木薯MeSWEET10a基因啟動(dòng)子EBE區(qū)編輯載體的構(gòu)建及驗(yàn)證
2021-12-23 16:55:29張彤王亞杰郭文雅李瑞梅劉姣郭建春胡新文姚遠(yuǎn)耿夢(mèng)婷
熱帶作物學(xué)報(bào) 2021年11期
張彤 王亞杰 郭文雅 李瑞梅 劉姣 郭建春 胡新文 姚遠(yuǎn) 耿夢(mèng)婷

摘 ?要:木薯細(xì)菌性枯萎病(cassava bacterial blight, CBB)是由地毯草黃單胞木薯萎蔫致病變種(Xanthomonas axonopodis pv. Manihotis, Xam)引起的木薯重要病害,影響木薯產(chǎn)量,我國(guó)主栽的木薯品種均不抗Xam。Xam分泌的TAL20效應(yīng)蛋白可結(jié)合木薯MeSWEET10a基因啟動(dòng)子的EBE區(qū),激活該基因表達(dá),促進(jìn)糖類運(yùn)輸至Xam侵染的部位,為病菌的繁殖提供碳水化合物。本研究利用在線軟件CRISPR-P v2.0設(shè)計(jì)EBE區(qū)的基因編輯靶點(diǎn),構(gòu)建CRISPR/Cas9基因編輯質(zhì)粒pCAMBIA1301-Cas9-EBE-sgRNA。將重組質(zhì)粒轉(zhuǎn)化LBA4404根癌農(nóng)桿菌,利用農(nóng)桿菌介導(dǎo)法侵染木薯脆性胚性愈傷組織。提取侵染和未侵染的木薯脆性胚性愈傷組織DNA,PCR擴(kuò)增靶點(diǎn)EBE區(qū)及潛在的脫靶位點(diǎn),并進(jìn)行Sanger測(cè)序分析編輯效果。結(jié)果發(fā)現(xiàn)EBE區(qū)被成功編輯,且沒(méi)有脫靶現(xiàn)象。本研究有助于進(jìn)一步獲得MeSWEET10a基因啟動(dòng)子EBE區(qū)域的木薯突變體,以增強(qiáng)木薯抗細(xì)菌性枯萎病的能力。
關(guān)鍵詞:木薯;細(xì)菌性枯萎病;MeSWEET10a;CRISPR/Cas9
中圖分類號(hào):S533 ? ? ?文獻(xiàn)標(biāo)識(shí)碼:A
Abstract: Cassava bacterial blight (CBB) is an important cassava disease caused by Xanthomonas axonopodis pv. manihotis (Xam), which affects the yield of cassava. The main cassava varieties in China are not resistant to Xam. The TAL20 effector protein secreted by Xam, could bind to the EBE region of the cassava MeSWEET10a promoter, activate the gene expression, promote the transport of sugars to the site of Xam infection, and provide carbohydrates for the reproduction of the pathogen. In this study, the gene editing target of EBE region was designed by using online software CRISPR-Pv2.0, and the CRISPR/Cas9 gene editing plasmid pCAMBIA1301-Cas9-EBE-sgRNA was constructed. The recombinant plasmid was transformed into Agrobacterium tumefaciens LBA4404, and the fragile embryogenic calli of cassava was infected by the A. tumefaciens-mediated method. DNA was extracted from the fragile embryogenic calli of infected and uninfected cassava. The target EBE region and potential miss sites were amplified by PCR, and the editing effect was analyzed by Sanger sequencing. It was found that the EBE region was edited successfully without off-target phenomenon. This study is helpful to further obtain cassava mutants in the EBE region of MeSWEET10a promoter and enhance the resistance of cassava to bacterial blight.
