黃酒貯存陳釀關(guān)鍵酸敗微生物的生物學(xué)特征
2021-12-17 08:08:36羅雪柯鋒劉文容徐巖陳雙
食品與發(fā)酵工業(yè) 2021年23期
關(guān)鍵詞:生長(zhǎng)
羅雪,柯鋒,劉文容,徐巖,陳雙*
1(江南大學(xué) 生物工程學(xué)院,江蘇 無(wú)錫,214122) 2(勁牌有限公司,湖北 黃石,435100)
黃酒是中國(guó)民族特色的酒精飲料[1],其典型特征是越陳越香。長(zhǎng)期貯存陳釀是生產(chǎn)高品質(zhì)黃酒必須的工藝環(huán)節(jié),但由于黃酒中含有豐富的糖類、氨基酸及多肽等營(yíng)養(yǎng)物質(zhì),易造成微生物污染生長(zhǎng),故在陳釀過(guò)程中時(shí)常出現(xiàn)酸敗變質(zhì)現(xiàn)象[2-6],給黃酒產(chǎn)業(yè)造成嚴(yán)重的經(jīng)濟(jì)損失[7-8]。
根據(jù)黃酒酸敗現(xiàn)象的不同,黃酒酸敗主要分為醋酸酸敗和乳酸酸敗[9],其中乳酸酸敗是造成高酒精度陳釀黃酒出現(xiàn)酸敗現(xiàn)象的主要原因,而乳桿菌(Lactobacillus)是引起乳酸酸敗的主要微生物。在密閉環(huán)境中、較高酒精度(18%,體積分?jǐn)?shù))條件下,乳酸酸敗時(shí)常發(fā)生。劉文容等[10]首次采用未培養(yǎng)技術(shù)與可培養(yǎng)技術(shù)相結(jié)合的方法,明確了食果糖乳桿菌(L.fructivorans)和耐酸乳桿菌(L.acetotolerans)為引起陳釀黃酒發(fā)生酸敗的主要微生物。章志超等[11]在確定從酸敗黃酒中分離得到的酸敗微生物為食果糖乳桿菌后,優(yōu)化了食果糖乳桿菌的最佳培養(yǎng)條件:在pH值為5.4的MRS固體培養(yǎng)基上,加入質(zhì)量分?jǐn)?shù)為0.08%的L-半胱氨酸和體積分?jǐn)?shù)為9%的乙醇,30 ℃下培養(yǎng)。但目前,對(duì)黃酒陳釀酸敗微生物的生物學(xué)特征的相關(guān)研究較少,對(duì)酸敗微生物的生理生化特征認(rèn)識(shí)不清,缺乏控制黃酒酸敗微生物的有效措施,這已成為黃酒產(chǎn)業(yè)發(fā)展的限制因素[12-13]。
