β3 腎上腺素能受體活化抑制慢性間歇低氧誘導的動脈粥樣硬化進展的機制
2022-01-06 05:40:12索旻王越劉夢如劉倍倍陳蕾蕾王悅王志強張慧娜吳小凡
中國循環雜志 2021年12期
索旻,王越,劉夢如,劉倍倍,陳蕾蕾,王悅,王志強,張慧娜,吳小凡
阻塞性睡眠呼吸暫停綜合征(OSA)被列為動脈粥樣硬化性疾病的新危險因素[1-2],慢性間歇低氧(CIH)已被證實是OSA 的主要病理生理過程,并且可通過上調氧化應激反應促進動脈粥樣硬化的進展[3]。CIH 下交感神經系統興奮性顯著升高,有研究表明持續興奮的交感神經系統會使β3 腎上腺素能受體(β3 AR)出現脫敏現象,導致其活性下降[4];常氧下活化β3 AR 可抑制巨噬細胞內煙酰胺腺嘌呤二核苷酸磷酸(NADPH)氧化酶(NOX)活性,進而減少活性氧產生[5],然而CIH 下β3 AR 活化對動脈粥樣硬化的影響及具體機制尚不明確。p22phox作為NOX 的重要亞基[6],已被證明能夠促進動脈粥樣硬化進展[7],因此,CIH/β3 AR/氧化應激通路很可能是CIH 促進動脈粥樣硬化進展的重要機制之一。本研究首先在載脂蛋白E 基因敲除(ApoE-/-)小鼠中觀察活化β3 AR 對氧化應激反應和動脈粥樣硬化進展的影響,進一步聚焦氧化應激反應發生的主要場所——巨噬細胞,觀察CIH 下活化β3 AR 抑制氧化應激反應的可能機制。
1 材料與方法
主要試劑及材料:β3 AR 特異性激動劑米拉貝隆購自美國MedChemExpress 公司;蘇木素-伊紅(HE)染液、青霉素-鏈霉素混合溶液和胰蛋白酶購自北京索萊寶科技有限公司;CD68、p22phox 和β3 AR 抗體均購自美國Abcam 公司;超氧化物陰離子熒光探針購自上海碧云天生物技術有限公司;甘油醛-3-磷酸脫氫酶 (GAPDH)抗體購自北京普利萊基因技術有限公司;辣根過氧化物酶標記的羊抗兔二抗和二氨基聯苯胺(DAB)顯色劑均購于北京中杉金橋公司;羊抗兔熒光二抗和羊抗小鼠熒光二抗均購自美國Li-Cor 公司;總抗氧化能力檢測試劑盒購自北京普利萊基因技術有限公司;胎牛血清和RPMI 1640 培養基均購自美國Gibco 公司。