Keywords: cassava; bacterial blight; MeSWEET10a; CRISPR/Cas9
DOI: 10.3969/j.issn.1000-2561.2021.11.010
木薯是熱帶地區(qū)重要的糧食作物,而病害是木薯產(chǎn)量提高的重要制約因素之一[1-2]。木薯細(xì)菌性枯萎病(cassava bacterial blight, CBB)是我國(guó)木薯面臨的主要病害,該病是由地毯草黃單胞菌木薯萎蔫致病變種(Xanthomonas axonopodis pv. Manihotis, Xam)引起的細(xì)菌性病害[3]。我國(guó)木薯主栽品種均不抗Xam病原菌的侵染,CBB曾導(dǎo)致我國(guó)木薯大面積減產(chǎn)。因此,培育抗CBB木薯新品種有益于我國(guó)木薯產(chǎn)業(yè)持續(xù)健康發(fā)展。
Xam病原菌侵染木薯時(shí),可通過(guò)Ⅲ型分泌系統(tǒng)將轉(zhuǎn)錄激活因子樣效應(yīng)物蛋白(transcription activator-like effectors,TAL)分泌到宿主細(xì)胞中[4],TAL效應(yīng)蛋白的可變重復(fù)區(qū)(repeat variable diresidues,RVD)識(shí)別宿主感病相關(guān)基因啟動(dòng)子上的效應(yīng)蛋白結(jié)合元件(effector-binding element,EBE),并激活下游感病基因表達(dá),以促進(jìn)Xam病原菌的侵染[5]。不同的Xam病原菌菌株包含的TAL效應(yīng)蛋白數(shù)目存在差異,一般為1~5個(gè)。其中TAL20的RVD區(qū)可識(shí)別木薯糖轉(zhuǎn)運(yùn)蛋白編碼基因MeSWEET10a啟動(dòng)子的EBE區(qū),激活MeSWEET10a基因的表達(dá),以利于糖類物質(zhì)轉(zhuǎn)運(yùn)至Xam病原菌侵染部位,以促進(jìn)病原菌的繁殖[6]。因此,MeSWEET10a基因啟動(dòng)子的EBE區(qū)是利用基因編輯技術(shù)創(chuàng)制抗CBB木薯種質(zhì)的重要靶標(biāo)。類似的抗病育種策略已應(yīng)用水稻抗白葉枯病育種。利用基因編輯技術(shù),編輯水稻SWEET基因啟動(dòng)子EBE區(qū),阻止水稻白葉枯病病原菌水稻黃單胞菌水稻致病變種(Xanthomonas oryzae pv. oryzae,Xoo)的TAL效應(yīng)蛋白激活水稻SWEET基因,可獲得廣譜抗白葉枯病的水稻種質(zhì)[7]。本研究利用CRISPR/Cas9技術(shù)構(gòu)建木薯糖轉(zhuǎn)運(yùn)蛋白基因MeSWEET10a啟動(dòng)子EBE區(qū)域的編輯載體,驗(yàn)證了載體的編輯效果,及分析了脫靶的可能性,為進(jìn)一步獲得MeSWEET10a啟動(dòng)子突變體,創(chuàng)制抗細(xì)菌性枯萎病木薯新種質(zhì)奠定了基礎(chǔ)。