貯存陳釀過(guò)程的微生物酸敗是酒精飲料行業(yè)中普遍存在的問(wèn)題。日本學(xué)者研究清酒微生物酸敗現(xiàn)象發(fā)現(xiàn),食果糖乳桿菌和同型腐酒乳桿菌(L.homohiochi)是導(dǎo)致清酒貯存陳釀酸敗的主要微生物[14-16],這2種酸敗微生物都具有很高的酒精耐受度,兩者適宜生長(zhǎng)的pH值都大約在4.5~5.0[17-18]。食果糖乳桿菌具有較好的耐熱性,在巴氏滅菌結(jié)束后仍可能繼續(xù)存活[19-20]。研究啤酒微生物酸敗現(xiàn)象發(fā)現(xiàn),導(dǎo)致啤酒酸敗的主要酸敗微生物是耐酸乳桿菌,這類酸敗微生物具有很強(qiáng)的乙醇耐受性和酒花耐受性[21-22]。由于對(duì)啤酒和清酒酸敗微生物生物學(xué)特征的系統(tǒng)研究,目前啤酒和日本清酒中都建立了針對(duì)這類酸敗微生物的有效控制方法體系。
針對(duì)黃酒貯存陳釀過(guò)程中存在的酸敗微生物的生物學(xué)特征認(rèn)識(shí)不清的問(wèn)題,本研究通過(guò)對(duì)黃酒陳釀關(guān)鍵酸敗微生物——食果糖乳桿菌和耐酸乳桿菌的生理生化特征進(jìn)行分析,包括營(yíng)養(yǎng)需求、糖分消耗、生長(zhǎng)溫度耐受、酒精耐受等4個(gè)方面,為黃酒陳釀過(guò)程中酸敗微生物的科學(xué)控制提供理論依據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)材料
1.1.1 實(shí)驗(yàn)樣品
菌種:食果糖乳桿菌(Lactobacillusfructivorans)和耐酸乳桿菌(Lactobacillasacetotolerans),江南大學(xué)釀造微生物與應(yīng)用酶學(xué)實(shí)驗(yàn)室保藏;正常黃酒酒樣,紹興某黃酒廠按照傳統(tǒng)紹興黃酒釀造工藝釀造。
1.1.2 試劑和儀器
無(wú)水乙醇,國(guó)藥集團(tuán)化學(xué)試劑有限公司;甲瓦龍酸,北京百靈威科技有限公司;MRS培養(yǎng)基,美國(guó)Oxoid公司;厭氧培養(yǎng)盒、厭氧產(chǎn)氣袋,日本三菱瓦斯化學(xué)株式會(huì)社;全波長(zhǎng)酶標(biāo)儀,Thermo公司。
1.1.3 培養(yǎng)基
粉末狀MRS培養(yǎng)基:8.0 g牛肉膏,20.0 g葡萄糖,10.0 g蛋白胨,2.0 g檸檬酸三銨,0.2 g MgSO4,2.0 g K2HPO4,5.0 g乙酸鈉,0.05 g MnSO4,4.0 g酵母提取物,1 mL山梨醇單油酸酯,終pH調(diào)至5.7。