實驗動物及細胞:6~7 周齡,體質量(25±3)g,無特定病原體級,雄性ApoE-/-小鼠15 只,購自北京華阜康生物科技股份有限公司,小鼠質量合格編號11401500043662,在首都醫科大學附屬北京安貞醫院實驗動物房內飼養。以下動物實驗已通過本院實驗動物倫理委員會批準。小鼠單核巨噬細胞白血病細胞RAW264.7 源自Abelson 鼠科白血病病毒誘導的腫瘤,購自國家實驗細胞資源共享平臺(北京總部)。
實驗分組及干預:ApoE-/-小鼠飼養:按照隨機數表法將小鼠平均分為3 組:(1)常氧組給予高脂飼料(含21%脂肪、0.15%膽固醇)喂養12 周。(2)低氧組給予高脂飼料喂養的同時,于實驗第4 周進入低氧動物箱,箱內循環充入氮氣和壓縮空氣,主控板控制氮氣和壓縮空氣的轉換,使艙內氧濃度在5%~21%之間循環,每個循環180 s[8]。(3)低氧+米拉貝隆組小鼠給予高脂飼料喂養的同時,于實驗第4 周進入低氧動物箱,同時每日以10 mg/kg 的米拉貝隆灌胃。常氧組和低氧組小鼠于實驗第4 周每日接受相同體積的米拉貝隆溶劑(即含10%二甲基亞砜的生理鹽水)灌胃,低氧組和低氧+米拉貝隆組小鼠每天在低氧動物箱內8 h,箱內溫度、濕度與室內相同。
RAW264.7 細胞培養:RAW264.7 細胞置于二氧化碳培養箱中,在37℃、5%二氧化碳條件下,以含10%胎牛血清的RPMI 1640 培養基培養,取對數生長期細胞應用于實驗。實驗分為常氧組、低氧組和低氧+米拉貝隆組,將細胞接種于6 孔板中,每組6 孔,調整細胞濃度為1×106個/孔。低氧+米拉貝隆組細胞給予0.5 μmol/L 米拉貝隆處理。低氧組和低氧+米拉貝隆組細胞置于37℃低氧培養箱內,二氧化碳濃度5%,氧濃度在5%~21%之間循環,每個循環1 h[9]。48 h 后收取細胞。
小鼠標本采集與處理:小鼠夜間禁食水12 h 后以5%的戊巴比妥鈉腹腔注射進行麻醉,自心尖抽取血液標本,置于-80℃冰箱中保存備用。留取小鼠心臟基底部,4%多聚甲醛固定,于4℃冰箱中保存備用[10];留取頸動脈,用包埋劑包埋后于-20℃冰箱中保存備用。