1 ?材料與方法
1.1 ?材料
植物材料:‘華南8號(hào)’(SC8)木薯品種來(lái)源于中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶作物品種資源研究所。
菌株及質(zhì)粒:大腸桿菌菌株DH5α、根癌農(nóng)桿菌菌株LBA4404、基因編輯質(zhì)粒pCAMBIA1301- Cas9-sgRNA為本實(shí)驗(yàn)室構(gòu)建。
試劑:T4 DNA ligase、DNA聚合酶Ex Taq采購(gòu)于Takara公司,BsaⅠ限制性內(nèi)切酶購(gòu)于NEB公司,質(zhì)粒提取試劑盒購(gòu)生工生物工程(上海)股份有限公司,植物DNA提取試劑盒購(gòu)于成都福際生物技術(shù)有限公司。
1.2 ?方法
1.2.1 ?靶點(diǎn)引物的設(shè)計(jì)及基因編輯載體構(gòu)建 ?根據(jù)MeSWEET10a(Manes.06G123400)基因的啟動(dòng)子序列,利用CRISPR-P v2.0在線軟件分析MeSWEET10a啟動(dòng)子EBE區(qū)的靶點(diǎn)及潛在脫靶位點(diǎn)。合成靶點(diǎn)退火引物(表1),參照李嶄等[8]的方法進(jìn)行退火及克隆至基因編輯載體。菌液PCR篩選陽(yáng)性克隆后送生工生物工程有限公司測(cè)序,將正確的質(zhì)粒命名為pCAMBIA1301-Cas9- EBE-sgRNA,并將該質(zhì)粒轉(zhuǎn)化LBA4404農(nóng)桿菌,菌液PCR驗(yàn)證正確后備用。
1.2.2 ?木薯脆性胚性愈傷組織的誘導(dǎo)及侵染 ?參照李嶄等[8]的方法誘導(dǎo)‘SC8’木薯脆性胚性愈傷組織,將攜帶基因編輯質(zhì)粒的LBA4404農(nóng)桿菌侵染脆性胚性愈傷組織,經(jīng)過(guò)共培養(yǎng)3 d,洗菌繼續(xù)培養(yǎng)21 d后用于檢測(cè)EBE區(qū)的編輯效果。
1.2.3 ?EBE區(qū)編輯效果及脫靶分析 ?設(shè)計(jì)MeSW??-E?ET10a啟動(dòng)子EBE區(qū)編輯靶點(diǎn)和4個(gè)潛在脫靶位點(diǎn)區(qū)域的PCR擴(kuò)增引物(表1)。提取農(nóng)桿菌侵染的木薯脆性胚性愈傷組織DNA,PCR擴(kuò)增獲得目的片段并測(cè)序。基于Sanger測(cè)序峰值圖,分析啟動(dòng)子EBE區(qū)編輯效果,以及是否存在脫靶情況。
2 ?結(jié)果與分析
2.1 ?啟動(dòng)子編輯靶點(diǎn)的設(shè)計(jì)及潛在脫靶位點(diǎn)分析
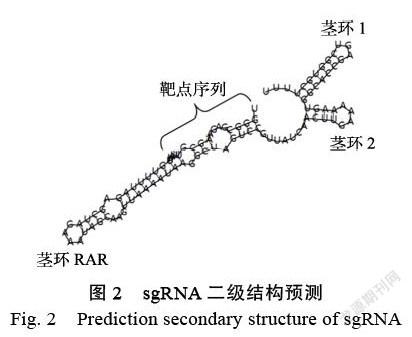
