鹽度、溫度和光照強度對針葉蕨藻的生長及光合活性的影響
2021-12-14 03:33:10鐘逸云楊蘊琪郜曉峰邢浩劉煒段元亮何文輝何培民
熱帶亞熱帶植物學報 2021年6期
鐘逸云, 楊蘊琪, 郜曉峰, 邢浩, 劉煒, 段元亮,2, 何文輝,3, 何培民,3
鹽度、溫度和光照強度對針葉蕨藻的生長及光合活性的影響
鐘逸云1a,1b, 楊蘊琪1a, 郜曉峰1a,1b, 邢浩1a,1b, 劉煒1a,1b, 段元亮1a,1b,2, 何文輝1a,3, 何培民1a,1b,3
(1. 上海海洋大學, a. 海洋生態與環境學院; b. 海洋科學研究院, 上海201306; 2. 四川省水產研究所, 成都 611731; 3. 水產科學國家級實驗教學示范中心(上海海洋大學), 上海 201306)
為探究環境因子對針葉蕨藻()生長的影響,對不同鹽度、溫度和光照強度下針葉蕨藻的生長和葉綠素熒光參數進行了研究。結果表明:藻體日特定生長率(SGR)、最大光量子產量(Fv/Fm)、實際光合效率(Yield)、電子傳遞速率(ETR)和光化學淬滅(qP)隨鹽度升高呈先上升后下降的變化趨勢,非光化學淬滅(qN)則呈相反的變化趨勢,藻體光合活性和固碳效率在鹽度27.5‰時達到最高,且與25‰和30‰鹽度的差異顯著(<0.05,=3)。藻體SGR、Fv/Fm、Yield、ETR和qP隨溫度升高而下降,qN則相反,藻體光合活性和固碳效率在26℃下達到最高,且與28℃和30℃的差異顯著(<0.05,=3)。藻體的SGR、Fv/Fm、Yield、ETR和qP隨光照強度升高呈先上升后下降的變化趨勢,qN則相反,且在18.75mol/(m2·s)弱光照下出現輕微光抑制,藻體生長、光合活性及固碳效率在光照強度25.00mol/(m2·s)時達到最高,但與18.75和31.25mol/(m2·s)的差異不顯著(>0.05,=3)。因此,針葉蕨藻在27.5‰鹽度、26℃和25.00mol/(m2·s)光照強度下生長最快且光合作用能力最高。
針葉蕨藻;葉綠素熒光參數;特定生長率;鹽度;溫度;光照強度
大型海藻是海洋生態系統中的初級生產者,通過吸收氮磷等營養物質降低富營養化程度,并抑制赤潮發生,常被應用于海域生態修復及水產養殖等,因而在生物地球化學循環過程中發揮了重要作用[1–2]。當前我國水族產業蓬勃發展[3],一些具有觀賞價值的大型海藻對大、中、小型海洋生態水族箱具有創建生境、凈化水質的效果。目前海洋生態水族箱常配置的觀賞大型海藻主要有針葉蕨藻()、杉葉蕨藻()、長莖葡萄蕨藻()、石莼()等,其中,針葉蕨藻最為常見,這些海藻具有較強的吸收營養鹽、釋放氧氣等生態功能,同時又具有很好的觀賞價值。
針葉蕨藻隸屬于綠藻門(Chlorophyta)羽藻綱(Bryopsidophyceae)蕨藻科(Caulerpaceae)蕨藻屬,藻體羽毛狀,十分美麗,俗稱羽毛藻,在我國大陸主要分布于廣東和海南沿海近岸,極具觀賞價值,且可食用,也成為南海近海養殖的一種名貴經濟大型海藻。其生長速度快,環境適應能力較強,且具有一定的凈化水質功能[4]。針葉蕨藻在鹽度20‰~ 40‰下均能生長[5],Mosquera-Murillo等[6]報道在鹽度15‰的水體中藻體逐漸發白;袁榮榮等[7]的研究表明,針葉蕨藻在25℃~30℃時生長良好,適宜的光照強度為25.00mol/(m2·s)。在相同環境條件下,針葉蕨藻的凈化水質效果優于長莖葡萄蕨藻和細齒麒麟菜()[8]。