主動脈根部HE 染色:在主動脈3 個瓣葉水平以5 μm 厚度對組織蠟塊進行連續切片,對主動脈根部組織進行HE 染色,顯微鏡拍照后利用NISELEMENT 軟件測定粥樣硬化斑塊表面積與動脈壁表面積,計算斑塊面積占動脈壁表面積的百分比[7]。
免疫組織化學染色:小鼠主動脈根部石蠟切片脫蠟后于檸檬酸鈉修復液中進行抗原修復,以3%H2O2水溶液滅活內源性過氧化物酶,用5%牛血清白蛋白封閉后分別滴加CD68、p22phox 抗體,4℃孵育過夜,次日滴加二抗室溫孵育1 h 后,滴加DAB 顯色液進行顯色。顯微鏡拍照,計算棕褐色陽性面積占組織面積的百分比[10]。
超氧化物陰離子熒光探針檢測活性氧:對頸動脈組織進行冰凍切片,厚度約7 μm,將熒光探針(10 mmol/L)滴于組織上,37℃孵育30 min,50%甘油封片后立即在顯微鏡下觀察、拍照,活性氧在激發光下呈紅色熒光。計算紅色熒光面積占血管組織總面積的百分比。
ABTS 法檢測總抗氧化能力:根據試劑盒說明,倍比稀釋標準品溶液用于標準曲線測定;于96 孔板中依次加入檢測工作液、樣品或標準品后,混勻,室溫孵育5 min 后利用酶標儀測定734 nm 波長處吸光度。根據標準曲線,計算樣品總抗氧化能力,細胞樣品根據蛋白濃度計算每毫克蛋白中總抗氧化能力。
RNA 提取與轉錄組測序:取ApoE-/-小鼠整條主動脈,分離周圍脂肪組織及筋膜,利用TRIzol 法提取總RNA,以BGISEQ500 平臺進行轉錄組測序(RNA-seq)。每個基因的表達水平通過RSEM 標準化為FPKM。利用差異分析軟件DEseq2 篩選三組間氧化應激相關的差異表達基因[11]。對差異表達基因進行基因本體論(GO)和京都基因與基因組百科全書(KEGG)富集分析,以P<0.05 為篩選條件。