通過(guò)在線設(shè)計(jì)軟件CRISPR-P v2.0,根據(jù)木薯MeSWEET10a基因啟動(dòng)子EBE區(qū)序列,設(shè)計(jì)獲得編輯MeSWEET10a基因啟動(dòng)子EBE區(qū)域的靶點(diǎn)序列,該靶點(diǎn)CG含量值為45%,PAM序列為CCT(圖1)。脫靶位點(diǎn)預(yù)測(cè)分析結(jié)果顯示(表2),該靶點(diǎn)在木薯基因組存在4個(gè)潛在脫靶位點(diǎn)。其中,Off-target-1的脫靶分值為0.587,有3個(gè)錯(cuò)配的堿基數(shù),有可能脫靶到14號(hào)染色體;Off-target-2的脫靶分值為0.142,有4個(gè)錯(cuò)配的堿基數(shù),位于15號(hào)染色體;Off-target-3的脫靶分值為0.102,?有3個(gè)錯(cuò)配的堿基數(shù),位于10號(hào)染色體;Off-target-4的脫靶分值為0.098,位于10號(hào)染色體。全部潛在脫靶位點(diǎn)均不屬于基因的編碼區(qū),表明即使脫靶也不會(huì)造成基因功能的缺失。sgRNA的二級(jí)結(jié)構(gòu)預(yù)測(cè)分析結(jié)果顯示(圖2),靶點(diǎn)序列二級(jí)結(jié)構(gòu)較為松散,有利于結(jié)合靶標(biāo)位置。
2.2 ?基因編輯載體的構(gòu)建
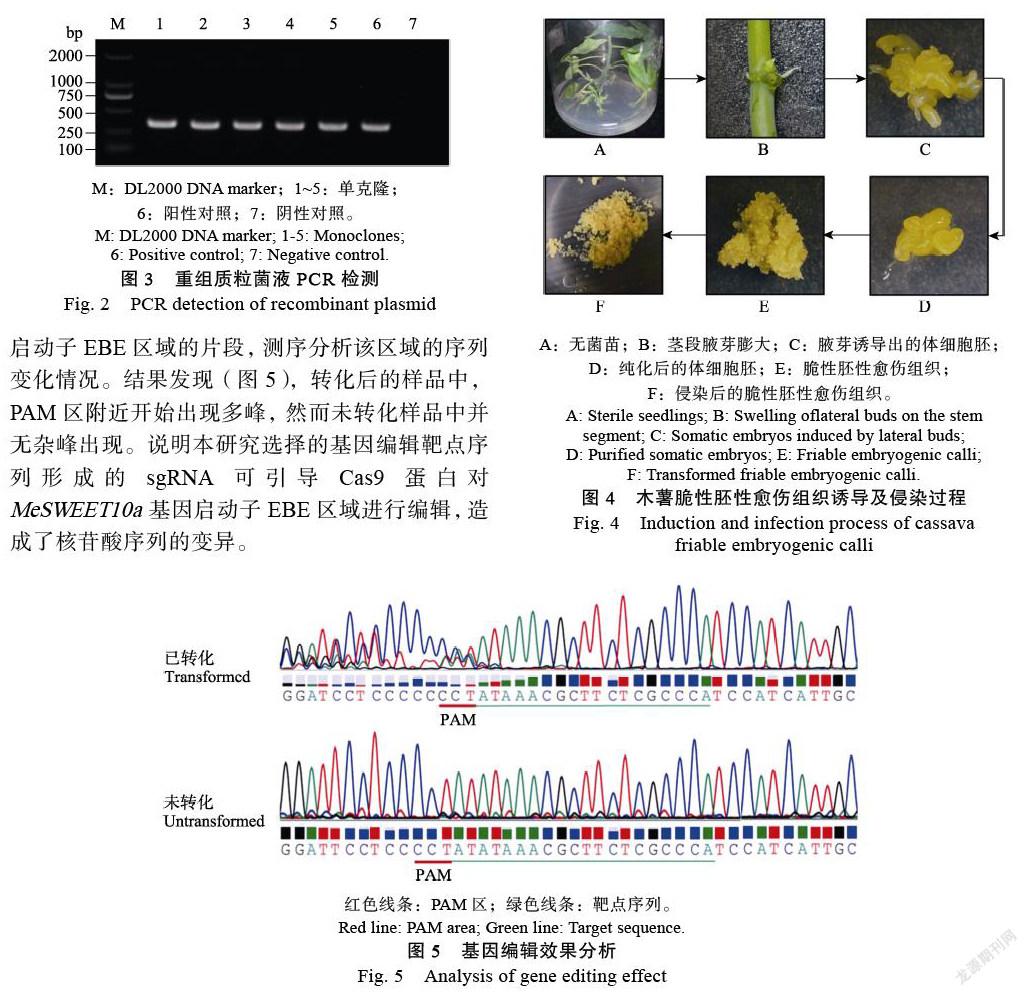
本研究所使用的基因編輯載體為pCAMBIA 1301-Cas9-sgRNA,采用線性化的質(zhì)粒與退火后的EBE區(qū)靶點(diǎn)引物連接,構(gòu)建AtU6-26-EBE- sgRNA表達(dá)盒。通過(guò)菌液PCR篩選陽(yáng)性大腸桿菌克隆,獲得約350 bp目的條帶(圖3)。測(cè)序分析進(jìn)一步證明基因編輯載體pCAMBIA1301- Cas9-EBE-sgRNA已構(gòu)建成功。提取質(zhì)粒后,轉(zhuǎn)LBA4404農(nóng)桿菌備用。
2.3 ?編輯載體轉(zhuǎn)化木薯脆性胚性愈傷組織及編輯效果驗(yàn)證
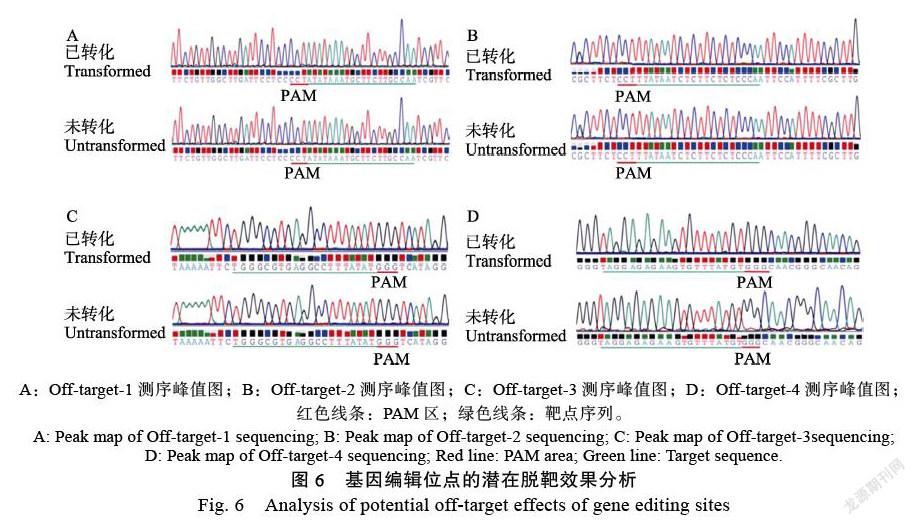
以‘SC8’木薯無(wú)菌苗為外植體,誘導(dǎo)腋芽膨大、然后誘導(dǎo)體細(xì)胞胚形成,最終誘導(dǎo)獲得脆性胚性愈傷組織(圖4)。將攜帶pCAMBIA1301- Cas9-EBE-sgRNA質(zhì)粒的LBA4404轉(zhuǎn)化木薯脆性胚性愈傷組織21 d,PCR擴(kuò)增MeSWEET10a基因啟動(dòng)子EBE區(qū)域的片段,測(cè)序分析該區(qū)域的序列變化情況。結(jié)果發(fā)現(xiàn)(圖5),轉(zhuǎn)化后的樣品中,PAM區(qū)附近開始出現(xiàn)多峰,然而未轉(zhuǎn)化樣品中并無(wú)雜峰出現(xiàn)。說(shuō)明本研究選擇的基因編輯靶點(diǎn)序列形成的sgRNA可引導(dǎo)Cas9蛋白對(duì)MeSWEET10a基因啟動(dòng)子EBE區(qū)域進(jìn)行編輯,造成了核苷酸序列的變異。