添加甲瓦龍酸的MRS培養(yǎng)基:5.0 mg甲瓦龍酸,52.0 g粉末狀MRS培養(yǎng)基,800~850 mL無(wú)菌水,150~200 mL無(wú)水乙醇,終pH調(diào)至5.0。
培養(yǎng)基滅菌條件:115 ℃滅菌20 min。
1.2 實(shí)驗(yàn)方法
1.2.1 黃酒關(guān)鍵酸敗微生物理化特征分析
1.2.1.1 酸敗微生物營(yíng)養(yǎng)需求分析
本研究用生態(tài)板(Biolog-ECO)方法探究了黃酒酸敗微生物的營(yíng)養(yǎng)需求情況。首先取10 mL黃酒酸敗微生物培養(yǎng)液(處于對(duì)數(shù)生長(zhǎng)期),經(jīng)1 000 r/min離心5 min后,將細(xì)胞沉淀置于裝有接種液的10 mL通用(general purpose,GP)管中重懸混勻,再取100 μL上述重懸液轉(zhuǎn)移到GP2微孔板中(GP2微孔板中有氨基酸、有機(jī)酸糖類等95種底物),最后將微孔板置于厭氧、30 ℃環(huán)境中培養(yǎng)72 h后,在波長(zhǎng)為590 nm下測(cè)定吸光度,并與以水為底物的A1微孔板在相同情況下測(cè)定的吸光度作差值,得到酸敗微生物對(duì)各種營(yíng)養(yǎng)物質(zhì)的吸收情況。
1.2.1.2 酸敗微生物對(duì)不同含量還原糖利用情況分析
首先取未發(fā)生酸敗的黃酒酒樣,將酒樣糖含量分別調(diào)至2、5、10 g/L,再在酒樣中接種2%的酸敗微生物培養(yǎng)液(處于對(duì)數(shù)生長(zhǎng)期),在厭氧、30 ℃條件下培養(yǎng)46 d,同時(shí)在培養(yǎng)過(guò)程中測(cè)定黃酒中糖度和酸度的變化情況。
1.2.1.3 酸敗微生物溫度耐受分析
首先取未發(fā)生酸敗的黃酒酒樣,將酒樣糖含量調(diào)至10 g/L,保證微生物可正常生長(zhǎng),再在黃酒酒樣中接種2%的酸敗微生物培養(yǎng)液(處于對(duì)數(shù)生長(zhǎng)期),最后將上述酒樣置于溫度分別為11、20、25、30、35和40 ℃下,厭氧培養(yǎng)20 d后,測(cè)定不同溫度下培養(yǎng)得到的黃酒在波長(zhǎng)為600 nm下的吸光度,判斷溫度變化對(duì)黃酒酸敗微生物生長(zhǎng)產(chǎn)生的影響。
1.2.1.4 酸敗微生物酒精耐受分析
