朱紅毛斑蛾繁殖行為的研究
2021-12-27 11:21:52吳海盼劉俊延王小云鄭霞林
林業(yè)科學(xué)研究 2021年4期
吳海盼,劉俊延,王小云,陸 溫,鄭霞林
(廣西農(nóng)業(yè)環(huán)境與農(nóng)產(chǎn)品安全重點實驗室,廣西大學(xué)農(nóng)學(xué)院,廣西南寧 530004)
斑蛾總科(Zygaenoidea )包括斑蛾科(Zygaenida)、毛斑蛾科(Phaudidae)和絨蛾科(Megalopygidae)等13 科2600 余種,其中斑蛾科和毛斑蛾科中的許多種類是農(nóng)業(yè)、園藝和林業(yè)上的重要害蟲[1-3]。目前,針對斑蛾總科害蟲中有關(guān)繁殖方面的研究主要集中性沖突/選擇[4]、延遲交配[5]、性信息素[6-8]和滯育[9]等方面。尚未見有關(guān)毛斑蛾科昆蟲繁殖行為方面的研究報道。
朱紅毛斑蛾(Phauda flammans)(鱗翅目Lepidoptera:毛斑蛾科Phaudidae)是榕樹上的一種常暴發(fā)的食葉害蟲,其危害不僅影響榕樹的生長發(fā)育,還影響城市景觀和生態(tài)效應(yīng)[10]。該蟲廣泛分布于東南亞和南亞諸國[11-17],且已明確了其生物生態(tài)學(xué)特性,包括形態(tài)特征[10,18-19]、寄主選擇[20]、年生活史[10]、發(fā)育起點溫度和有效積溫[21]、抗寒力[22]、天敵種類[23],越冬蟲態(tài)的調(diào)查方法[24]和防治技術(shù)[8,25-27]等。目前,關(guān)于朱紅毛斑蛾的繁殖行為尚無相關(guān)研究報道。鑒于此,本研究研究了朱紅毛斑蛾的羽化、求偶、交配和產(chǎn)卵行為及節(jié)律,以期明確朱紅毛斑蛾的繁殖行為,為該蟲的種群管理提供基礎(chǔ)資料。
1 材料與方法
1.1 供試蟲源
在廣西大學(xué)校園內(nèi)(108°29′ E,22°85′ N)的小葉榕上采集6 齡朱紅毛斑蛾幼蟲,帶回室內(nèi)置于圓形塑料盒(底部直徑20.5 cm,頂部直徑25.0 cm,高15.0 cm;10 頭·盒?1)內(nèi),在溫度26 ± 2℃、相對濕度70% ± 10%、光周期為16 L:8 D 的環(huán)境條件下飼養(yǎng),每日更換新鮮小葉榕葉片直至化蛹,收集繭(蛹)待用。
1.2 研究方法
1.2.1 朱紅毛斑蛾羽化行為的研究 將繭(蛹)置于圓形塑料盒內(nèi)(底部直徑18.5 cm,頂部直徑23.0 cm;高10 cm;100 頭·盒?1),在上述飼養(yǎng)條件下,人工連續(xù)觀察和記錄塑料盒中朱紅毛斑蛾蛹的羽化情況,每隔1 小時統(tǒng)計羽化的雌蟲(n=493)和雄蟲(n= 536)數(shù)量并將其取出,直至無成蟲羽化為止,分析成蟲的羽化行為、日節(jié)律及晝夜節(jié)律。