針對針葉蕨藻的生長和營養鹽代謝能力等方面開展了一些研究[8],但未見針對其在不同環境條件下光合活性的研究報道。基于針葉蕨藻的觀賞性被廣泛應用于海水觀賞水族箱中,本研究結合常見觀賞魚類如眼斑雙鋸魚()等的適宜生存條件[9],探索針葉蕨藻在不同環境條件下的生長和光合活性的生理響應,為今后針葉蕨藻進一步開發利用奠定基礎。
1 材料和方法
1.1 藻體來源和暫養條件
針葉蕨藻()采集于南海文昌近岸海區。采集后采用航空低溫運輸運回實驗室,然后用消毒海水進行清洗,去除藻體上的雜藻和附著物。藻體培養在25‰鹽度、25℃和光照強度25.00mol/(m2·s)下備用。經過7 d復蘇培養的針葉蕨藻生長旺盛(圖1)。

圖1 水族箱復蘇培養的針葉蕨藻
1.2 培養條件和培養裝置
針葉蕨藻復蘇培養后,選取生長旺盛的藻體剪取鮮綠分枝末梢,長度為(10±0.50) cm、鮮質量為(1±0.50) g,放入盛有1 L人工海水的定制圓形玻璃容器中進行充氣懸浮培養,并加入適量霍格蘭(Hoa- gland)培養液。培養容器放置在光照培養箱(HGZ- 400N, 上海慧泰)中,光照周期為光照∶黑暗=12∶12,設置不同的海水鹽度、溫度、光照強度進行試驗。試驗周期為15 d,每5 d更換1次培養液。人工海水采用海水晶(藍鴻星,海鼎科技)配制及使用光學鹽度計檢測并確定鹽度,過濾消毒后備用。
試驗設置鹽度、溫度、光照強度3個環境因子,每個因子設3個水平,每個試驗設3組平行。鹽度分別設置25‰、27.5‰和30‰;溫度分別設置26℃、28℃和30℃;光照強度分別設置18.75、25.00和31.25mol/(m2·s)。某因子試驗時其余環境條件分別設定為鹽度27.5‰、溫度26℃和光照強度25.00mol/(m2·s)。
1.3 藻體生長和特定生長率測定
每隔5 d采集1次藻體,用吸水紙吸去表面多余水分后稱量鮮質量,并計算藻體日特定生長率(SGR)。SGR (%)=(ln1-ln0)/×100%, 式中,1為本次收集的藻體生物量;0為上次收集的藻體生物量;為間隔時間。
1.4 藻體葉綠素熒光參數測定
針葉蕨藻培養15 d后,采用葉綠素熒光儀(Dual- PAM-100,Walz Germany)測定相關參數。藻體樣品經15 min暗處理后測量,得到最小熒光值(Fo)、最大熒光值(Fm)、穩態熒光值(Ft)、最大熒光值(F?m)和最小熒光值(F?o)。計算藻體PS II最大光化學效率(Fv/Fm)實際量子產量(Yield)、電子傳遞速率(ETR)、光化學淬滅(qP)和非光化學淬滅(qN)。Fv/Fm=(Fm- Fo)/Fm, Yield=F/F?m=(F?m-Ft)/F?m, ETR=PAR×Yield×0.84×0.5, qP=1-(F-F?o)/(F?m-F?o), qN=1-(F?m-F?o)/(Fm-Fo)。
1.5 數據的統計和分析
所有數據使用GraphPad Prism 7.04處理并繪圖,使用SPSS 23.0進行單因素方差分析。實驗數據以平均數±標準偏差表示,以<0.05表示差異顯著。
2 結果和分析
2.1 對環境因子的響應
鹽度 從圖2可見,在鹽度27.5‰下培養15 d,藻體平均鮮質量提高了1.69 g,且SGR均保持在較高水平,達(6.35±0.67)%,顯著高于其余鹽度處理(<0.05,=3)。在鹽度25‰和30‰下培養15 d, 藻體平均鮮質量僅提高了0.91 g,且0~5 d藻體的SGR只有1.88%,顯著低于27.5‰鹽度(<0.05,=3), 0~ 10 d的SGR提高到5.10%~5.24%,0~15 d的SGR僅為4.06%~4.43%, 說明針葉蕨藻在鹽度27.5‰下生長最好。
