澳洲堅果MiSTPP3和MiSTPP7基因克隆與表達分析
2021-12-23 16:55:29蔡元保楊祥燕龐新華李季東林玉虹
熱帶作物學報 2021年11期
蔡元保 楊祥燕 龐新華 李季東 林玉虹



摘 ?要:以澳洲堅果品種‘O.C.’為試材,利用RT-PCR技術克隆獲得2個絲氨酸/蘇氨酸磷酸酶基因,分別命名為MiSTPP3和MiSTPP7,其編碼序列長度分別為2095 bp和1700 bp。序列分析和系統進化分析表明,MiSTPP3含有MPP_PP5_C結構域,屬于PP5亞家族;MiSTPP7含有MPP_PP7結構域,屬于PP7亞家族。蛋白理化性質和亞細胞定位分析表明,MiSTPP3是一個穩定的親水性蛋白,可能定位于質膜;MiSTPP7是一個不穩定的親水性蛋白,可能定位于線粒體基質。生物信息學分析表明,MiSTPP3和MiSTPP7都是非分泌跨膜蛋白,其二級和三級結構的主要元件都是α螺旋、無規則卷曲和延伸鏈。RT-qPCR分析表明,MiSTPP3和MiSTPP7在澳洲堅果根、莖、葉、花和小果中都有不同的表達量;MiSTPP3受低溫脅迫下調表達,而MiSTPP7上調表達,且二者都受干旱脅迫上調表達。因此,推測MiSTPP3和MiSTPP7可能參與澳洲堅果的生長發育及逆境脅迫反應。
關鍵詞:澳洲堅果;PP5;PP7;基因克隆;表達分析
中圖分類號:S664.9 ? ? ?文獻標識碼:A
Abstract: Two serine / threonine phosphatase genes were named MiSTPP3 and MiSTPP7, with coding sequences 2095 bp and 1700 bp in length, respectively, which were cloned by RT-PCR from macadamia cultivar ‘O.C.’ (Macadamia integrifolia). Sequence analysis and evolution analysis indicated that MiSTPP3 with the MPP_PP5_C domain belonged to PP5 subfamily, and MiSTPP7 with the MPP_PP7 domain belonged to PP7 subfamily. Physicochemical properties of protein and subcellular localization analysis showed that MiSTPP3 as a stable hydrophilic protein may be located in the plasma membrane, and MiSTPP7 as an unstable hydrophilic protein may be located in the mitochondrial matrix. Bioinformatics analysis indicated that MiSTPP3 and MiSTPP7 were non-secreted transmembrane proteins, α-helix, random coil and extended strand were the backbone of the secondary and 3D protein structures. RT-qPCR analysis indicated that MiSTPP3 and MiSTPP7 expressed differently in roots, stems, leaves, flowers and small fruits of M. integrifolia; MiSTPP3 was down regulated by low temperature stress, while MiSTPP7 was up-regulated, and both of them were up-regulated by drought stress. The results suggested that MiSTPP3 and MiSTPP7 may be involved in the growth and development of M. integrifolia and the stress response.