2.4 ?脫靶分析
以轉(zhuǎn)化和未轉(zhuǎn)化基因編輯載體的木薯脆性胚性愈傷組織DNA為模板,PCR擴(kuò)增MeSWEET10a基因啟動(dòng)子EBE區(qū)編輯靶點(diǎn)的4個(gè)潛在脫靶位點(diǎn)區(qū)域片段,Sanger測(cè)序分析該靶點(diǎn)是否有脫靶的現(xiàn)象。結(jié)果表明(圖6),潛在脫靶位點(diǎn)的序列在已轉(zhuǎn)化和未轉(zhuǎn)化的樣品中沒(méi)有發(fā)生改變。說(shuō)明本研究構(gòu)建的基因編輯載體pCAMBIA1301- Cas9-EBE-sgRNA可對(duì)MeSWEET10a基因啟動(dòng)子EBE區(qū)進(jìn)行特異編輯,并且無(wú)脫靶現(xiàn)象。
3 ?討論
植物病原微生物引起的病害對(duì)作物的產(chǎn)量構(gòu)成嚴(yán)重威脅。病原微生物利用宿主的易感(S)基因來(lái)促進(jìn)增殖,敲除或降低S基因可能會(huì)干擾宿主和病原體之間的親和性,從而為宿主提供廣譜和持久的抗病能力。利用基因組編輯技術(shù)編輯作物S基因的編碼區(qū)或其啟動(dòng)子,提高抗病性的報(bào)道主要集中于水稻、小麥、西紅柿、柑橘等作物。霉菌抗性位點(diǎn)O(mildew resistance locus O, MLO)的突變可賦予植物對(duì)白粉病的抗病性,如編輯番茄SlMlo1基因[9],小麥TaMLO-A1、TaMLO-B1、TaMLO-D1可提高小麥和番茄的抗白粉病能力[10]。小麥TaEDR1的3個(gè)同源基因被編輯突變,可增強(qiáng)小麥對(duì)白粉病的抗病能力[11]。轉(zhuǎn)錄因子CsLOB1在促進(jìn)柑橘潰瘍病病菌生長(zhǎng)和膿包形成過(guò)程中起關(guān)鍵作用,編輯CsLOB1基因增強(qiáng)柑橘抗柑橘潰瘍病的能力[12]。在敲除S基因啟動(dòng)子關(guān)鍵元件,干擾病原微生物對(duì)作物S基因表達(dá)的調(diào)控,可創(chuàng)制抗病種質(zhì)。水稻白葉枯病S基因OsSWEET11、OsSWEET14和OsSWEET13啟動(dòng)子的EBE區(qū)敲除,阻止TAL效應(yīng)蛋白激活這些S基因表達(dá),賦予水稻對(duì)白葉枯病原菌的廣譜抗病能力[13]。