溫度 從圖3可見,在溫度26℃下培養15 d,藻體平均鮮質量提高了2.10 g, SGR高達(7.11±0.13)%,顯著高于其余溫度處理(<0.05,=3)。在溫度28℃和30℃下培養15 d,藻體平均鮮質量僅提高2.22~ 2.68g,0~5 d的SGR為5.80%,0~10 d的SGR下降,為4.73%~5.54%,0~15 d的SGR僅為4.10%~ 4.72%, 說明針葉蕨藻在溫度26℃下生長最快。

圖2 不同鹽度下針葉蕨藻的生長和日特定生長率(SGR)。相同培養天數柱上不同字母表示差異顯著(P<0.05, n=3)。下圖同。
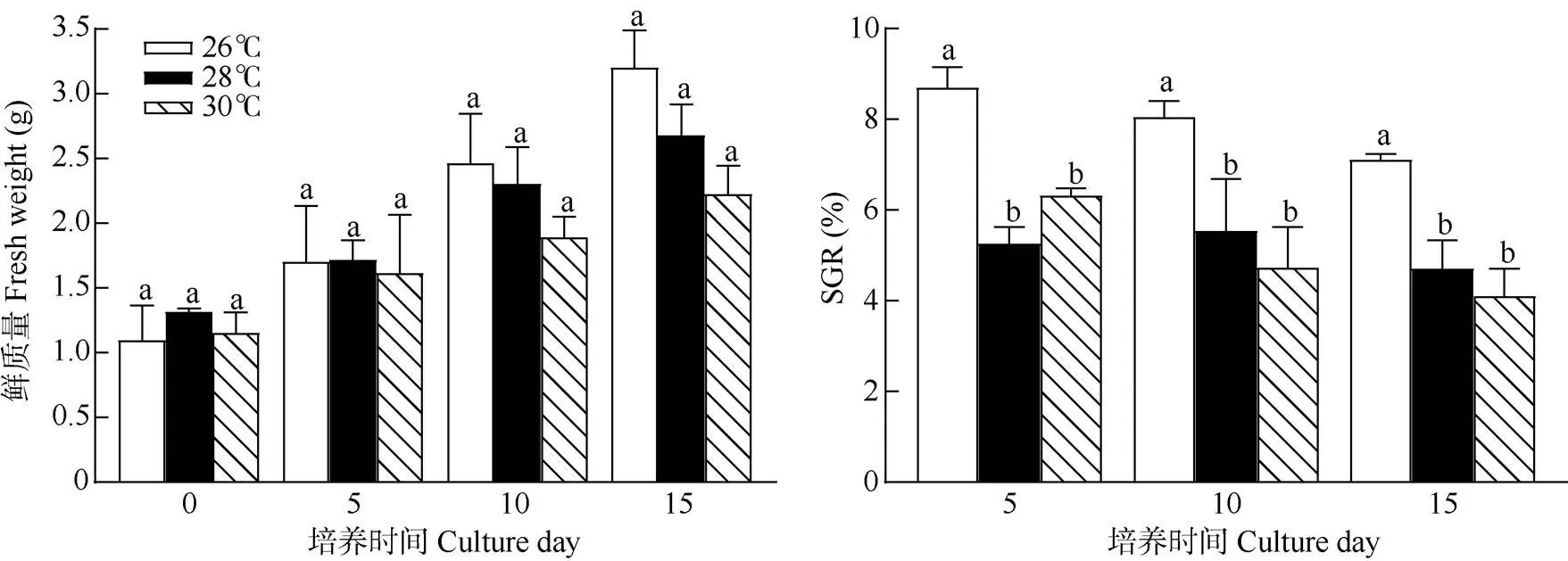
圖3 不同溫度下針葉蕨藻的生長和日特定生長率(SGR)
光照強度 從圖4可見,在不同光照強度下培養15 d,藻體的SGR差異不顯著(>0.05,=3)。
但在光照強度25.00mol/(m2·s)下的藻體生長最快,平均鮮質量從(0.87±0.04) g提高到(3.24±0.70) g, SGR最高,達(8.67±1.25)%;其次為31.25mol/(m2·s) 光照強度,培養15 d藻體的平均鮮質量提高了1.85g, SGR為(7.70±1.12)%;18.75mol/(m2·s)光照強度的最低,培養15 d藻體的平均鮮質量僅提高1.54g, SGR僅為(6.95±0.46)%, 說明針葉蕨藻在光照強度25.00mol/(m2·s)下的生長較好。
2.2 光合活性對環境因子的響應
對鹽度的響應 從表1可見,在25‰~30‰鹽度下藻體的熒光參數表現出一定差異,但不顯著(>0.05,=3)。不同鹽度下藻體的Fv/Fm為0.75~ 0.79,Yield為0.52~0.61,ETR為40.75~48.55。與鹽度25‰相比,27.5‰鹽度的平均Fv/Fm、Yield和ETR分別升高了4.32%、18.74%和19.14%;而當鹽度為30‰時,平均Fv/Fm、Yield和ETR則分別降低了2.28%、8.00%和8.25%。這表明針葉蕨藻的光合效率在鹽度27.5‰下最高,25‰鹽度的最低。鹽度27.5‰時藻體的平均qP最高,鹽度25‰的最低; 而藻體的平均qN則相反,以鹽度25‰最高。這說明在鹽度27.5‰下藻體光合活性最大且熱耗散最小。
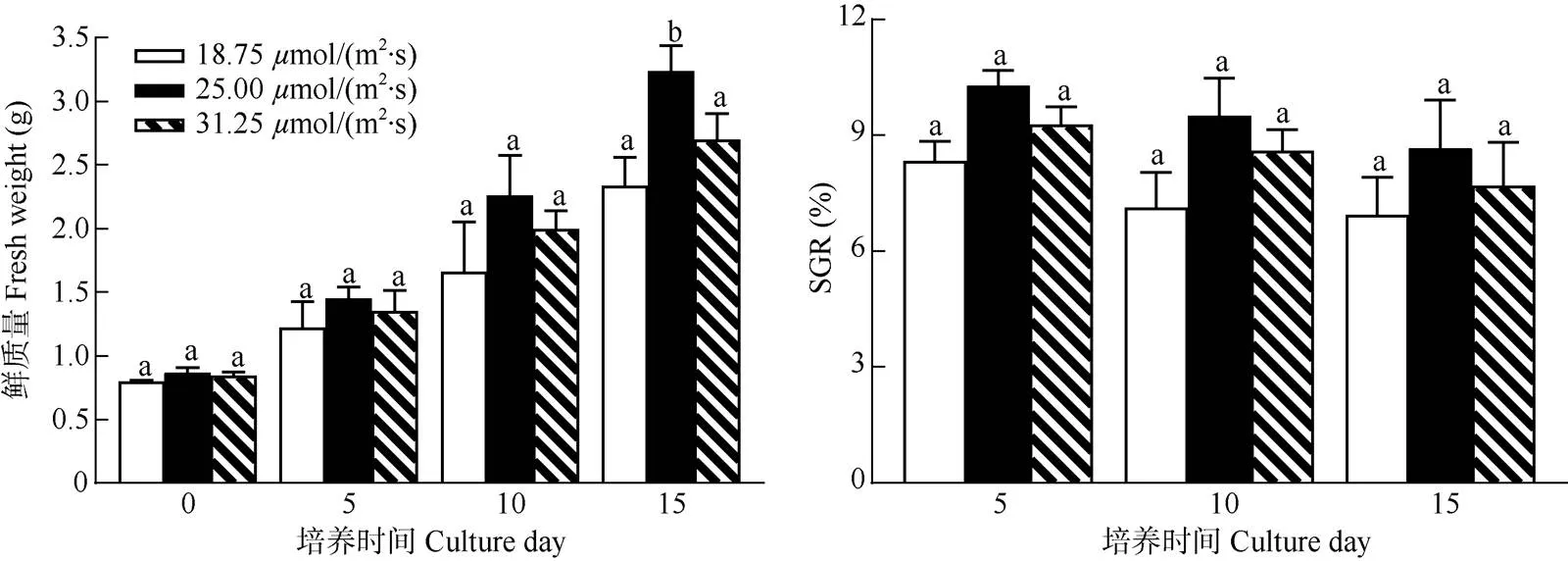
圖4 不同光照強度下針葉蕨藻的生長和日特定生長率(SGR)
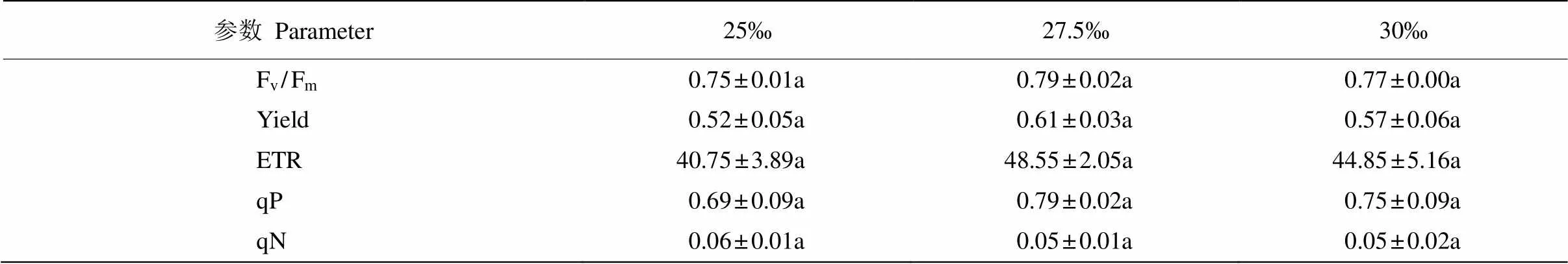
表1 不同鹽度下針葉蕨藻的Fv/Fm、Yield、ETR、qP和qN
同行數據后不同字母表示差異顯著(<0.05,=3)。下表同。
Data followed different letters within line indicate significant differences at 0.05 level (=3). The same is followed Tables.