Keywords: Macadamia ternifolia; PP5; PP7; gene cloning; expression analysis
DOI: 10.3969/j.issn.1000-2561.2021.11.007
蛋白質可逆磷酸化是通過蛋白激酶和蛋白磷酸酶催化蛋白質磷酸化和去磷酸化[1-2],這一過程幾乎涉及細胞所有的生命活動,如生長發育、代謝調控、信號感知與轉導、分化與衰老等[3-6]。蛋白磷酸酶催化的去磷酸化負調控蛋白激酶的生化作用,從而抑制蛋白激酶的細胞學功能,并在細胞應答信號通路中起到重要的作用,特別對于調控植物生長發育和響應逆境脅迫具有重要意義。
蛋白磷酸酶是一個龐大的蛋白家族,主要分為絲氨酸/蘇氨酸蛋白磷酸酶(PSP)和酪氨酸蛋白磷酸酶(PTP)兩大類,根據蛋白質系統演化分析PSP大家族又分為PPP家族和PPM家族,其中PPP家族主要包括PP1、PP2A、PP2B(PP3)、PP4、PP5、PP6、PP7、SLP、Kelch、PPP Unique共10個亞家族[7-8]。這10個亞家族中,只有PP1、PP2A和PP2B的功能研究比較多,而其他亞家族的功能研究鮮有報道[9-12]。PP5和PP7的蛋白組分與結構、理化特性在進化史上的親緣關系較近[13-14],與其他亞家族存在明顯的差異。PP5蛋白通過TPR結構域,與分子伴侶Hsp90或Hsp70結合從而激活PP5蛋白的活性[15]。PP5蛋白在調控植物耐熱性、抗病性等逆境脅迫中發揮重要的作用[16-17]。PP7蛋白是植物中第一個報道與鈣調素(CaM)相互作用的絲氨酸/蘇氨酸蛋白磷酸酶,在植物耐熱性和氧化還原反應調控、光敏色素信號轉導及脅迫信號轉導中發揮重要作用[13, 18-20]。但是,目前有關PP5和PP7亞家族的功能研究并不多,有待深入研究。
澳洲堅果又稱夏威夷果,原產于亞熱帶雨林,對生長環境條件要求比較高[21-22]。低溫、干旱等逆境條件會影響澳洲堅果的正常生長發育,甚至造成澳洲堅果產量下降或死亡[23]。蛋白磷酸酶
PP5和PP7在調控植物生長發育和響應逆境脅迫中發揮關鍵的作用[14, 20],但目前國內外關于澳洲堅果PP5和PP7基因的功能研究還未見報道。因此,本研究從澳洲堅果光殼種中克隆獲得PP5基因MiSTPP3和PP7基因MiSTPP7,利用生物信息學分析其編碼蛋白的結構特征,并利用RT-qPCR分析其在不同組織和不同脅迫中的表達特性,以期為闡明澳洲堅果PP5和PP7基因在澳洲堅果生長發育及響應冷害、干旱逆境脅迫的分子機制提供科學依據。
1 ?材料與方法
1.1 ?材料
植物試材為澳洲堅果光殼種(Macadamia integrifolia)品種‘O.C.’,取自廣西農業科學院廣西亞熱帶作物研究所澳洲堅果種質資源圃。采集樹齡為5~6年的‘O.C.’植株幼根、幼莖、嫩葉、剛開放的小花及謝花后30~45 d的小果。分別對處于苗期(接穗為4個月苗齡)的嫁接苗進行4 ℃低溫處理(0.5、1、3、6、12、24 h)和20%濃度的PEG6000干旱處理(1、3、6、12、24、36 h),并采集處理后的嫩葉。其中,無脅迫處理的植株作為對照(CK),每個試驗重復3次。
1.2 ?方法
