響Table 2 Effects of enzymes addition on pH,DM loss,AN and DM content of napier grass during ensiling
2.2 添加酶制劑對象草青貯過程中有機酸含量的影響
如表3所示:酶制劑處理、青貯天數及交互作用對青貯過程中乳酸、乙酸、丙酸和丁酸含量有顯著影響(P<0.05)。青貯過程中乳酸含量逐漸提高,青貯第30 d達到峰值,之后逐漸降低。CE,XE,CX組乳酸含量在21.28~48.44 g·kg-1DM之間,顯著高于對照組(P<0.05);CX組乳酸含量始終高于CE,XE組。CE與XE組間青貯前期乳酸含量無顯著差異,但青貯60 d,90 d后CE組乳酸含量顯著高于XE組(P<0.05)。
隨著青貯天數的延長,乙酸含量逐漸提高。CE,XE,CX組乙酸含量始終低于CK組。青貯過程中乳酸/乙酸下降,但CE,XE,CX組均高于CK組(P<0.05);除青貯第7 d以外,CX組乳酸/乙酸均顯著高于CE,XE組(P<0.05)。青貯過程中各組丙酸含量逐漸提高,除對照組青貯30 d后檢測到少量丙酸(3.49~4.85 g·kg-1DM)外,其余處理組均僅檢測出微量丙酸(0.38~1.86 g·kg-1DM)。
整個青貯過程中,各組丁酸含量有所提高,但含量均較低。青貯30 d后CE,XE,CX組檢測到少量的丁酸(2.95~6.17 g·kg-1DM),顯著低于CK組(7.11~13.02 g·kg-1DM,P<0.05)。
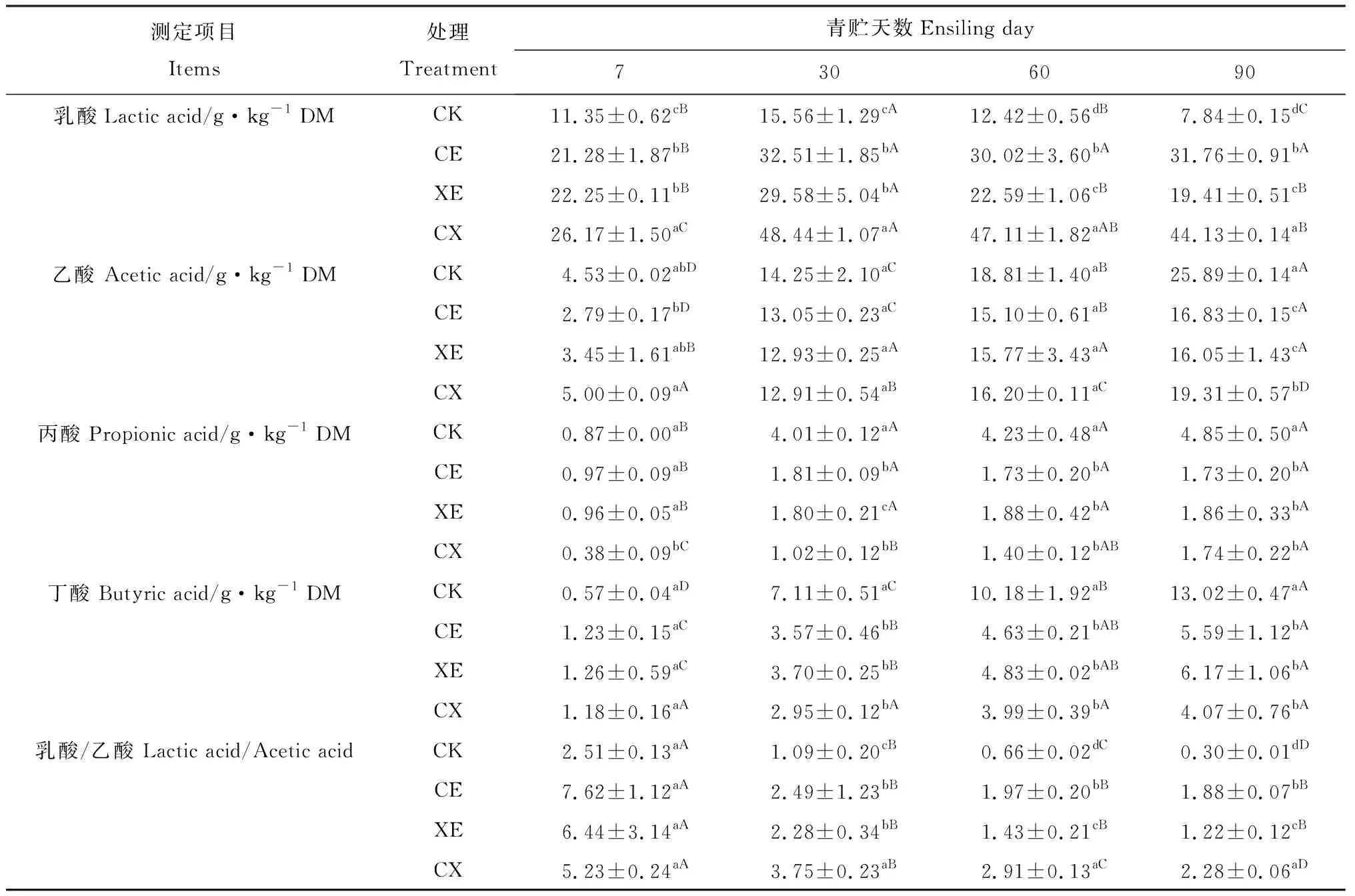
表3 添加酶制劑對象草青貯過程中有機酸含量的影響Table 3 Effects of enzymes addition on organic acid content of napier grass during ensiling
2.3 添加酶制劑對象草青貯過程中結構性碳水化合物組分含量的影響
如表4所示,酶制劑處理、青貯天數及交互作用顯著影響青貯過程中除ADL以外所有結構性碳水化合物組分含量(P<0.05)。青貯過程中各組NDF含量逐漸降低,CE,XE,CX顯著低于CK組(P<0.05);CX組NDF含量最低,在青貯第30 d與90 d顯著低于CE,XE組(P<0.05);CE與XE組除青貯第30 d外無顯著差異。