對溫度的響應 從表2可見,藻體的熒光參數在26℃~30℃間表現出一定差異,但不顯著(> 0.05,=3)。不同溫度下藻體的平均Fv/Fm為0.77~ 0.79,Yield為0.55~0.61,ETR為43.15~47.50,與26℃相比,28℃下藻體的平均Fv/Fm、Yield和ETR分別降低了2.79%、6.69%和6.11%;而當溫度升至30℃時,藻體的平均Fv/Fm基本不變,Yield和ETR又下降了3.01%和3.36%。這表明針葉蕨藻的光合效率在26℃下最高,并隨溫度上升而下降。藻體的平均qP在26℃下最高,并隨溫度上升而降低;而藻體的平均qN則相反,以26℃的最低,并隨溫度上升而增高。這表明在26℃下,藻體光合活性最大且熱耗散最小。
對光照強度的響應 從表3可見,藻體熒光參數在光照強度18.75~25.00mol/(m2·s)下表現出一定差異,僅最大光量子產量有顯著差異(<0.05,=3),其余參數的差異均不顯著。不同光照強度下藻體的平均Fv/Fm為0.74~0.79,光照強度為25.00和31.25mol/(m2·s)時均為0.79±0.02, 但光照強度18.75mol/(m2·s)時顯著較低(<0.05,=3), 僅為0.74±0.001。不同光照強度下藻體的平均Yield為0.54~0.62, ETR為42.45~48.80, 與18.75mol/(m2·s)相比,光照強度為25.00mol/(m2·s)時,藻體的平均Yield和ETR分別升高了14.98%和14.96%;而光照強度為31.25mol/(m2·s)時,藻體的平均Yield和ETR則分別降低了4.30%和4.61%,表明針葉蕨藻的光合效率在光照強度25.00mol/(m2·s)下最高,與31.25mol/(m2·s)的相近,而在18.75mol/(m2·s)光照強度下藻體光合作用受到輕微抑制。在光照強度31.25mol/(m2·s)下藻體的平均qP最高, 18.75mol/(m2·s)的最低;而平均qN則相反,以18.75mol/(m2·s)的最高,這表明25.00mol/(m2·s) 光照強度下,藻體的光合活性最大且熱耗散最小, 而在18.75mol/(m2·s)下藻體光保護能力最高。
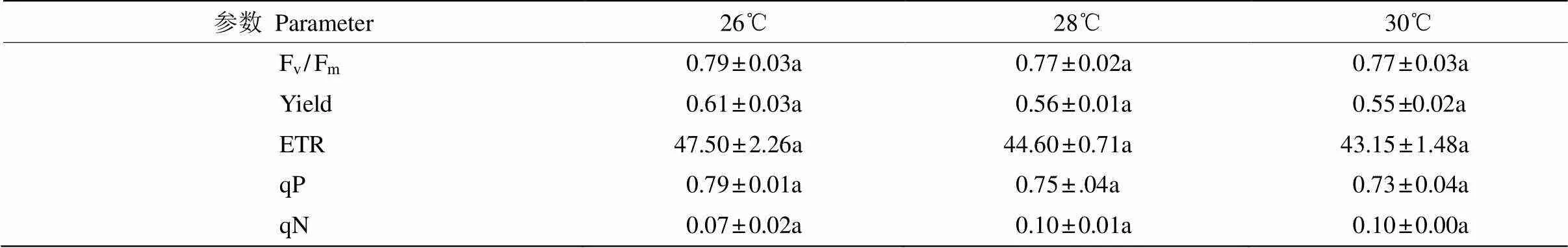
表2 不同溫度下針葉蕨藻的Fv/Fm、Yield、ETR、qP和qN

表3 不同光照強度下針葉蕨藻的Fv/Fm、Yield、ETR、qP和qN
3 結論和討論
3.1 生長量和生長率
大型海藻在生長過程中通過吸收海水中的營養物質,同化成自身需要的營養成分,且能夠儲存大量的營養鹽,成為海洋生態系統中重要的氮庫和磷庫,進而保護海洋生態環境[10–14]。
鹽度是影響海藻生長的重要環境因子。姚瑤[5]的研究表明針葉蕨藻在鹽度35‰下生長最好,SGR可達(3.08±0.44)%;Mosquera-Murillo等[6]則認為其在鹽度25‰下生長最快,SGR達4.82%。本研究結果表明,在鹽度27.5‰下針葉蕨藻的生長量和生長率最大,SGR高達(6.35±0.67)%。溫度是影響大型海藻吸收營養的重要因子[15],袁榮榮等[7]的研究表明,針葉蕨藻在25℃~30℃的生長較好,本研究也證明26℃時針葉蕨藻生長最好,能最快適應所處環境。光照強度是藻類生長發育的又一重要影響因子,對其代謝物質的合成與積累具有重要影響[16],本研究結果表明,光照強度為18.75~25.00mol/(m2·s), 針葉蕨藻的SGR隨光照強度升高而增加,25.00~31.25mol/(m2·s)時SGR隨光照強度升高而下降, 這與袁榮榮等[7]的研究結果相似,但光強過弱亦會影響其生長。