1.2.1 ?MiSTPP3和MiSTPP7基因全長cDNA克隆根據蔡元保等[24]的CTAB改良法提取澳洲堅果總RNA,并參照通用型逆轉錄試劑盒進行澳洲堅果cDNA第1鏈合成。根據本課題組從澳洲堅果轉錄組獲得的2個蛋白磷酸酶(PP5和PP7)基因,設計擴增這2個基因開放閱讀框ORF的PCR引物(表1),其中PP3-F和PP3-R擴增MiSTPP3,以及PP7-F和PP7-R擴增MiSTPP7。取1.5 μL葉片的cDNA模板進行PCR擴增:94 ℃預變性2 min;94 ℃變性35 s,53.5 ℃(MiSTPP3基因)或52.6 ℃(MiSTPP7基因)退火35 s,72 ℃延伸100 s,33個循環;72 ℃再延伸7 min,10 ℃保溫。
1.2.2 ?蛋白磷酸酶的生物信息學分析 ?利用NCBI網站的BLAST程序檢索蛋白磷酸酶的同源序列,用其保守結構域庫(CDD)分析蛋白磷酸酶的結構域,用ORF Finder分析蛋白磷酸酶的開放閱讀框ORF,用PROSITE軟件預測基元和功能結構域,用ProtParam程序分析蛋白磷酸酶的基本理化性質,用NetPhos 3.1在線程序分析蛋白質的磷酸化位點,用PSORT軟件預測亞細胞定位,用TMpred軟件和SignalP 4.1軟件分別在線預測蛋白磷酸酶的跨膜結構和信號肽,用SOPMA軟件預測蛋白質的二級結構組分及其含量,采用同源建模使用SWISS-MODEL在線工具構建蛋白質核心區域的三級結構。在NCBI網站GenBank數據庫中搜索其他植物蛋白磷酸酶的同源蛋白,采用DNAMAN 6.0軟件進行蛋白質多序列比對,并構建蛋白磷酸酶與其他同源蛋白的系統進化樹。
1.2.3 ?MiSTPP3和MiSTPP7基因的表達分析 ?采用實時熒光定量RT-qPCR方法分析澳洲堅果MiSTPP3和MiSTPP7基因的表達情況,以澳洲堅果蘋果酸脫氫酶基因MDH(GenBank登錄號:MN627210)為內參基因[25],引物為QMDH-F和QMDH-R;MiSTPP3基因引物為QPP3-F和QPP3-R;MiSTPP7基因引物為QPP7-F和QPP7-R(表1)。利用RT-qPCR分析澳洲堅果MiSTPP3和MiSTPP7基因在不同組織(根、莖、葉、花和小果)和不同脅迫下(低溫和干旱脅迫)的表達情況。RT-qPCR反應程序為:94 ℃預變性3 min;94 ℃變性20 s,56.3~58.0 ℃(表1)退火20 s,72 ℃延伸30 s,40個循環。每個試驗重復3次,根據2?ΔΔCT方法處理數據以確定MiSTPP3和MiSTPP7基因的相對表達量。
1.3 ?數據處理
利用Excel軟件對基因的相對表達量作圖,應用SPSS 18.0軟件進行數據的差異顯著性分析。
2 ?結果與分析
2.1 ?MiSTPP3和MiSTPP7基因的全長序列克隆
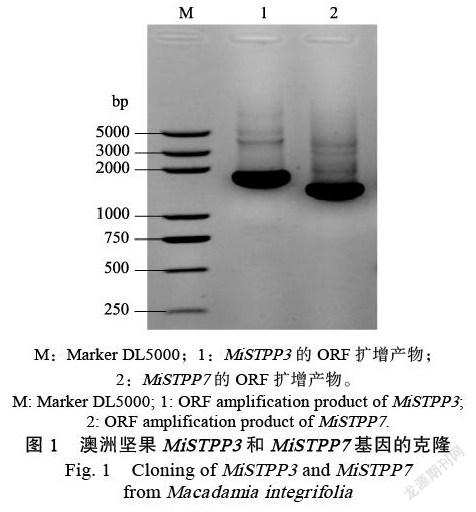
PCR擴增、核苷酸測序及蛋白同源檢索結果顯示(圖1),獲得的澳洲堅果PP3基因序列全長2095 bp,包含1455 bp的完整編碼區CDS序列,共編碼含有484個氨基酸的多肽;PP7基因序列全長1700 bp,包含1338 bp的完整編碼區CDS序列,共編碼含有445個氨基酸的多肽,其中MiSTPP3蛋白和絲氨酸/蘇氨酸磷酸酶PP5,以及MiSTPP7蛋白和絲氨酸/蘇氨酸磷酸酶PP7都具有極高的氨基酸序列相似性。因此,將這2個基因分別命名為MiSTPP3和MiSTPP7,GenBank注冊號分別為MT374550和MT374554。
2.2 ?MiSTPP3和MiSTPP7蛋白氨基酸序列比對