效應(yīng)蛋白在病原菌與植物的互作過(guò)程中扮演著重要的作用,病原菌分泌的效應(yīng)蛋白可調(diào)控宿主感病相關(guān)基因的表達(dá),干擾植物細(xì)胞的正常活動(dòng),創(chuàng)造病原菌侵染和繁殖的有利條件,同時(shí)植物可協(xié)同進(jìn)化,獲得識(shí)別和監(jiān)控效應(yīng)蛋白的能力,從而激活植物的免疫反應(yīng),增強(qiáng)宿主的抗病性。病原菌通過(guò)效應(yīng)蛋白結(jié)合植物的SWEET基因啟動(dòng)子,劫持SWEET基因?yàn)椴≡陨淼纳L(zhǎng)提供養(yǎng)分。白粉菌浸染擬南芥后誘導(dǎo)AtSWEET12基因表達(dá)升高,灰霉菌侵染后則誘導(dǎo)AtSWEET4、AtSWEET15、AtSWEET17;灰霉菌侵染葡萄誘導(dǎo)VvSWEET4基因表達(dá)量升高[14-16]。木薯Xam病原菌分泌的TAL20效應(yīng)蛋白,結(jié)合MeSWEET10a基因啟動(dòng)子的EBE區(qū),調(diào)控該基因表達(dá)。為了創(chuàng)制抗CBB的木薯新種質(zhì),本研究成功構(gòu)建了EBE區(qū)的CRISPR/Cas9編輯載體。李嶄等[8]證明了利用基因編輯載體,遺傳轉(zhuǎn)化‘SC8’木薯脆性胚性愈傷組織,可快速檢測(cè)淀粉合成酶基因SSIII-1和SSIII-2編碼區(qū)的編輯效果。本研究證明脆性胚性愈傷組織轉(zhuǎn)化檢測(cè)方式,同樣適用于檢測(cè)GC含量較低的MeSWEET10a基因啟動(dòng)子EBE區(qū)的編輯效果。脫靶是CRISPR/Cas9基因編輯技術(shù)面臨的一個(gè)重要問(wèn)題。本研究設(shè)計(jì)的EBE區(qū)編輯靶點(diǎn)在木薯基因組中存在4個(gè)潛在脫靶位點(diǎn),其中Off-target-1和4與設(shè)計(jì)的靶點(diǎn)僅有3個(gè)堿基差異。脫靶檢測(cè)分析發(fā)現(xiàn),EBE區(qū)被編輯的同時(shí),潛在脫靶位點(diǎn)的序列沒(méi)有發(fā)生突變,表明本研究設(shè)計(jì)的EBE區(qū)編輯靶點(diǎn)具有良好的特異性。本研究有助于進(jìn)一步獲得木薯MeSWEET10a基因啟動(dòng)子EBE區(qū)突變體,以增強(qiáng)木薯的抗細(xì)菌性枯萎病能力,為木薯抗病育種奠定材料和研究基礎(chǔ)。
參考文獻(xiàn)
[1] Yoodee S, Kobayashi Y, Songnuan W, et al. Phytohormone priming elevates the accumulation of defense-related gene transcripts and enhances bacterial blight disease resistance in cassava[J]. Plant Physiology and Biochemistry, 2018, 122: 65-77.