3.2 葉綠素熒光參數
葉綠素熒光參數能夠反映植物葉綠體PS I和PS II (主要是PS II)的光能吸收利用、傳遞及耗散過程[17],葉綠素熒光技術廣泛應用于不同環境因子對植物光合作用的影響研究[18]。
Fv/Fm是PS II的最大光能轉換效率,即最大光化學量子產量,可受作用于PS II的脅迫條件的影響,是反映植物所受生理壓力的探針,用來監測海藻的生活環境是否良好[19]。而在無脅迫條件下大型海藻的Fv/Fm變化極小,已有研究表明,鹽度對羊棲菜()幼體、強壯硬毛藻()的Fv/Fm影響不顯著(>0.05)[20–21]。本試驗中,針葉蕨藻的Fv/Fm為0.74~0.79。對大型海藻的研究表明[22–24],健康綠藻約為0.75,基本不低于0.70。故可認為本試驗中針葉蕨藻在光照強度18.75mol/(m2·s)下受到了輕微光抑制,而在其余環境條件下藻體生理狀況良好,并未受到脅迫。在不同水平的3種環境因子下,藻體的Fv/Fm表現出一定差異,分別在27.5‰鹽度、26℃和25.00~ 31.25mol/(m2·s)光照強度下達到最大;且光照強度對針葉蕨藻的Fv/Fm影響最大,這與王陽陽等[25]認為扁滸苔的熒光活性對光照強度變化最為敏感的研究結果相似。
Yield是指PS II實際光化學效率,反映光照下PS II反應中心部分關閉時的實際光化學效率;ETR是經過PS II的相對線性電子流速率[26]。有研究表明,ETR與光合釋氧或CO2的固定速率呈線性關系[27]。本試驗中,在鹽度25‰~30‰、光照18.75~ 25.00mol/(m2·s)時,藻體的Yield和ETR隨鹽度或光照強度的上升呈先上升后下降的趨勢;溫度為26℃~30℃時,藻體的Yield和ETR則隨溫度上升而下降。可見,在鹽度27.5‰、26℃和光照強度25.00mol/(m2·s)下能夠提高針葉蕨藻PS II電子傳遞活性,從而獲得最高固碳效率。
qP為光化學淬滅系數,是PS II吸收的能量用于進行光化學反應的比例,反映光合活性的高低; qN為非光化學淬滅系數,是PS II吸收的能量耗散為熱量的比例,也就是植物耗散過剩光能為熱量的能力,即光保護能力[28]。本試驗中,藻體的qP隨鹽度變化的趨勢為鹽度25‰<30‰<27.5‰,隨溫度變化的趨勢為30℃<28℃<26℃,隨光照強度[mol/(m2·s)]變化的趨勢為18.75<25.00<31.25。與qP的變化趨勢相反,藻體qN隨鹽度上升呈先升后降,隨溫度上升而上升,隨光照強度降低呈先降后升,表明在鹽度27.5‰、26℃和25.00mol/(m2·s)光照強度下,針葉蕨藻的光合活性最大,通過熱耗散丟失的能量最少。
綜上所述,針葉蕨藻在鹽度27.5‰、26℃和光照強度25.00mol/(m2·s)下的光合活性及固碳效率最大,熱耗散丟失的能量最少,因而生長最好且光合活性最高。本研究基于大型海藻光合熒光特性, 探究不同環境條件下針葉蕨藻的光合能力,進一步確定了針葉蕨藻培育的最適環境條件,為其開發利用奠定了基礎。
[1] ZHANG S Y, SUN H C. Research progress on seaweed bed ecosystem and its engineering [J]. Chin J Appl Ecol, 2007, 18(7): 1647–1653.