1.2.2 朱紅毛斑蛾求偶行為的研究 將雌雄蛹分開保存在不同的塑料盒內(nèi),置于上述飼養(yǎng)條件下。待成蟲羽化后,隨機選取1 日齡健康雌雄蟲放入裝有盆栽小葉榕的網(wǎng)籠(長×寬×高= 100 cm×100 cm×180 cm)中(100 對·籠?1),人工觀察和統(tǒng)計每小時雄蟲求偶(雄蟲求偶時,腹部翹起幾乎與地面垂直,并快速振翅)的數(shù)量(n= 1299),并將統(tǒng)計過的雄蟲移出籠外,避免重復(fù)計數(shù)。
1.2.3 朱紅毛斑蛾交配行為的研究 隨機選取300 對1 日齡健康雌雄蟲放入網(wǎng)籠內(nèi),在上述飼養(yǎng)條件下人工觀察,發(fā)現(xiàn)交配時將交配的雌雄蟲輕輕取出并置于塑料盒中(1 對·盒?1),統(tǒng)計每小時內(nèi)的交配數(shù),并分析其交配行為和晝夜節(jié)律。對置于塑料盒內(nèi)交配中的朱紅毛斑蛾雌雄蟲采用數(shù)碼攝像機(DS-2CD3T45D-13,海康威視數(shù)字技術(shù)股份有限公司,杭州,浙江)連續(xù)拍攝,直至其結(jié)束交配,統(tǒng)計其交配時長。
1.2.4 朱紅毛斑蛾產(chǎn)卵行為的研究 隨機選取交配結(jié)束后的朱紅毛斑蛾雌蟲置于塑料盒中(1 頭·盒?1),并給以新鮮小葉榕枝條,在上述飼養(yǎng)條件下,使用數(shù)碼攝像機連續(xù)拍攝直至雌蟲死亡,記錄雌蟲每天每小時內(nèi)的產(chǎn)卵情況,分析其產(chǎn)卵行為和晝夜節(jié)律,并統(tǒng)計每頭雌蟲(n= 202)的總產(chǎn)卵量。每天收集榕樹葉上的卵塊置于墊有濕潤濾紙的培養(yǎng)皿內(nèi)(直徑= 12 cm),統(tǒng)計其在上述飼養(yǎng)條件下的孵化率。
1.3 數(shù)據(jù)分析
使用SPSS 16.0(SPSS Inc.,Chicago,IL,USA)軟件對24 h 內(nèi)不同時間段雌蟲的產(chǎn)卵率進行數(shù)據(jù)統(tǒng)計分析,采用ANOVA 單因素方差分析,并用Tukey’s HSD 進行多重比較(P<0.05)。統(tǒng)計分析前,百分率數(shù)據(jù)進行平方根轉(zhuǎn)換。
2 結(jié)果與分析
2.1 朱紅毛斑蛾的羽化行為
2.1.1 朱紅毛斑蛾的羽化行為特征 朱紅毛斑蛾羽化前,蛹體漸變?yōu)樯罴t色。近羽化時蛹體輕微扭動,隨后頻率逐漸加快,約3~5 min 后蛹的頭部先鉆出繭室一側(cè)留下的稀薄羽化孔,然后蟲體繼續(xù)抖動,頭胸部先鉆出蛹?xì)ぃㄟ^胸足的不斷爬行利用繭室羽化孔的摩擦逐步褪去腹部蛹?xì)ぃ細(xì)ち粲谟际覂?nèi)。成蟲爬出繭室后,在蛹室附近靜伏,并用前足梳理觸角,數(shù)分鐘后成蟲會通過其排泄孔射出棕黃色排泄物。待完全展翅后,雙翅豎立數(shù)分鐘,隨后飛行離開(圖1)。