首先在MRS-甲瓦龍酸培養(yǎng)基中添加無(wú)水乙醇,將各個(gè)培養(yǎng)基的酒精度(體積分?jǐn)?shù))分別調(diào)到14%、18%、20%、23%、25%和30%,再在培養(yǎng)基中接種2%的酸敗微生物培養(yǎng)液(處于對(duì)數(shù)生長(zhǎng)期),最后在厭氧、30 ℃條件下培養(yǎng)2周,測(cè)定培養(yǎng)基中酸敗微生物數(shù)量和還原糖變化量,探究酸敗微生物在不同酒精度培養(yǎng)基中的生長(zhǎng)狀況。根據(jù)工廠的實(shí)際生產(chǎn)情況,首先取未發(fā)生酸敗的黃酒酒樣(酒精度為14%)添加無(wú)水乙醇,將黃酒的酒精度分別調(diào)到14%、18%、20%、23%、25%和30%,再在黃酒中接種2%的黃酒酸敗微生物培養(yǎng)液(處于對(duì)數(shù)生長(zhǎng)期),最后在厭氧、30 ℃條件下培養(yǎng)2周,測(cè)定黃酒中酸敗微生物數(shù)量和還原糖變化量,探究酸敗微生物在不同酒精度黃酒中的生長(zhǎng)狀況。
2 結(jié)果與分析
2.1 酸敗微生物營(yíng)養(yǎng)需求特征解析
選擇耐酸乳桿菌30-1、19-1-1和食果糖乳桿菌13-2、7-2進(jìn)行實(shí)驗(yàn),培養(yǎng)結(jié)束后,接種不同酸敗微生物的GP2微孔板與以水為底物的A1微孔板在波長(zhǎng)590 nm下的吸光度差值變化情況如下表1所示。

表1 酸敗微生物對(duì)不同營(yíng)養(yǎng)物質(zhì)的利用情況Table 1 The use of different nutrients by spoilage microbes
酸敗微生物主要以糖類作為能源物質(zhì),能夠消耗部分有機(jī)酸,基本不消耗氨基酸。在糖類中,酸敗微生物對(duì)葡萄糖的利用能力最強(qiáng),對(duì)麥芽糖的利用能力次之,可以利用少量環(huán)糊精,幾乎不利用淀粉作為能源物質(zhì);在有機(jī)酸中,可以利用部分乳酸、L-蘋(píng)果酸、丙酮酸,幾乎不利用乙酸。耐酸乳桿菌對(duì)麥芽糖的利用能力明顯高于食果糖乳桿菌,而食果糖乳桿菌對(duì)環(huán)糊精的利用能力則更強(qiáng)。傳統(tǒng)黃酒中含有糖分、有機(jī)酸及氨基酸等營(yíng)養(yǎng)物質(zhì),這些營(yíng)養(yǎng)物質(zhì)特別是糖類能夠?yàn)樗釘∥⑸锏纳L(zhǎng)提供有利條件,因此可以通過(guò)控制黃酒中糖分的含量來(lái)抑制酸敗微生物的生長(zhǎng)。本研究還發(fā)現(xiàn),黃酒中酸敗微生物與日本清酒中的酸敗微生物在營(yíng)養(yǎng)需求上具有一定的相似性。日本清酒中的酸敗微生物也是主要利用葡萄糖作為能源物質(zhì),還能夠消耗丙酮酸、檸檬酸、蘋(píng)果酸等有機(jī)酸作為營(yíng)養(yǎng)物質(zhì)[16]。