青貯過程中添加酶處理組ADF含量逐漸降低,且均低于CK組。從青貯第30 d起,CE與CX組ADF含量顯著低于XE組(P<0.05),CE與CX組無顯著差異。整個青貯過程中,各組ADL含量呈略有提高的趨勢,CE,XE,CX組ADL含量始終高于CK組,但各酶添加組差異均不顯著。
隨著青貯天數的延長,纖維素、半纖維素含量逐漸降低。CE,XE,CX組纖維素含量低于CK組,青貯30 d后CE,CX組顯著低于XE組(P<0.05),CE,CX組無顯著差異。CE,XE,CX組半纖維素含量低于CK組,CX組半纖維素含量最低,在青貯30 d與60 d時顯著低于CE,XE組(P<0.05),XE組半纖維素含量低于CE組。
2.4 添加酶制劑對象草青貯過程中水溶性碳水化合物、葡萄糖、果糖和蔗糖含量的影響
如圖1所示,整個青貯過程中,水溶性碳水化合物含量持續降低,CE,XE,CX水溶性碳水化合物含量高于CK組,其中CX組最高,CE與XE無顯著差異。青貯過程中,各組葡萄糖含量逐漸降低,CE,XE,CX組葡萄糖含量高于CK組,CX組含量最高,且在青貯第7 d,30 d顯著高于CE,XE組(P<0.05)。隨著青貯時間的延長,蔗糖、果糖含量逐漸降低,添加酶處理組始終顯著高于CK組(P<0.05),CX組蔗糖、果糖含量最高,CE與XE組無顯著差異。
2.5 添加酶制劑對象草青貯飼料體外消化率、體外產氣量的影響
如表5所示,整個體外發酵過程中,CE,XE,CX組48,72 h累計產氣量均高于CK組,CX組48,72 h累計產氣量顯著高于CE,XE組(P<0.05),CE,XE組無顯著差異。添加酶制劑處理后的象草青貯飼料IVDMD在57.09%~61.30%之間,顯著高于對照組(48.03%,P<0.05),CX組IVDMD顯著高于CE,XE組(P<0.05),CE和XE組無顯著差異。添加酶制劑象草青貯IVNDFD提高了4.33%~8.98%,IVADFD提高了7.83%~12.46%,均顯著高于對照組(P<0.05)。
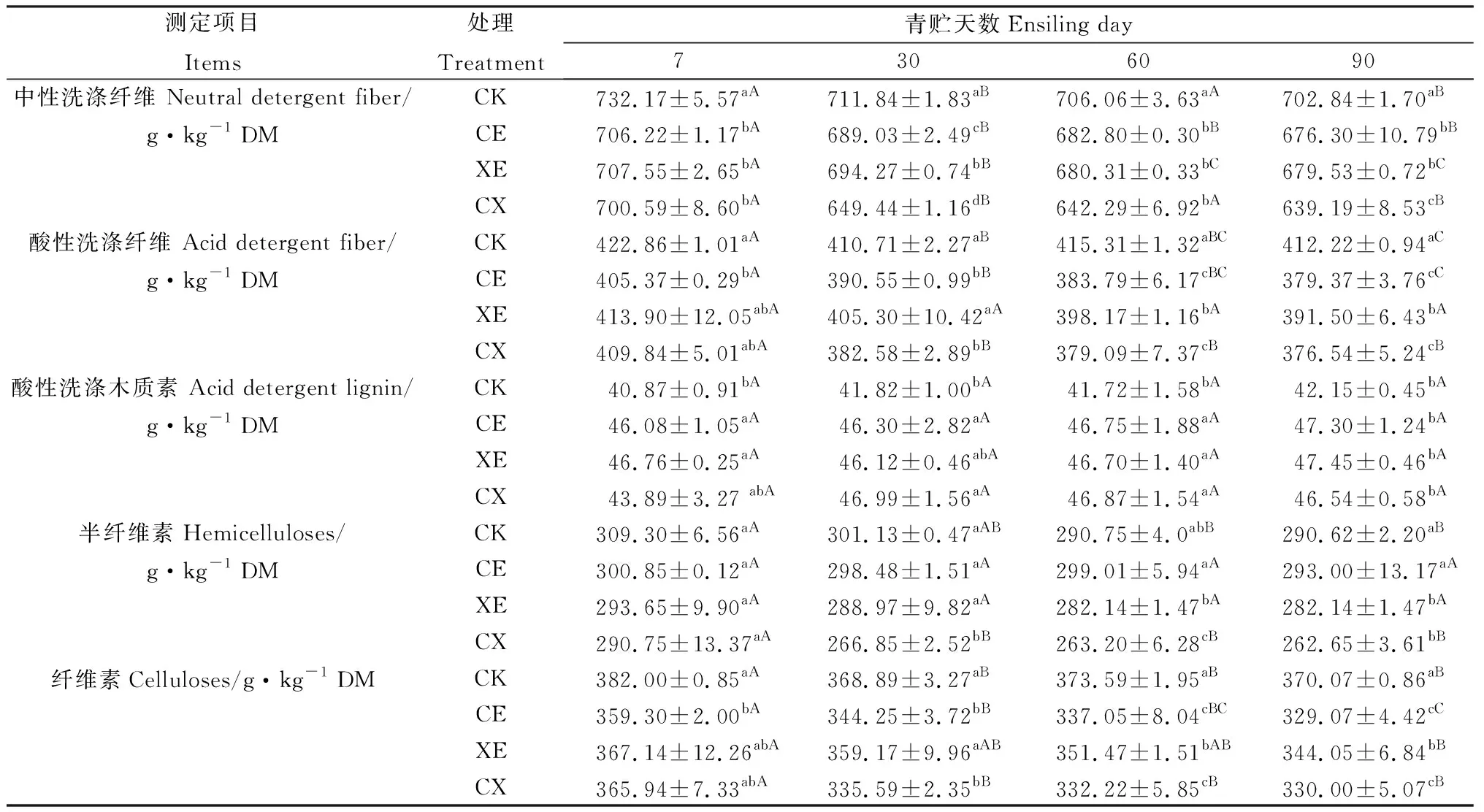