采用DNAMAN 6.0軟件對澳洲堅果和其他植物的蛋白磷酸酶進行蛋白質多序列比對,并用PROSITE軟件分析其功能結構域。分析結果顯示(圖2),澳洲堅果MiSTPP3與葡萄VvPP5、玫瑰木屬植物RaPP5和辣椒CaPP5的氨基酸序列同源性分別為90.68%、89.63%和89.23%,都含有PP5亞家族的典型結構域MPP_PP5_C和N端的TPR結構域;MiSTPP7與橡膠樹HbPP7、麻風樹JcPP7和水芙蓉NnPP7的氨基酸序列同源性分別為77.48%、77.48%和77.38%,都含有PP7亞家族的典型結構域MPP_PP7。
2.3 ?MiSTPP3和MiSTPP7與其他蛋白磷酸酶的系統發育分析
選擇澳洲堅果MiSTPP3和MiSTPP7蛋白與不同物種的代表性蛋白磷酸酶構建系統發育樹。結果顯示(圖3),25個絲氨酸/蘇氨酸蛋白磷酸酶被聚成PP1、PP2A、PP2B(PP3)、PP5和PP7共5個亞家族,其中澳洲堅果MiSTPP3蛋白屬于PP5亞家族,與同一亞家族的玫瑰木屬植物RaPP5和葡萄VvPP5的親緣關系最近;MiSTPP7蛋白屬于PP7亞家族,與同一亞家族的水芙蓉NnPP7和棗ZjPP7的親緣關系最近;而且,在這5個亞家族中,PP5和PP7亞家族的親緣關系最近,PP5和PP7亞家族又與PP1亞家族的親緣關系最近,推測具有相同的進化起源和相似的生物學功能。
2.4 ?MiSTPP3和MiSTPP7蛋白的一級結構和理化性質分析
MiSTPP3蛋白的一級結構和理化性質分析結果顯示,其分子質量為54.58 kDa,分子式為C2449H3786N646O719S25,理論等電點(pI)為5.81,帶負電荷殘基數(Asp+Glu)為69個,帶正電荷殘基數(Arg+Lys)為58個,不穩定系數為35.42,為穩定蛋白。脂肪族指數為79.79,親水性系數GRAVY預測為負值(?0.287),為親水性蛋白。因此,MiSTPP3蛋白是一個穩定的親水性蛋白。
MiSTPP7蛋白的一級結構和理化性質分析結果顯示,其分子質量為49.30 kDa,分子式為C2211H3433N585O665S14,理論等電點(pI)為5.50,帶負電荷殘基數(Asp+Glu)為57個,帶正電荷殘基數(Arg+Lys)為47個,不穩定系數為41.81,為不穩定蛋白。脂肪族指數為81.46,親水性系數GRAVY預測為負值(?0.319),為親水性蛋白。因此,MiSTPP7蛋白是一個不穩定的親水性蛋白。
2.5 ?MiSTPP3和MiSTPP7蛋白的基本功能分析
Net Phos 3.1軟件預測表明,MiSTPP3蛋白有41個磷酸化位點,包括22個絲氨酸,8個蘇氨酸,11個酪氨酸。PSORT軟件預測表明,MiSTPP3蛋白定位于質膜可能性最高,為70%。SignalP4.1和TMpred軟件預測顯示,MiSTPP3蛋白不存在信號肽,有2個跨膜結構域(262~286位,311~331位氨基酸),故MiSTPP3為非分泌跨膜蛋白。
Net Phos 3.1軟件預測表明,MiSTPP7蛋白有50個磷酸化位點,包括32個絲氨酸,11個蘇氨酸,7個酪氨酸。PSORT軟件預測表明,MiSTPP7蛋白定位于線粒體基質可能性最高,為46.8%。SignalP和TMpred軟件預測顯示,MiSTPP7蛋白不存在信號肽,有3個跨膜結構域(76~99位,155~177位,225~242位氨基酸),故MiSTPP7為非分泌跨膜蛋白。
2.6 ?MiSTPP3和MiSTPP7蛋白的二級和三級結構預測