2.2 還原糖含量對(duì)黃酒關(guān)鍵酸敗微生物的影響
比較黃酒酸敗關(guān)鍵微生物在不同糖含量黃酒中糖分消耗和酸度的變化情況,如圖1和圖2所示。在不同糖分含量的黃酒中,2類微生物的生長(zhǎng)趨勢(shì)大體一致,黃酒的有機(jī)酸含量均隨著還原糖的消耗而上升,說(shuō)明2類酸敗微生物代謝黃酒中的糖分生成了有機(jī)酸。糖代謝和有機(jī)酸生成曲線顯示黃酒中的還原糖含量越高,酸敗微生物代謝產(chǎn)生的有機(jī)酸含量就會(huì)越高,當(dāng)黃酒中的總酸含量達(dá)到一定濃度時(shí),就會(huì)發(fā)生酸敗現(xiàn)象,因此可以通過(guò)調(diào)節(jié)黃酒糖度來(lái)控制黃酒中的有機(jī)酸含量,預(yù)防黃酒發(fā)生酸敗。黃酒陳釀過(guò)程中酸敗現(xiàn)象的發(fā)生和黃酒發(fā)酵過(guò)程是相似的。在黃酒發(fā)酵過(guò)程中,乳酸菌代謝糖類產(chǎn)生乳酸,導(dǎo)致黃酒中酸度上升,當(dāng)酸度達(dá)到一定濃度時(shí),黃酒就會(huì)發(fā)生酸敗現(xiàn)象[23]。
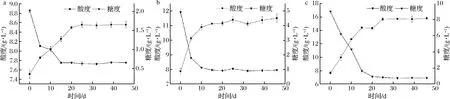
a-還原糖含量2 g/L;b-還原糖含量5 g/L;c-還原糖含量10 g/L圖1 還原糖含量黃酒對(duì)耐酸乳桿菌30-1代謝的影響Fig.1 The effects of reducing sugar content in Huangjiu on the metabolism of L.acetotolerans 30-1
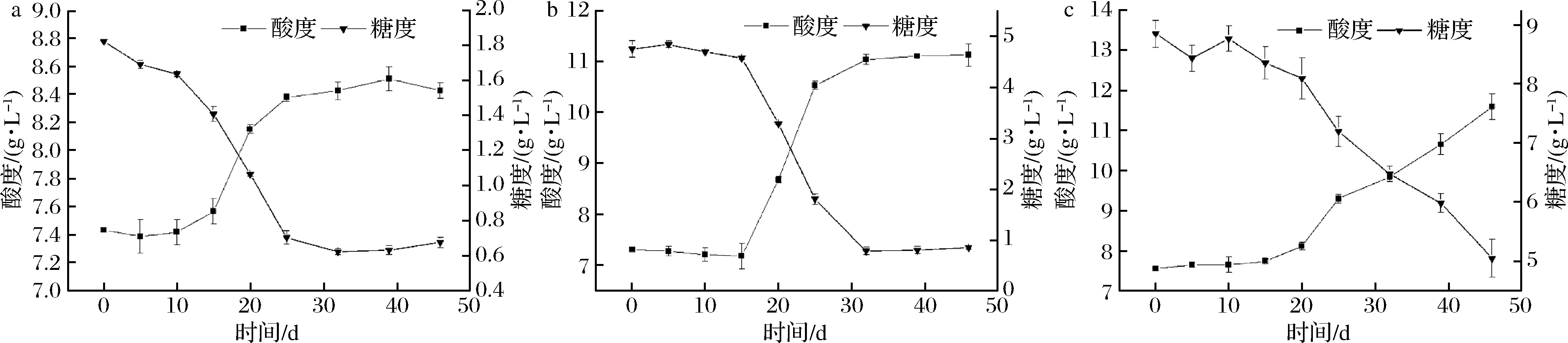
a-還原糖含量2 g/L;b-還原糖含量5 g/L;c-還原糖含量10 g/L圖2 還原糖含量黃酒對(duì)食果糖乳桿菌13-21代謝的影響Fig.2 The effects of reducing sugar content in Huangjiu on the metabolism of L.fructivorans 13-21
2.3 溫度對(duì)黃酒關(guān)鍵酸敗微生物的影響