表4 添加酶制劑對象草青貯過程中結構性碳水化合物組分含量的影響Table 4 Effects of enzymes addition on structural carbohydrate components of napier grass during ensiling

圖1 酶制劑對象草青貯過程中水溶性碳水化合物、葡萄糖、果糖、蔗糖含量的影響Fig.1 Effects of enzymes addition on water soluble carbohydrate,glucose,fructose,sucrose contents of napier grass during ensiling注:不同小寫字母表示同一時間內不同處理差異顯著(P<0.05)Note:Different lowcase letter indiacte significant differences at the 0.05 level
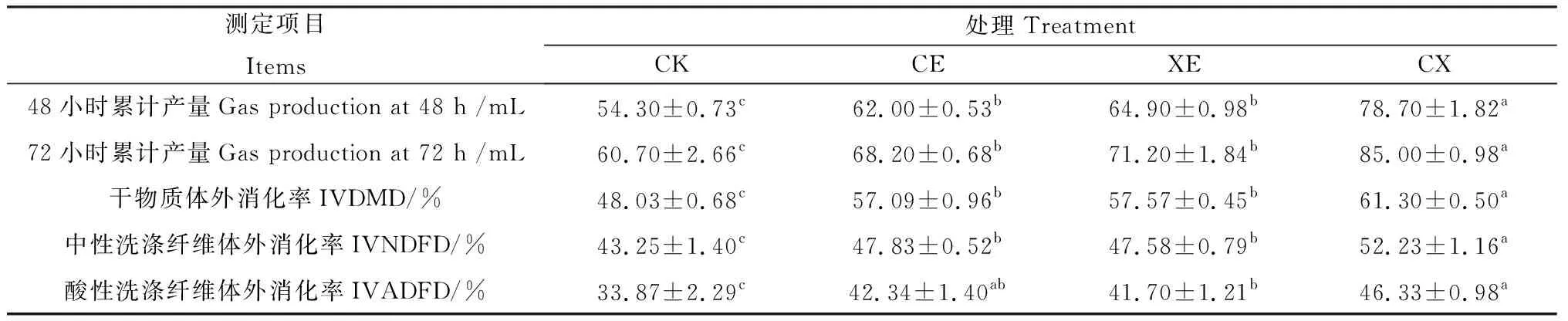
表5 酶制劑對青貯90 d象草青貯飼料體外消化率和體外產氣量的影響Table 5 In vitro digestibility coefficient and gas production of napier grass silage treated with enzymes after 90 days of ensiling
3 討論
3.1 添加酶制劑對象草青貯過程中發酵品質的影響
青貯過程中CE,XE及CX組干物質損失在18.11~56.34 g·kg-1DM之間,低于對照組22.11~79.03 g·kg-1DM,表明添加酶制劑組減少了干物質的損失。干物質損失主要是青貯早期的植物呼吸作用及青貯中、后期的微生物代謝所致[19]。青貯7 d后,酶制劑組乳酸含量顯著高于對照組,表明在象草青貯中添加酶制劑促進了粗纖維的降解,釋放水溶性碳水化合物,為乳酸菌提供了額外的發酵底物,快速產生乳酸,降低pH,抑制了不良微生物的活性,這與酶制劑組WSC含量顯著高于對照組相呼應[20]。CE,XE及CX組在青貯30 d后pH值顯著低于對照,CX組pH降至4.21,乳酸含量達48.84 g·kg-1DM,高于CE,XE組,說明在青貯中組合添加酶發酵品質優于單一添加。本研究與Smerjai等[21]研究結果一致。