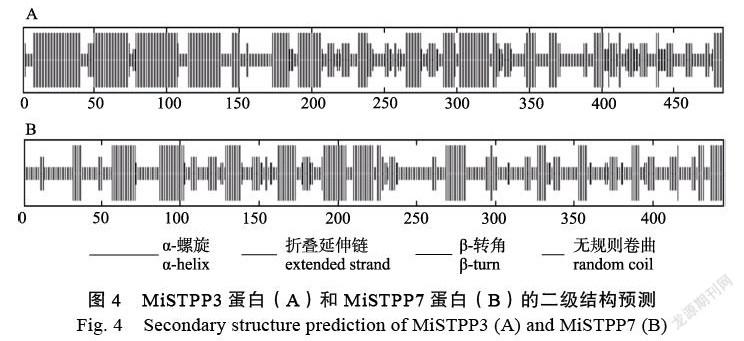
SOPMA軟件預測顯示,澳洲堅果MiSTPP3蛋白二級結構的主要結構元件是α-螺旋、無規則卷曲、延伸鏈和β-轉角,分別占46.28%、30.99%、15.70%和7.02%,α-螺旋主要集中在N端(圖4A);MiSTPP7蛋白二級結構的主要結構元件是無規則卷曲、α-螺旋、延伸鏈和β-轉角,分別占46.74%、30.34%、16.40%和6.52%(圖4B)。
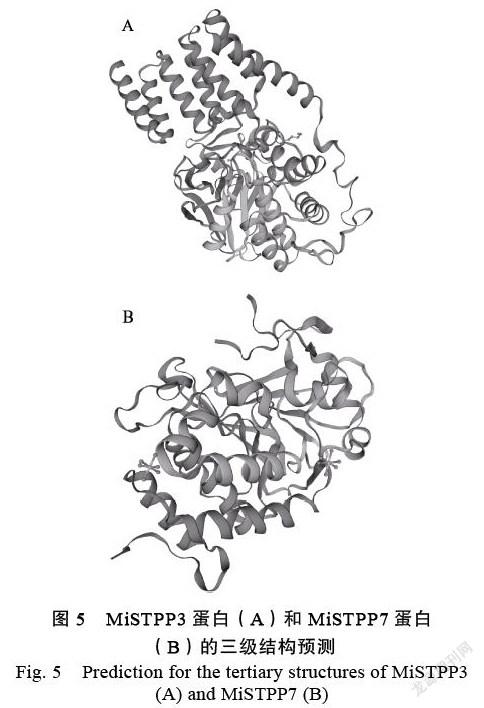
以絲氨酸/蘇氨酸磷酸酶PP5的晶體結構(PDB ID:5obl.1.A)為同源模板,利用在線工具SWISS-MODEL預測MiSTPP3蛋白的三級結構。結果表明,MiSTPP3蛋白的氨基酸序列與模板序列一致性為85.74%,GMQE值為0.98,同源建模區域的組成以α-螺旋和無規卷曲為主,以延伸鏈為輔(圖5A)。目前,SWISS-MODEL軟件的蛋白數據庫中還沒有絲氨酸/蘇氨酸磷酸酶PP7的晶體結構,只能以親緣關系最近的PP5晶體結構(PDB ID:5hpe.1.A)為同源模板,利用SWISS-MODEL在線工具構建MiSTPP7蛋白的三級結構。結果表明,MiSTPP7蛋白的氨基酸序列與模板序列一致性為36.94%,GMQE值為0.55,同源建模區域主要由無規卷曲和α-螺旋組成(圖5B)。與MiSTPP3和MiSTPP7蛋白的二級結構預測結果相符合。
2.7 ?MiSTPP3和MiSTPP7基因的表達分析
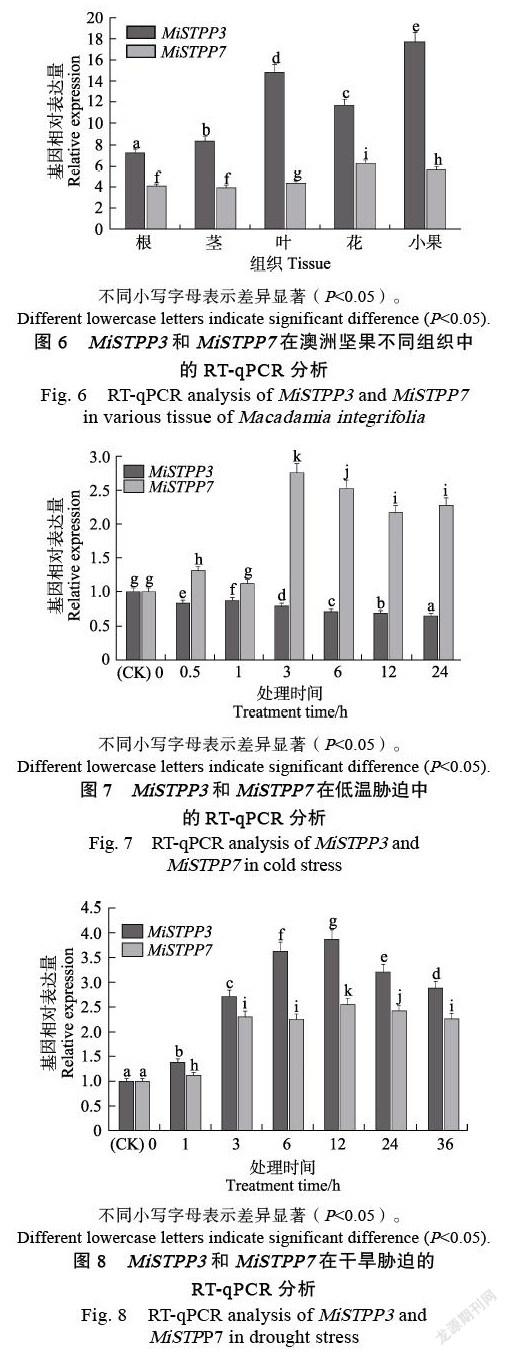
MiSTPP3和MiSTPP7基因在不同組織的表達分析顯示(圖6),MiSTPP3和MiSTPP7在澳洲堅果根、莖、葉、花和小果中呈現顯著性差異表達,其中MiSTPP3在小果中的表達量最高,MiSTPP7在花中的表達量最高。4 ℃低溫脅迫的表達分析顯示(圖7),澳洲堅果苗期受低溫脅迫后MiSTPP3在葉片中表達顯著性下調;而MiSTPP7表達顯著上調,且脅迫3 h后的表達量達到峰值。PEG模擬干旱脅迫的表達分析顯示(圖8),澳洲堅果苗期受干旱脅迫后葉片中MiSTPP3 和MiSTPP7的表達都顯著上調,且在脅迫處理12 h時達到峰值。
3 ?討論
目前,PP5和PP7亞家族基因在動物中的研究較多,在細胞生長發育調控、響應逆境脅迫以及細胞應答信號通路中扮演重要角色[26-27],但在植物中這2個亞家族基因的鑒定、結構及功能研究較少報道。PP5和PP7是PPP家族的獨特成員[7-8],MiSTPP3和MiSTPP7分別作為PP5和PP7亞家族的新成員,分別含有典型結構域MPP_PP5_C和MPP_PP7。
蛋白去磷酸化是蛋白可逆磷酸化修飾的2個關鍵步驟之一[5, 28],MiSTPP3和MiSTPP7蛋白在發揮去磷酸化作用的過程中主要以絲氨酸進行磷酸化修飾。植物PP5與PP7蛋白的亞細胞定位取決于mRNA前體的選擇性剪接[29]。MiSTPP3和MiSTPP7作為非分泌跨膜蛋白,分別定位于質膜和線粒體基質,至于其亞細胞定位的mRNA前體剪接機制還有待于深入研究。
PP5與PP7蛋白在進化過程中都具有保守的氨基酸序列,二者親緣關系相對于其他亞家族最為相近[14],具有相同的進化起源和相似的生物學功能。已有研究表明,這2個亞家族基因在調控組織的生長發育中發揮重要作用,如調控擬南芥氣孔的生長發育[29]。MiSTPP3和MiSTPP7可能參與澳洲堅果不同組織(如根、莖、葉、花和小果等)的生長發育,尤其在調控花和小果生長發育中發揮重要作用。