圖1 朱紅毛斑蛾的羽化行為Fig.1 Emergence of Phauda flammans adult
2.1.2 朱紅毛斑蛾的羽化節(jié)律 朱紅毛斑蛾雌雄成蟲僅在光期06:00—16:00 羽化,在11:00—13:00達到羽化高峰期,且雌蟲羽化時間比雄蟲提早約1 h,雌雄成蟲的晝夜羽化節(jié)律均呈“單峰”型。采集的同一批樣本中,朱紅毛斑蛾化蛹持續(xù)15 d,蛹期11~17 d,雌雄成蟲的羽化持續(xù)19 d,雌蟲早于雄蟲約2 d 羽化,提早1 d 結(jié)束。雌雄蟲的日羽化高峰期均出現(xiàn)在開始羽化后的8~11 d,在羽化高峰期的羽化率分別占雌雄成蟲總羽化率的59.35%和57.01%,雌雄蟲的日羽化節(jié)律均呈“單峰”型(圖2)。
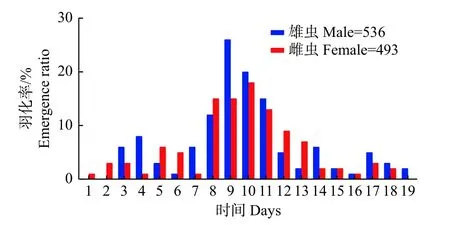
圖2 朱紅毛斑蛾成蟲的羽化節(jié)律Fig.2 Emergence of Phauda flammans adults
2.2 朱紅毛斑蛾的求偶行為
2.2.1 朱紅毛斑蛾的求偶行為特征 朱紅毛斑蛾雌雄蟲僅在光期求偶。雌蟲羽化后,大多數(shù)個體順著樹干爬至寄主植物枝干或葉片上,也有少量雌蟲通過飛行抵達寄主植物。雌蟲在寄主植物上尋找到合適的位置后,開始靜止等待雄蟲,在此過程中,常見雌蟲無規(guī)律快速震顫腹部,偶見腹部末端翹起現(xiàn)象。與此同時,雄蟲飛行搜索雌蟲。當(dāng)飛抵雌蟲附近時,雄蟲繼續(xù)爬行,并伴隨觸角快速擺動偵測雌蟲位置。待雄蟲準(zhǔn)確定位雌蟲后立即表現(xiàn)出明顯的求偶行為,包括雄蟲觸角不停敲擊雌蟲觸角和身體,雄蟲在雌蟲周圍快速振翅,腹部高高翹起幾乎垂直于地面,然后暴露陽莖試圖與雌蟲交配。雌蟲若接受雄蟲的求偶,則立即接受交配;雌蟲若不接受該雄蟲的求偶,則快速爬行以掙脫雄蟲,而雄蟲則窮追不舍,并重復(fù)上述求偶行為,試圖再次與雌蟲進行交配,若雌蟲接受該雄蟲的再次求偶則立即交配,否則雌蟲飛走,雄蟲需重新尋找和定位新的雌蟲(圖3)。

圖3 朱紅毛斑蛾的覓偶和求偶行為Fig.3 Mate searching,courtship and calling behaviors of Phauda flammans
2.2.2 朱紅毛斑蛾求偶節(jié)律 朱紅毛斑蛾從09:00—18:00 均有個體求偶,求偶高峰期為11:00—16:00,占求偶個體總數(shù)的69.44%(圖4)。
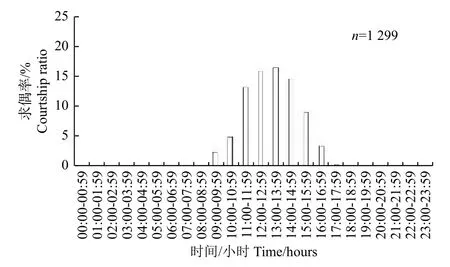
圖4 朱紅毛斑蛾求偶的晝夜節(jié)律Fig.4 Circadian rhythm of courtship of Phauda flammans
2.3 朱紅毛斑蛾的交配行為
2.3.1 朱紅毛斑蛾的交配節(jié)律及行為求偶率 朱紅毛斑蛾的交配行為在10:00—18:00 均可發(fā)生,14:00—16:00 為交配高峰期,暗期未見交配行為。朱紅毛斑蛾交配姿勢多為“一”或“V”字型(圖5),交配地點多在寄主植物葉背和枝干,交配時長為16.16 ± 0.26 h(n= 190),交配率63.33%,雌雄成蟲均一生只交配1 次,為單配制蛾類。