蛋白免疫印跡(Western blot)檢測:收取細胞進行蛋白定量后,進行蛋白變性[12],將蛋白樣本等量上樣至十二烷基硫酸鈉聚丙烯酰胺凝膠進行電泳后,以200 mA電流冰上轉膜至聚偏二氟乙烯(PVDF)膜上,10%脫脂奶粉封閉1 h 后,于4℃分別孵育β3 AR、p22phox 和GAPDH 一抗過夜,次日以熒光標記二抗室溫避光孵育1 h,用Odyssey 雙色紅外激光成像系統掃描PVDF 膜檢測灰度值。
統計學方法:采用SPSS 25.0 軟件進行統計分析。正態分布的計量資料以均數±標準差表示,多組間比較采用單因素方差分析,采用LSD 法進行多重比較。以P<0.05 表示差異有統計學意義。
2 結果
2.1 各組小鼠動脈粥樣硬化斑塊情況及β3 AR、p22phox 表達
低氧組較常氧組和低氧+米拉貝隆組小鼠:主動脈根部HE 染色動脈粥樣硬化斑塊面積占動脈壁表面積的百分比更高[(27.88±5.83)%vs. (19.84±5.13)%; (27.88±5.83)% vs.(16.33±3.60)%,P均<0.05,n=5](圖1A);斑塊內CD68 陽性面積百分比更高[(10.63±3.18)% vs.(5.31±2.77)%;(10.63±3.18)% vs. (5.20±1.40)%,P均<0.05,n=5)(圖1B);斑塊內β3 AR 陽性面積百分比更高[(5.06±0.44)% vs. (2.99±1.56)%;(5.06±0.44)% vs. (2.61±0.95)%,P均<0.05,n=5](圖1C);p22phox 陽性面積百分比更高[(4.82±0.85)%vs. (1.29±0.27)%;(4.82±0.85)% vs. (0.94±0.63)%,P均<0.05,n=5](圖1D)。各組小鼠斑塊內p22phox陽性面積百分比與β3 AR 的陽性面積百分比具有正性相關(r=0.90,P<0.05,圖1E)。
2.2 各組小鼠氧化應激水平及血清總抗氧化能力
低氧組較常氧組和低氧+ 米拉貝隆組小鼠,頸動脈組織活性氧陽性面積百分比更高[(29.25±4.87)% vs. (12.58±1.73)%,(29.25±4.87)% vs. (6.00±2.08)%;P均<0.05],見圖2A。低氧組較常氧組和低氧+米拉貝隆組小鼠,血清總抗氧化力水平降低[(2.09±0.11) mmol/L vs. (2.74±0.33) mmol/L; (2.09±0.11) mmol/L vs.(2.73±0.15) mmol/L,P均<0.05],見圖2B。