分析黃酒中2類關(guān)鍵酸敗微生物在不同溫度下的生長(zhǎng)情況。如圖3和圖4所示,在20~30 ℃,2種酸敗微生物菌體濃度較高,生長(zhǎng)情況較好,而在11和40 ℃條件下,2種酸敗微生物幾乎不能生長(zhǎng)。在11 ℃的低溫環(huán)境下,酸敗微生物的生命活動(dòng)受到影響,生長(zhǎng)受到抑制。在40 ℃條件下,培養(yǎng)得到的黃酒中幾乎不存在酸敗微生物,表明在該溫度下長(zhǎng)時(shí)間培養(yǎng)會(huì)導(dǎo)致酸敗微生物死亡。為驗(yàn)證該結(jié)論的一般性,從上述兩類酸敗菌中分別挑選8株菌株,將微生物培養(yǎng)液(處于生長(zhǎng)對(duì)數(shù)期)接種到正常陳釀的黃酒中,再將黃酒置于30、40 ℃條件下,厭氧培養(yǎng),觀察記錄酸敗微生物的生長(zhǎng)狀況,所得結(jié)果如表2所示。在30 ℃條件下,2種酸敗微生物的生長(zhǎng)狀況良好,而在40 ℃條件下,2種酸敗微生物幾乎不能生長(zhǎng),從而驗(yàn)證了上述結(jié)論的一般性。因此,通過(guò)控制黃酒在較低的儲(chǔ)藏溫度或在黃酒殺菌過(guò)程中優(yōu)化殺菌的溫度、時(shí)間等參數(shù)能夠一定程度上抑制酸敗微生物的生長(zhǎng),防止黃酒酸敗現(xiàn)象的發(fā)生。這與章志超等[11]從酸敗黃酒中分離得到的食果糖乳桿菌在溫度方面的單因素試驗(yàn)結(jié)果基本吻合。分離得到的食果糖乳桿菌在30 ℃時(shí)生長(zhǎng)得最快,當(dāng)溫度升高時(shí),食果糖乳桿菌的生長(zhǎng)逐漸受到抑制,可計(jì)數(shù)菌落逐漸減少。

a-耐酸乳桿菌7-2;b-耐酸乳桿菌19-1-1圖3 耐酸乳桿菌在不同溫度下的生長(zhǎng)狀況Fig.3 The growth of L.acetotolerans at different temperatures
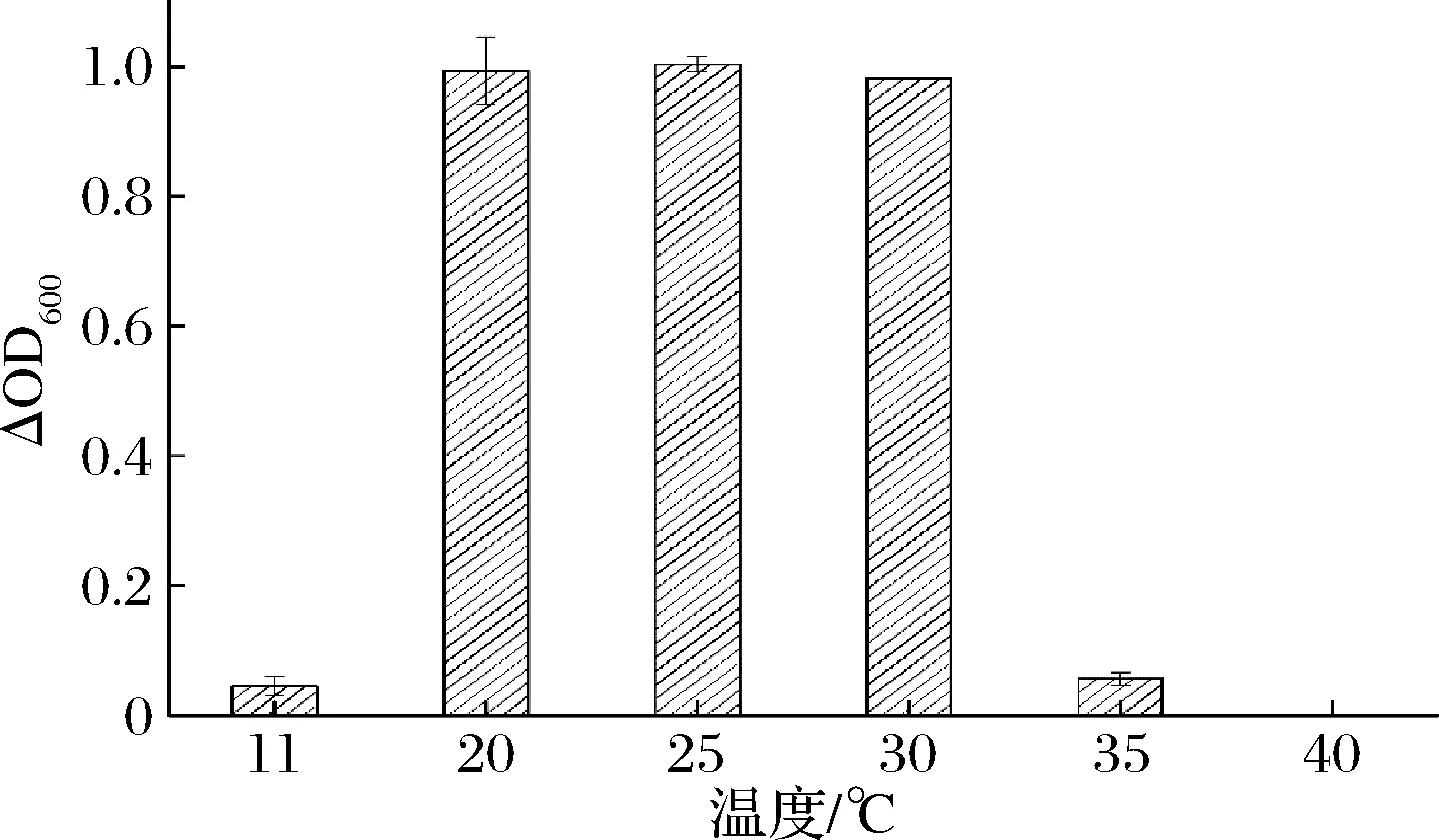
圖4 食果糖乳桿菌7-2在不同溫度下的生長(zhǎng)狀況Fig.4 The growth of L.fructivorans 7-2 at different temperatures

表2 耐酸乳桿菌和食果糖乳桿菌在不同溫度下的生長(zhǎng)狀況Table 2 The growth of L.fructivorans and L.acetotolerans at different temperatures
2.4 酒精度對(duì)黃酒關(guān)鍵酸敗微生物的影響