章守宇, 孫宏超. 海藻場生態系統及其工程學研究進展[J]. 應用生態學報, 2007, 18(7): 1647–1653.
[2] ROBERTSON A I, DUKE N C. Mangroves as nursery sites: Compa- risons of the abundance and species composition of fish and crusta- ceans in mangroves and other nearshore habitats in tropical Australia [J]. Mar Biol, 1987, 96(2): 193–205. doi: 10.1007/BF00427019.
[3] LIU Y, LI L, SUN D C. Development prospect and the current situation of ornamental fish market in Shanghai [J]. J Anhui Agric Sci, 2015, 43(12): 347–351. doi: 10.13989/j.cnki.0517-6611.2015.12.119.
劉穎, 李麗, 孫大川. 上海觀賞水族市場調查及發展前景分析[J]. 安徽農業科學, 2015, 43(12): 347–351. doi: 10.13989/j.cnki.0517- 6611.2015.12.119.
[4] ZHENG X, CUI Y L, LIU A D, et al. Purification of nitrogen and phosphorus from mariculture effluent by[J]. J Trop Biol, 2016, 7(1): 43–47. doi: 10.15886/j.cnki.rdswxb.2016.01.007.
鄭興, 崔云亮, 劉傲東, 等. 針葉蕨藻對養殖廢水中氮鹽的凈化[J]. 熱帶生物學報, 2016, 7(1): 43–47. doi: 10.15886/j.cnki.rdswxb.2016. 01.007.
[5] YAO Y. The influence of environmental factors on uptake kinetics mechanism of ammonia and growth of[D]. Haikou: Hainan University, 2016.
姚瑤. 環境因子對針葉蕨藻()生長及氨氮吸收動力學的影響[D]. 海口: 海南大學, 2016.
[6] MOSQUERA-MURILLO Z, PE?A-SALAMANCA E J. Effect of salinity on growth of the green alga(Bryopsi- dales, Chlorophyta) under laboratory conditions [J]. Hidrobiológica, 2016, 26(2): 277–282.
[7] YUAN R R, HE W H, SONG H Y, et al. Preliminary study on effect of environmental factors on growth of feather algae [C]// Proceedings of the 2015 Annual Meeting of the Chinese Society for Environmental Sciences. Shenzhen: Chinese Society for Environmental Sciences, 2015: 1677–1682.
袁榮榮, 何文輝, 宋海燕, 等. 環境因子對羽毛藻生長的初步研究[C]// 2015年中國環境科學學會學術年會論文集.深圳: 中國環境科學學會, 2015: 1677–1682.
[8] CHENG F, LI L, YIN X L, et al. Comparison on purification effect of three macroalgae species on ammonia salt in inorganic and organic water [J]. Genom Appl Biol, 2017, 36(3): 1084–1089. doi: 10.13417/j. gab.036.001084.
程芬, 李樂, 殷小龍, 等. 3種海藻對無機鹽和有機物水體中氨態鹽凈化效果比較[J]. 基因組學與應用生物學, 2017, 36(3): 1084–1089. doi: 10.13417/j.gab.036.001084.
[9] WANG S S. Effects of environmental factors on egg laying and embryo development of[D]. Zhoushan: Zhejiang Ocean University, 2015.
王珊珊. 環境因子對棘頰雀鯛的產卵及胚胎發育的影響[D]. 舟山: 浙江海洋學院, 2015.
[10] LüNING K K. Lüning Seaweeds: Their environment, biogeography, and ecophysiology [J]. J Mar Biol Assoc U K, 1991, 71(1): 246. doi: 10.1017/S0025315400037632.