圖5 朱紅毛斑蛾交配行為Fig.5 Mating postures of Phauda flammans
2.4 朱紅毛斑蛾的產(chǎn)卵行為
2.4.1 朱紅毛斑蛾的產(chǎn)卵行為特征 朱紅毛斑蛾雌蟲在交配結(jié)束約1 h 或次日光期即可產(chǎn)卵。產(chǎn)卵前,雌蟲常在寄主植物上爬行,并用觸角不停地觸碰寄主植物以搜尋合適的產(chǎn)卵地。觀察發(fā)現(xiàn),卵多產(chǎn)于寄主植物的葉柄或葉尖部位。產(chǎn)卵時,腹部末端對準(zhǔn)產(chǎn)卵地,不斷抖動腹部,卵從腹部產(chǎn)卵孔排出,卵粒粘結(jié)呈線狀或鱗片狀,無重疊(圖6)。每頭雌蟲一生產(chǎn)1~2個卵塊,單雌平均產(chǎn)卵量為107.3 ± 3.0 粒,平均孵化率為88.4 %±1.6%。

圖6 朱紅毛斑蛾的產(chǎn)卵行為Fig.6 Oviposition behaviors of Phauda flammans
2.4.2 朱紅毛斑蛾的產(chǎn)卵節(jié)律 在5:00—23:00 均可觀察到朱紅毛斑蛾雌蟲產(chǎn)卵,但主要集中在10:00—14:00(圖7),且其產(chǎn)卵率顯著高于其他時間段(F= 32.73,df= 24,725,P<0.001)。
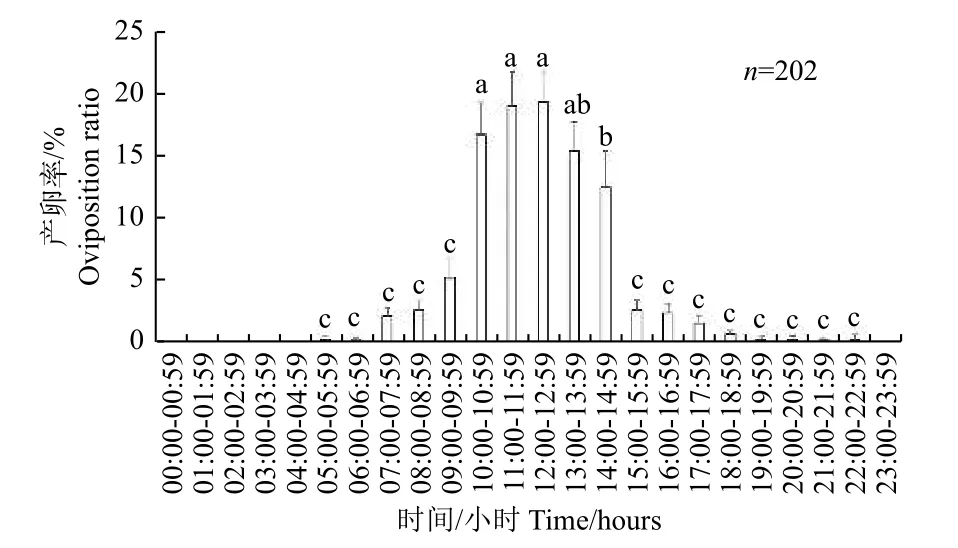
圖7 朱紅毛斑蛾產(chǎn)卵的晝夜節(jié)律Fig.7 Circadian rhythm of oviposition of Phauda flammans
3 討論
一般情況下,昆蟲經(jīng)過長期的進化,其繁殖行為及節(jié)律相對穩(wěn)定,主要是受其生物鐘的調(diào)節(jié)[28]。本研究結(jié)果表明,朱紅毛斑蛾的羽化、求偶和交配行為均發(fā)生在光期,為典型的日行性蛾類,與已報道的斑蛾科其他昆蟲的繁殖行為和節(jié)律相似,如云南錦斑蛾(Achelurayunnanensis)[29]、朱頸褐錦斑蛾(Soritialeptalina)[30]、大葉黃楊長毛斑蛾(Pryeriasinica)[31],說明毛斑蛾科昆蟲經(jīng)過長期進化也表現(xiàn)出與斑蛾科類似的在光期繁殖的生物學(xué)特性。前人研究結(jié)果表明,昆蟲繁殖行為及節(jié)律的變化可以減少物種對同一資源的直接競爭,并通過種群行為的同步,確保相似種間的遺傳隔離[32]。事實上,蛾類的繁殖行為及節(jié)律除了由內(nèi)在的生理因素決定之外,還受環(huán)境溫度、光周期、濕度等生態(tài)因子的影響。然而,控制朱紅毛斑蛾繁殖行為及節(jié)律的內(nèi)外因子目前均尚不明確,需進一步研究。