分子伴侶Hsp90是PP5蛋白催化的去磷酸化的關鍵酶,兩者的結合可調控脅迫誘導的細胞信號轉導[14]。如熱激脅迫條件下,擬南芥AtPP5與底物MDH相互作用,與AtHsp90結合形成復合體從而增強植株的耐熱性[16];番茄R蛋白I-2通過與PP5和Hsp90的直接相互作用,增強了植株對尖孢鐮刀菌的抗性[30]。MiSTPP3受低溫誘導后顯著下調表達,受干旱誘導后顯著上調表達,這2種不同的表達模式都證實該基因參與澳洲堅果逆境脅迫反應。但是,MiSTPP3是否與Hsp90結合從而響應這些逆境脅迫反應還需深入研究。
植物PP7蛋白在光和脅迫信號傳遞途徑的交叉點上發揮了重要的調控作用[20]。擬南芥AtPP7除了調控氧化還原反應外[13],還與鈣調素(CaM)結合從而增強植株的耐熱性[18],且作為正調控因子調控其光敏色素信號轉導[19,31]。與已知的PP7亞家族基因一樣,MiSTPP7受低溫和干旱誘導后都顯著上調表達。因此,推測MiSTPP7可能作為正調控因子,參與澳洲堅果逆境脅迫的信號轉導。
本研究結果表明,MiSTPP3和MiSTPP7可能參與了澳洲堅果植株生長發育和逆境脅迫響應。在后續的工作中,利用轉基因技術對澳洲堅果MiSTPP3和MiSTPP7基因功能的深層次研究,有助于闡明這2個基因在響應逆境脅迫的生物學功能和作用機制,尤其是對于揭示植物PP5和PP7亞家族基因的信號轉導網絡及其亞家族間的相互關系具有重要作用。
參考文獻
[1] Macek B, Mann M, Olsen J V. Global and site-specific quantitative phosphoproteomics: principles and applica-tions[J]. Annual Review of Pharmacology and Toxicology, 2009, 49(1): 199-221.
[2] Goel R, Harsha H C, Pandey A, et al. Human protein refer-ence database and human proteinpedia as resources for phosphoproteome analysis[J]. Molecular BioSystems, 2012, 8(2): 453-463.
[3] Ptacek J, Devgan G, Michaud G, et al. Global analysis of protein phosphorylation in yeast[J]. Nature, 2005, 438(7068): 679-684.
[4] Aivaliotis M, Macek B, Gnad F, et al. Ser/thr/Tyr protein phosphorylation in the archaeon Halobacterium salina-rum— A representative of the third domain of life[J]. PLoS One, 2009, 4(3): e4777.