[11] ZHANG S Y, LIU S R, ZHOU X J, et al. Ecological function of seaweed-formed habitat and discussion of its application to sea ranching [J]. J Fish China, 2019, 43(9): 2004–2014. doi: 10.11964/jfc. 20190711873.
章守宇, 劉書榮, 周曦杰, 等. 大型海藻生境的生態功能及其在海洋牧場應用中的探討[J]. 水產學報, 2019, 43(9): 2004–2014. doi: 10.11964/jfc.20190711873.
[12] MAO Y Z, LI J Q, XUE S Y, et al. Ecological functions of the kelpin integrated multi-trophic aquaculture, Sanggou Bay, China [J]. Acta Ecol Sin, 2018, 38(9): 3230–3237. doi: 10.5846/ stxb201703160444.
毛玉澤, 李加琦, 薛素燕, 等. 海帶養殖在桑溝灣多營養層次綜合養殖系統中的生態功能[J]. 生態學報, 2018, 38(9): 3230–3237. doi: 10.5846/stxb201703160444.
[13] DUARTE C M, WU J P, XIAO X, et al. Can seaweed farming play a role in climate change mitigation and adaptation? [J]. Front Mar Sci, 2017, 4: 100. doi: 10.3389/fmars.2017.00100.
[14] XU S N, WEN S S, WU W X, et al. Bioremediation of caged fish aqua- culture by the red algain an integrated multi- trophic aquaculture system [J]. Acta Ecol Sin, 2008, 28(4): 1466–1475. doi: 10.3321/j.issn:1000-0933.2008.04.013.
徐姍楠, 溫珊珊, 吳望星, 等. 真江蘺()對網箱養殖海區的生態修復及生態養殖匹配模式[J]. 生態學報, 2008, 28(4): 1466–1475. doi: 10.3321/j.issn:1000-0933.2008.04.013.
[15] TALBOT P, DE LA NOüE J. Tertiary treatment of wastewater with(Schmidle) under various light and temperature conditions [J]. Water Res, 1993, 27(1): 153–159. doi: 10.1016/0043- 1354(93)90206-W.
[16] ZHANG Z S, WANG X M, LIU F, et al. Analysis of physiology and regulation mechanism ofunder stress [J]. Mar Environ Res, 2020, 39(3): 473–479. doi: 10.13634/j.cnki.mcs.2020.03.023.
張忠山, 王曉梅, 劉峰, 等. 石莼屬綠藻應答環境脅迫的生理特征與機制研究進展[J]. 海洋環境科學, 2020, 39(3): 473–479. doi: 10.13634/j.cnki.mcs.2020.03.023.
[17] WANG L L, LI Q, SONG X H, et al. Effects of environmental conditions on chlorophyll fluorescence parameters of plants [J]. J Shenyang Norm Univ (Nat Sci), 2019, 37(4): 362–367. doi: 10.3969/j. issn.1673-5862.2019.04.013.
王蘭蘭, 李琦, 宋曉卉, 等. 環境條件對植物葉綠素熒光參數影響研究進展[J]. 沈陽師范大學學報(自然科學版), 2019, 37(4): 362–367. doi: 10.3969/j.issn.1673-5862.2019.04.013.
[18] ZHANG S R. A discussion on chlorophyll fluorescence kinetics parameters and their significance [J]. Chin Bull Bot, 1999, 16(4): 444– 448. doi: 10.3969/j.issn.1674-3466.1999.04.021.
張守仁. 葉綠素熒光動力學參數的意義及討論 [J]. 植物學通報, 1999, 16(4): 444–448. doi: 10.3969/j.issn.1674-3466.1999.04.021.
[19] MAXWELL K, JOHNSON G N G. Chlorophyll fluorescence: A practical guide [J]. J Exp Bot, 2000, 51(345): 659–668. doi: 10.1093/jexbot/51. 345.659.
[20] SONG X D, CHEN B B, MA Z L, et al. Effects of salinity level on photosynthetic characteristics ofseedlings [J]. Acta Agric Zhejiang, 2020, 32(9): 1634–1644. doi: 10.3969/j.issn. 1004–1524.2020.09.12.
宋新丹, 陳斌斌, 馬增嶺, 等. 鹽度對羊棲菜()幼體光合特性的影響 [J]. 浙江農業學報, 2020, 32(9): 1634–1644. doi: 10.3969/j.issn.1004–1524.2020.09.12.
[21] HAN W J, XU Y T, JIANG Y Y, et al. Effects of N/P ratio, salinity and pH on growth and photosynthesis of sea weed[J]. J Dalian Ocean Univ, 2019, 34(6): 776–784. doi: 10.16535/j.cnki. dlhyxb.2019–014.
韓汶均, 許彥婷, 蔣陽楊, 等. 氮磷比、鹽度、pH對強壯硬毛藻生長和光合作用的影響[J]. 大連海洋大學學報, 2019, 34(6): 776–784. doi: 10.16535/j.cnki.dlhyxb.2019–014.
[22] YANG X Z, ZHENG X Q, LIN R C, et al. Photosynthetic capacity of three common species of macroalgae and the application in coral aquarium [J]. Chin J Ecol, 2014, 33(6): 1528–1533. doi: 10.13292/j. 1000-4890.20140327.006.