朱紅毛斑蛾羽化后,雌蟲多停留在隱蔽處等待雄蟲覓偶,覓偶成功后均是雄蟲主動發(fā)起求偶,這與多數(shù)蛾類昆蟲的求偶行為相似,如柚木野螟(Eutectona machaeralis)、櫟黃枯葉蛾(Trabala vishnou gigantina)、草原毛蟲(Gynaephora alpherakii)等[33-35]。前期研究證實,朱紅毛斑蛾雌蟲釋放的性信息素對雄蟲具有顯著的引誘作用[8],推測朱紅毛斑蛾雄蟲在覓偶過程中依靠雌蟲釋放性信息素進行異性定位,但雄蟲對性信息素的感受機制需進一步研究。
在蛾類昆蟲中,多數(shù)種類均有多次交配的現(xiàn)象,因為多次交配對其有物質(zhì)利益和基因利益等[36]。然而,朱紅毛斑蛾雌雄成蟲一生僅交配1次,為單配制蛾類,說明朱紅毛斑蛾進行一次交配就能提供足夠的精子供雌蟲使用。在部分蛾類昆蟲中,也觀察到這種現(xiàn)象,如馬尾松毛蟲(Dendrolimus punctatus)、小菜蛾(Plutella xylostella)、粉斑螟(Cadra cautella)和棉褐環(huán)野螟(Haritalodes derogata)[37-40]。朱紅毛斑蛾雖然一生僅交配1 次,但雌蛾的累計產(chǎn)卵量和孵化率均較高,說明該蟲繁殖系數(shù)高,群體積累快,可能是其常暴發(fā)的潛在原因之一。
“選擇性和適合度”假說(preference-performance hypothesis,PPH)認(rèn)為由于植食性昆蟲幼蟲期不能遠距離移動,所以雌蟲常將卵產(chǎn)于適合其子代發(fā)育的寄主上[41]。朱紅毛斑蛾是寡食性害蟲,雌蟲將卵產(chǎn)于小葉榕和垂葉榕葉柄或葉尖,推測是朱紅毛斑蛾雌蟲為便于其子代孵化后能有適口和充足的食物有關(guān)[42]。研究表明,卵的成功發(fā)育需要一個合適的產(chǎn)卵場所,其非生物條件適合,捕食率、寄生率和疾病率較低,及給子代留有充足的食物[43]。朱紅毛斑蛾雌蟲將卵產(chǎn)于寄主植物葉柄或葉尖,除了為子代提供充足的食物之外,是否還與抵御天敵的捕食或寄生有關(guān)尚不明確。此外,植食性昆蟲產(chǎn)卵場所的選擇是昆蟲與寄主植物長期協(xié)同進化的結(jié)果[44],但朱紅毛斑蛾產(chǎn)卵場所選擇的進化驅(qū)動力仍有待進一步研究。
本研究結(jié)果還提示,可基于朱紅毛斑蛾的繁殖行為及節(jié)律,在野外利用性信息素干擾其交配,使其錯過最適交配時間,降低交配率,從而達到控制其種群數(shù)量的目的[5];在其繁殖高峰期,應(yīng)用性信息素大量誘捕和監(jiān)測該蟲野外的種群動態(tài)及密度,為其防控、預(yù)測預(yù)報和制定綜合防治策略等提供參考依據(jù)。
4 結(jié)論
朱紅毛斑蛾僅在光期求偶、交配和產(chǎn)卵,并形成了相對固定的行為和節(jié)律,是一種日行性蛾類;一生僅交配1 次,為單配制蛾類。