[5] Barrero-Gil J, Salinas J. Post-translational regulation of cold acclimation response[J]. Plant Science, 2013, 205/206: 48- 54.
[6] 肖 ?冬, 崔燕嬌, 王寧寧. 葉片衰老過程中的蛋白激酶和蛋白磷酸酶[J]. 植物生理學報, 2014, 50(9): 1267-1273.
[7] Cohen P. The structure and regulation of protein phospha-tases[J]. Annual Review of Biochemistry, 1989, 58(1): 453- 508.
[8] Owens D M, Keyse S M. Differential regulation of MAP kinase signalling by dual-specificity protein phosphatases[J]. Oncogene, 2007, 26(22): 3203-3213.
[9] Durian G, Rahikainen M, Alegre S, et al. Protein phospha-tase 2A in the regulatory network underlying biotic stress resistance in plants[J]. Frontiers in Plant Science, 2016, 7: 812.
[10] Khan Z H, Agarwal S, Rai A, et al. Co-expression network analysis of protein phosphatase 2A (PP2A) genes with stress-responsive genes in Arabidopsis thaliana reveals 13 key regulators[J]. Scientific Reports, 2020, 10(1): 1-16.
[11] Tan S T, Abas M, Verstraeten I, et al. Salicylic acid targets protein phosphatase 2A to attenuate growth in plants[J]. Current Biology, 2020, 30(3): 381-395.e8.
[12] Casamayor A, Ari?o J. Controlling Ser/Thr protein phosphatase PP1 activity and function through interaction with regulatory subunits[M]//Advances in Protein Chemistry and Structural Biology. Amsterdam: Elsevier, 2020: 231-288.
[13] Andreeva A V, Solov'eva O V, Kakuev D L, et al. Purifica-tion of plant protein phosphatase PP7 and evidence for its redox regulation[J]. Archives of Biochemistry and Biophysics, 2001, 396(1): 65-70.