楊小舟, 鄭新慶, 林榮澄, 等. 三種大型綠藻光合能力的差異及其在珊瑚養殖中的應用[J]. 生態學雜志, 2014, 33(6): 1528–1533. doi: 10.13292/j.1000-4890.20140327.006.
[23] HAN Z G, HE L J, GU J G, et al. Photosynthetic performance ofduring its dehydration and rehydration[J]. Chin J Ecol, 2005, 24(11): 1291–1294. doi: 10.13292/j.1000-4890.2006.0146.
韓志國, 賀立靜, 顧繼光, 等. 束生剛毛藻脫水和復水過程中光合作用的變化[J]. 生態學雜志, 2005, 24(11): 1291–1294. doi: 10. 13292/j.1000-4890.2006.0146.
[24] ZHANG S Y, XIANG C, ZHOU X J, et al. Photosynthetic fluorescence characteristics of six macroalgae species in seaweed beds of Gouqi Island, Zhejiang, China [J]. Chin J Appl Ecol, 2018, 29(10): 3441– 3448. doi: 10.13287/j.1001-9332.201810.031.
章守宇, 向晨, 周曦杰, 等. 枸杞島海藻場6種大型海藻光合熒光特性比較[J]. 應用生態學報, 2018, 29(10): 3441–3448. doi: 10. 13287/j.1001-9332.201810.031.
[25] WANG Y Y, HUO Y Z, CAO J C, et al. Influence of low temperature and low light intensity on growth of[J]. J Fish Sci China, 2010, 17(3): 593–599.
王陽陽, 霍元子, 曹佳春, 等. 低溫、低光照強度對扁滸苔生長的影響[J]. 中國水產科學, 2010, 17(3): 593–599.
[26] HARBINSON J, GENTY B, BAKER N R. Relationship between the quantum efficiencies of photosystems I and II in pea leaves [J]. Plant Physiol, 1989, 90(3): 1029–1034. doi: 10.1104/pp.90.3.1029.
[27] BEER S, ILAN M.measurements of photosynthetic irradianceresponses of two Red Sea sponges growing under dim light conditions [J]. Mar Biol, 1998, 131(4): 613–617. doi: 10.1007/s002270050353.
[28] SCHREIBER U, SCHLIWA U, BILGER W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer [J]. Photosynth Res, 1986, 10(1/2): 51–62. doi: 10.1007/BF00024185.
Effects of Salinity, Temperature and Light Intensity on Growth and Photo- synthetic Activity of
Zhong Yiyun1a,1b, Yang Yunqi1a, GAO Xiaofeng1a,1b, Xing Hao1a,1b, LIU Wei1a,1b, DUAN Yuanliang1a,1b,2, HE Wenhui1a,3, HE Peimin1a,1b,3
(1a. College of Marine Ecology and Environment; 1b. Institute of Marine Science, Shanghai Ocean University, Shanghai 201306, China; 2. Sichuan Fisheries Research Institute,Chengdu 611731, China; 3. National Demonstration Center for Experimental Fisheries Science Education (Shanghai Ocean University),Shanghai 201306, China)
In order to explore the effects of environment factors on growth of, the changes in growth and chlorophyll fluorescence parameters were studied under different salinity, temperature and light intensity. The results showed that the specific growth rate (SGR), maximum light quantum yield (Fv/Fm), actual photosynthetic efficiency (Yield), electron transfer rate (ETR) and photochemical quenching (qP) increased at first and then decrease with increasing salinity, while non-photochemical quenching (qN) was opposite. The photosynthetic activity and carbon fixation efficiency of the algae under 27.5‰ salinity were the highest, and there were significant difference with those under 25‰ and 30‰ salinity (<0.05,=3). The SGR, Fv/Fm, Yield,ETR and qP of the algae decreased with temperature rising, and qN was opposite. The photosynthetic activity and carbon fixation efficiency of algae under 26℃ were the highest, and there were significant difference with those under 28℃and 30℃(<0.05,=3). The SGR, Fv/Fm, Yield, ETR and qP of the algae increased at first and then decreased with increment of light intensity, while qN was opposite, and there was slight photoinhibition under 18.75mol/(m2·s) light intensity. The growth, photosynthetic activity and carbon fixation efficiency of algae were the highest under 25.00mol/(m2·s) light intensity, but no significant difference with those under 18.75 and 27.5mol/(m2·s) light intensity (>0.05,=3). Therefore, it was suggested thatcould grow fast with high photosynthesis ability under the conditions of 27.5‰ salinity, 26℃, and 25.00mol/(m2·s) light intensity.
; Chlorophyll fluorescence parameter; Specific growth rate; Salinity; Temperature; Light intensity
10.11926/jtsb.4378
2021-01-14
2021-03-15
上海市海洋局科研項目(滬海科2015-02)資助
This work was supported by the Project for Scientific Research of Shanghai Municipal Bureau of Oceanography (Grant No. 2015-02).
鐘逸云(1995~ ),女,碩士研究生,主要研究方向為人工生態系統。E-mail: 13501619793@139.com
通信作者Corresponding author. E-mail: pmhe@shou.edu.cn
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
小讀者(2021年2期)2021-03-29 05:03:48
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年11期)2019-01-07 01:28:02
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