[2] Mora R E, Rodriguez M A, Gayosso L Y, et al. Using in vitro plants to study the cassava response to Xanthomonas phaseoli pv. manihotis infection[J]. Tropical Plant Pathology, 2019, 44(5): 423-429.
[3] 李超萍, 時(shí) ?濤, 劉先寶, 等. 國(guó)內(nèi)木薯病害普查及細(xì)菌性萎蔫病安全性評(píng)估[J]. 熱帶作物學(xué)報(bào), 2011, 32(1): 116-121.
[4] Cohn M, Morbitzer R, Lahaye T, et al. Comparison of gene activation by two TAL effectors from Xanthomonas axono-podis pv. manihotis reveals candidate host susceptibility genes in cassava[J]. Molecular Plant Pathology, 2016, 17(6): 875-889.
[5] Castiblanco L F, Gil J, Rojas A, et al. TALE1 from Xantho-monas axonopodis pv. manihotis acts as a transcriptional activator in plant cells and is important for pathogenicity in cassava plants[J]. Molecular Plant Pathology, 2013, 14(1): 84-95.
[6] Cohn M, Bart R S, Shybut M, et al. Xanthomonas axonopo-dis virulence is promoted by a transcription activator-like effector–mediated induction of a SWEET sugar transporter in cassava[J]. Molecular Plant-Microbe Interactions, 2014, 27(11): 1186-1198.
[7] Oliva R, Ji C H, Atienza-Grande G, et al. Broad-spectrum resistance to bacterial blight in rice using genome editing[J]. Nature Biotechnology, 2019, 37(11): 1344-1350.
[8] 李 ?嶄, 王亞杰, 陸小花, 等. 木薯MeSSⅢ基因的CRISPR/Cas9基因編輯載體構(gòu)建及驗(yàn)證[J]. 分子植物育種, 2020, 18(16): 5367-5372.
[9] Nekrasov V, Wang C M, Win J, et al. Rapid generation of a transgene-free powdery mildew resistant tomato by genome deletion[J]. Scientific Reports, 2017, 7(1): 1-6.
[10] Wang Y P, Cheng X, Shan Q W, et al. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew[J]. Nature Biotechnology, 2014, 32(9): 947-951.
[11] Zhang Y W, Bai Y, Wu G H, et al. Simultaneous modifica-tion of three homoeologs of TaEDR1 by genome editing en-hances powdery mildew resistance in wheat[J]. Plant Journal, 2017, 91(4): 714-724.
[12] Peng A, Chen S, Lei T, et al. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the suscep-tibility gene CsLOB1 promoter in Citrus[J]. Plant Biotech-nology Journal, 2017, 15: 1509-1519.
[13] Xu Z Y, Xu X M, Gong Q, et al. Engineering broad-spectrum bacterial blight resistance by simultaneously disrupting variable TALE-binding elements of multiple susceptibility genes in rice[J]. Molecular Plant, 2019, 12(11): 1434-1446.
[14] Chen L Q, Hou B H, Lalonde S, et al. Sugar transporters for intercellular exchange and nutrition of pathogens[J]. Nature, 2010, 468(7323): 527-532.
[15] Ferrari S, Galletti R, Denoux C, et al. Resistance to Botrytis cinerea induced in Arabidopsis by elicitors is independent of salicylic acid, ethylene, or jasmonate signaling but requires PHYTOALEXIN DEFICIENT3[J]. Plant Physiology, 2007, 144(1): 367-379.
[16] Chong J L, Piron M C, Meyer S, et al. The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea[J]. Journal of Expe-rimental Botany, 2014, 65(22): 6589-6601.
責(zé)任編輯:黃東杰