選擇耐酸乳桿菌30-1、19-1-1和食果糖乳桿菌13-2、7-2進(jìn)行實(shí)驗(yàn),培養(yǎng)結(jié)束后,不同酒精度培養(yǎng)基中酸敗微生物數(shù)量及還原糖變化量的測(cè)定結(jié)果如表3和圖5所示。結(jié)果表明,在低酒精度(14%,體積分?jǐn)?shù))下酸敗微生物的生長(zhǎng)能力很強(qiáng),2種酸敗微生物的數(shù)量增加較多,對(duì)還原糖的消耗量較大,有明顯的生長(zhǎng)跡象。隨著培養(yǎng)基中的酒精度上升,耐酸乳桿菌和食果糖乳桿菌的生長(zhǎng)逐漸受到抑制。當(dāng)酒精度達(dá)到23%(體積分?jǐn)?shù))時(shí),培養(yǎng)基中的2種酸敗微生物仍然能夠生長(zhǎng)產(chǎn)酸。當(dāng)酒精度達(dá)到25%(體積分?jǐn)?shù))以上時(shí),培養(yǎng)基中的2種酸敗微生物不存在生長(zhǎng)跡象,生長(zhǎng)明顯受到抑制。
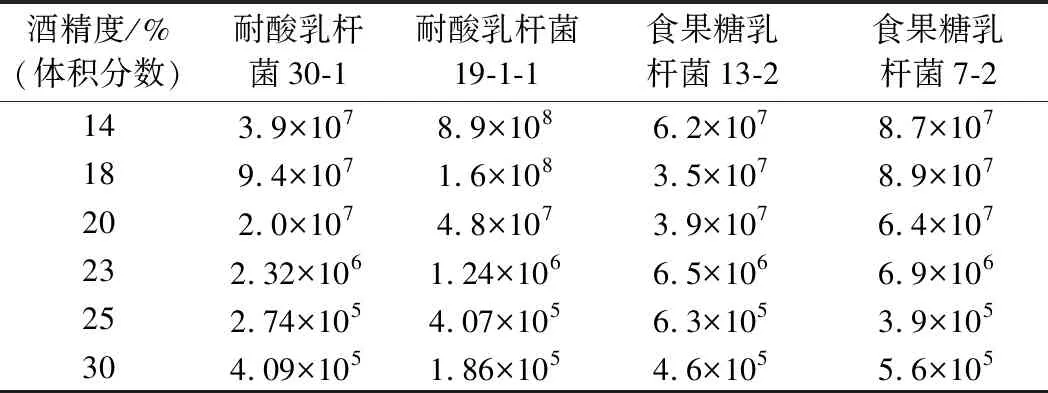
表3 不同酒精度培養(yǎng)基中酸敗微生物的數(shù)量 單位:個(gè)/mL
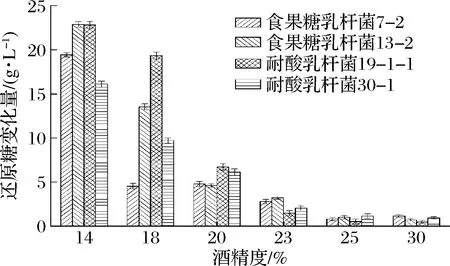
圖5 不同酒精度培養(yǎng)基中微生物對(duì)還原糖的消耗情況Fig.5 The consumption of reducing sugars by spoilage microbes in different alcoholic medium
為了驗(yàn)證黃酒酸敗微生物在真實(shí)黃酒中的酒精耐受性,根據(jù)工廠的實(shí)際生產(chǎn)情況,分析不同酒精度黃酒中酸敗微生物數(shù)量及還原糖變化量的測(cè)定結(jié)果。如表4和圖6所示,與食果糖乳桿菌7-2、13-1相比,耐酸乳桿菌19-1-1、30-1在黃酒中具有更強(qiáng)的生長(zhǎng)能力。在較低酒精度下,黃酒中耐酸乳桿菌的數(shù)量最多,對(duì)還原糖的消耗量較高,最高可達(dá)到17.86 g/L;而食果糖乳桿菌在不同酒精度黃酒中的數(shù)量相對(duì)較少,對(duì)還原糖的消耗量相對(duì)較低,都在4.5 g/L以下。當(dāng)酒精度達(dá)到23%(體積分?jǐn)?shù))時(shí),黃酒中的2種酸敗微生物仍然能夠生長(zhǎng)產(chǎn)酸。當(dāng)酒精度達(dá)到25%(體積分?jǐn)?shù))以上時(shí),黃酒中的2種酸敗微生物不存在生長(zhǎng)跡象,生長(zhǎng)明顯受到抑制。