通常青貯飼料中乳酸/乙酸大于2時,以同型乳酸發酵為主[22]。本試驗中青貯前期乳酸含量提高,pH下降,以同型乳酸發酵為主,隨著青貯的進行耐酸的異型發酵乳酸菌逐漸替代同型發酵乳酸菌成為優勢菌群,既能產生乙酸,也能利用乳酸異化產生乙酸,導致乙酸含量提高,乳酸/乙酸下降[23-25]。
Tao等人[26]報道優質青貯飼料氨態氮含量應小于100 g·kg-1TN,本試驗氨態氮含量最高為80 g·kg-1TN(< 100 g·kg-1TN),整個青貯過程中酶制劑組氨態氮含量低于對照組,青貯60 d顯著低于對照組,這是由于酶添加組乳酸含量高、pH低,抑制了不良微生物的活性,減少了對蛋白質的降解,使氨態氮含量降低。Li等[27]在狼尾草(Pennisetumsinese)青貯中添加纖維素酶檢測出微量的丙酸、丁酸,與本試驗結果一致,表明在象草青貯中添加酶制劑能有效抑制有害微生物活性,減少了丁酸和丙酸的產生[3,14]。
3.2 添加酶制劑對象草青貯過程中碳水化合物的影響
青貯過程中添加酶制劑,有助于植物細胞壁成分的降解,使纖維素、半纖維素含量降低,水溶性碳水化合物含量提高[18]。本試驗添加酶制劑、青貯天數及兩者的交互作用顯著降低了象草青貯過程中除ADL以外所有結構性碳水化合物組分的含量。青貯過程中CE,XE及CX組的纖維素、半纖維素含量均低于對照組,葡萄糖、果糖、蔗糖及水溶性碳水化合物含量高于對照組,認為酶解作用直接引起了纖維素、半纖維素的降解,導致水溶性碳水化合物含量的提高[9]。寇江濤等[28]報道在皇竹草中添加木聚糖酶,顯著降低了ADF含量。不同種類的酶制劑之間具有協同降解粗纖維的作用,組合添加更有利于青貯早期將結構性碳水化合物降解為水溶性碳水化合物,本試驗組合添加酶制劑組中纖維素酶與木聚糖酶協同作用導致纖維素、半纖維素含量顯著低于單獨添加纖維素酶、木聚糖酶[12],這與組合添加酶組具有較高的WSC含量相呼應。
Shao等[29]研究表明葡萄糖和果糖是乳酸菌優先利用的底物,青貯開始后乳酸菌增殖,利用葡萄糖和果糖,產生乳酸,乳酸含量逐漸提高,葡萄糖和果糖含量逐漸降低,青貯后期趨于穩定。本試驗整個青貯過程中,CE,XE及CX組水溶性碳水化合物、葡萄糖、果糖和蔗糖含量均高于對照組,表明纖維素酶和木聚糖酶降解了部分纖維素和半纖維素,釋放了葡萄糖、果糖和蔗糖,提高了水溶性碳水化物含量;其次添加纖維素酶和木聚糖酶加速了青貯過程中乳酸的生成,抑制了有害微生物對水溶性碳水化合物的消耗,使更多的WSC得以保留。