[14] Oberoi J, Dunn D M, Woodford M R, et al. Structural and functional basis of protein phosphatase 5 substrate specificity[J]. PNAS, 2016, 113(32): 9009-9014.
[15] Connarn J N, Assimon V A, Reed R A, et al. The molecular chaperone Hsp70 activates protein phosphatase 5 (PP5) by binding the tetratricopeptide repeat (TPR) domain[J]. Journal of Biological Chemistry, 2014, 289(5): 2908-2917.
[16] Park J H, Kim W Y, Chae H B, et al. Serine/threonine pro-tein phosphatase 5 (PP5) interacts with substrate under heat stress conditions and forms protein complex in Arabidop-sis[J]. Plant Signaling & Behavior, 2012, 7(5): 535-538.
[17] Van Bentem S D L F, Vossen J H, Vries K J, et al. Heat shock protein 90 and its co-chaperone protein phosph?atase 5 interact with distinct regions of the tomato I-2 disea?se resistance protein[J]. Plant Journal, 2005, 43(2): 284-298.
[18] Liu H T, Li G L, Chang H, et al. Calmodulin-binding protein phosphatase PP7 is involved in thermotolerance in Arabidopsis[J]. Plant Cell & Environment, 2007, 30(2): 156-164.
[19] Genoud T, Trevi?o Santa Cruz M, Kulisic T, et al. The protein phosphatase 7 regulates phytochrome signaling in Arabidopsis[J]. PLoS One, 2008, 3(7): e2699.
[20] Andreeva A V, Kutuzov M A. PPEF/PP7 protein Ser/Thr phosphatases[J]. Cellular and Molecular Life Sciences, 2009, 66(19): 3103-3110.
[21] Buthelezi N M D, Magwaza L S, Tesfay S Z. Postharvest pre-storage processing improves antioxidants, nutritional and sensory quality of Macadamia nuts[J]. Scientia Horticulturae, 2019, 251: 197-208.
[22] Herbert S W, Walton D A, Wallace H M. The influence of pollen-parent and carbohydrate availability on Macadamia yield and nut size[J]. Scientia Horticulturae, 2019, 251: 241-246.
[23] 蔡元保, 楊祥燕, 陳顯國, 等. 澳洲堅果SCoT反應體系的建立及應用[J]. 熱帶亞熱帶植物學報, 2013, 21(3): 253- 258.
[24] 蔡元保, 楊祥燕, 孫光明, 等. 菠蘿花發育相關基因AcMADS1的克隆與組織表達特性分析[J]. 植物學報, 2014, 49(6): 692-703.
[25] 楊 ?倩, 楊子平, 周婭麗, 等. 澳洲堅果實時熒光定量PCR分析中內參基因的篩選[J]. 熱帶作物學報, 2020, 41(8): 1505-1512.
[26] Kostich M, English J, Madison V, et al. Human members of the eukaryotic protein kinase family[J]. Genome Biology, 2002, 3(9): research0043.1.
[27] 鄒 ?奕, 閆彩燕, 邳 ?植. 甜菜蛋白磷酸酶超家族分析[J]. 中國糖料, 2018, 40(6): 19-22.
[28] 阮班軍, 代 ?鵬, 王 ?偉, 等. 蛋白質翻譯后修飾研究進展[J]. 中國細胞生物學學報, 2014, 36(7): 1027-1037.
[29] Van Bentem S D L F, Vossen J H, Vermeer J E M, et al. The subcellular localization of plant protein phosphatase 5 isoforms is determined by alternative splicing[J]. Plant Physiology, 2003, 133(2): 702-712.
[30] Sun X D, Kang X J, Ni M. Hypersensitive to red and blue 1 and its modification by protein phosphatase 7 are im-plicated in the control of Arabidopsis stomatal aperture[J]. PLoS Genetics, 2012, 8(5): e1002674.
[31] M?ller S G, Kim Y S, Kunkel T, et al. PP7 is a positive regulator of blue light signaling in Arabidopsis[J]. Plant Cell, 2003, 15(5): 1111-1119.
責任編輯:黃東杰