從以上的結(jié)果分析可以得到,食果糖乳桿菌和耐酸乳桿菌對(duì)酒精的耐受能力很強(qiáng),在低酒精度下有較強(qiáng)的生長(zhǎng)能力,在23%(體積分?jǐn)?shù))的酒精度下仍存在生長(zhǎng)跡象,在25%(體積分?jǐn)?shù))的酒精度下生長(zhǎng)受到抑制作用。與黃酒酸敗微生物相似,啤酒和日本清酒的酸敗微生物也具有較高的酒精耐受性,清酒酸敗微生物可達(dá)20%(體積分?jǐn)?shù))以上的酒精耐受性[16];而食醋中的微生物和黃酒酸敗微生物相反,幾乎不具有酒精耐受性[24]。
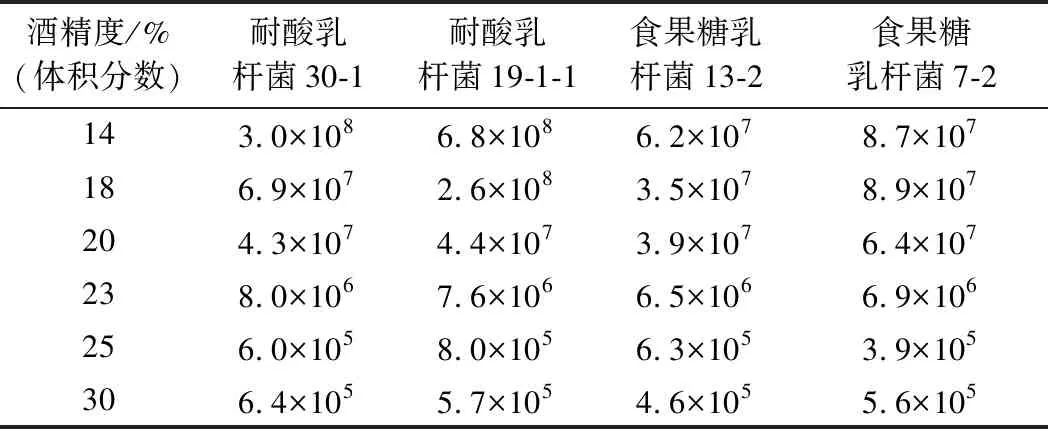
表4 不同酒精度黃酒中酸敗微生物的數(shù)量 單位:個(gè)/mL

圖6 不同酒精度正常陳釀黃酒中微生物對(duì)還原糖的消耗情況Fig.6 The consumption of reducing sugars by spoilage microbes in Huangjiu with different alcoholicity
3 結(jié)論
本研究從微生物營(yíng)養(yǎng)需求、糖分消耗、生長(zhǎng)溫度耐受、酒精耐受4個(gè)方面探究了黃酒陳釀關(guān)鍵酸敗微生物的生物學(xué)特征。黃酒酸敗微生物主要以糖類作為能源物質(zhì),能夠利用部分有機(jī)酸,不利用氨基酸,其中對(duì)葡萄糖的利用能力最強(qiáng)。黃酒酸敗微生物會(huì)利用酒體中的還原糖產(chǎn)酸,低糖條件下,酸敗微生物代謝產(chǎn)生的有機(jī)酸含量不足以導(dǎo)致黃酒發(fā)生酸敗。黃酒酸敗微生物在20~30 ℃生長(zhǎng)能力極強(qiáng),在11和40 ℃下,生長(zhǎng)受到明顯的抑制。黃酒酸敗微生物的酒精耐受性極強(qiáng),在低酒精度下,有利于酸敗微生物的生長(zhǎng);在23%(體積分?jǐn)?shù))的酒精度下,酸敗微生物仍存在生長(zhǎng)跡象;在25%(體積分?jǐn)?shù))的酒精度下,酸敗微生物的生長(zhǎng)受到抑制。因此,通過(guò)控制黃酒的糖濃度、酒精度以及儲(chǔ)存溫度可有效預(yù)防黃酒酸敗。以上對(duì)黃酒陳釀酸敗微生物生物學(xué)特征的研究,為控制黃酒酸敗微生物生長(zhǎng)提供了一定的理論基礎(chǔ)。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