黃秋連[30]和趙金鵬等[6]在羊草和水稻秸稈青貯中也得到相似結果。CX組WSC含量始終高于CE,XE組,纖維素和半纖維素含量顯著低于CE,XE組,說明纖維素酶和木聚糖酶組合添加更有利于提高象草青貯發酵品質。
3.3 酶制劑對象草青貯飼料體外消化率的影響
體外模擬瘤胃發酵在一定程度上可以反映反芻動物對青貯飼料的消化率,IVDMD,IVNDFD,IVADFD和體外產氣量是重要的衡量指標[31]。研究表明青貯飼料IVDMD與粗纖維含量負相關[20]。與對照組相比,酶制劑組青貯90 d后的青貯飼料IVDMD顯著高于對照組,這與Chen等[32]研究結果一致。Li等[27]報道酶降解作用可以改變青貯飼料表面結構,增加纖維間的孔隙度、表面積和可及性,增加反芻動物瘤胃微生物酶與青貯飼料的接觸面積,提高IVNDFD和IVADFD。本試驗添加酶制劑組與對照相比IVNDFD提高了4.33%~8.98%,IVADFD提高了7.83%~12.46%,說明添加酶制劑組較低的ADF,NDF有利于瘤胃微生物對粗纖維的降解。CX組IVNDFD,IVADFD高于CE,XE組,這是由于纖維素酶與木聚糖酶之間的協同作用,不僅提高了青貯飼料的WSC、乳酸含量為瘤胃微生物提供了發酵底物,也裂解了象草中纖維素與半纖維素之間的連接鍵,使纖維素結晶度降低,比表面積增大,利于瘤胃微生物附著及降解[9]。
青貯飼料的發酵產物、碳水化合物和含氮物質是體外產氣的主要底物[33-34]。體外產氣速率越快,累計產氣量越多,飼料消化率越高。本試驗添加酶制劑組72 h累計產氣量、產氣速率常數和理論最大產氣量均顯著高于對照組,表明添加酶制劑可提高青貯飼料的消化率。IVNDFD,IVADFD的提高增加了飼料比表面積,利于瘤胃微生物的附著與增殖[35]。CX組72 h累計產氣量顯著高于CE,XE組,也證明了組合添加酶組青貯飼料消化率更高。
4 結論
綜上所述,添加纖維素酶、木聚糖酶降低了纖維素、半纖維素含量,提高了葡萄糖、果糖、蔗糖及總水溶性碳水化合物含量,促進了青貯過程中乳酸發酵,快速產生乳酸、降低pH、氨態氮含量,提高了發酵品質、體外消化率和體外產氣量。但纖維素酶和木聚糖酶組合添加效果最好。