進一步通過RNA-seq 對3 組小鼠的基因表達譜進行檢測,發現與常氧組相比,低氧組小鼠存在796 種差異表達基因,其中437 種基因表達上調,359 種基因表達下調;與低氧組相比,低氧+米拉貝隆組小鼠存在474 種差異表達基因,其中130 種基因表達上調,344 種基因表達下調。上述兩兩比較后的差異表達基因有183 種重復,對這183 種差異表達基因進行分子功能(圖2C)和生物過程(圖2D)的GO 富集分析,篩選富集基因數前10 名的GO 號,發現3 組小鼠差異表達基因在氧化還原過程中具有明顯富集。
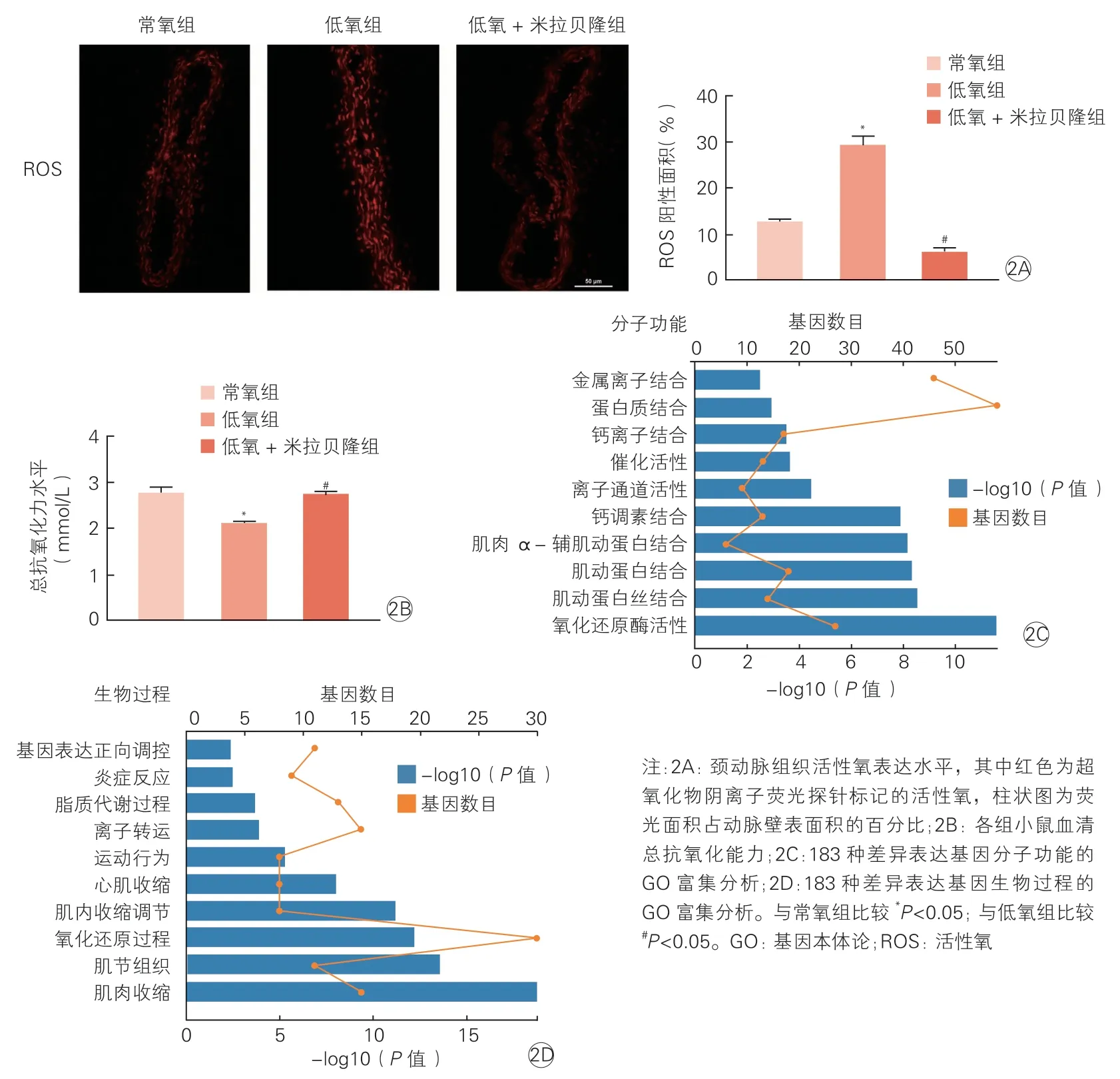
圖2 小鼠動脈組織氧化應激水平及血清總抗氧化能力(n=5)
故進一步在183 個差異基因中篩選與氧化應激相關的基因,共發現36 個基因,其在各組小鼠中的表達水平如圖3A 所示,在這些基因中,Hephl1、Marc1、Moxd1、Alox15、Hpdl、Th、Dbh、Sqle、Adh7、Nsdhl、Fasn、Gpd1、Sc5d、Sdr39u1、Hsd11b1、Dio2、Gmpr、Pdha1、Pdhb、Uqcrfs1、Etfdh、Sdhb、Idh3a、Hsd17b12、Gpd2、Dlat、Me1等基因主要起促進氧化的作用,與常氧組相比,其在低氧組的表達水平上調,給予米拉貝隆后,其表達水平降低,其中Fasn曾被報道可隨p22phox 水平降低而降低[11]。Cyp2b10主要起抑制氧化應激的作用,與常氧組相比,其在低氧組的表達水平降低,給予米拉貝隆后,其表達水平上調。對這36 個差異基因進行KEGG 信號通路富集分析,共發現17 種有統計學意義的信號通路(圖3B)。
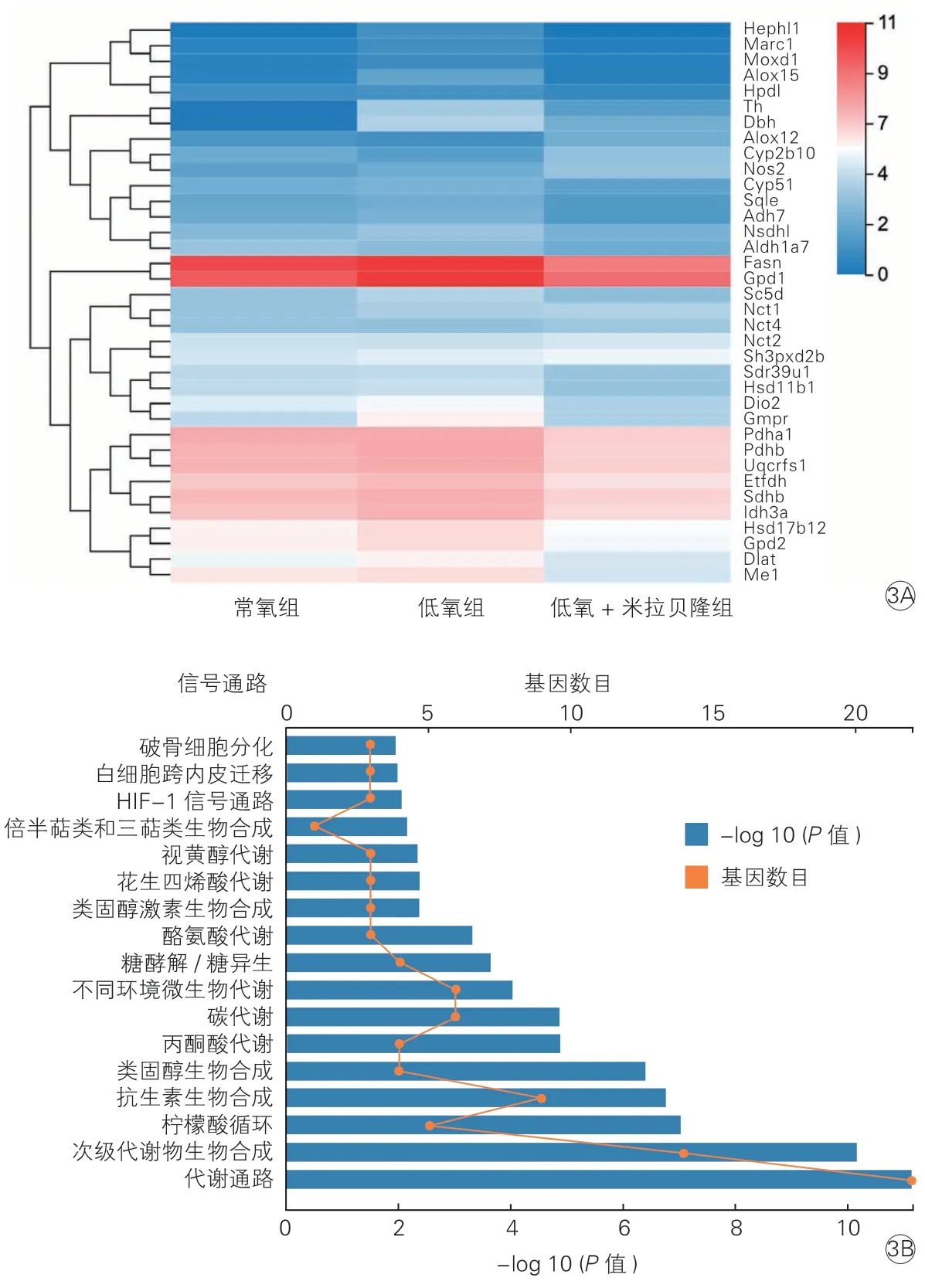
圖3 各組小鼠主動脈組織36 個氧化應激相關基因的表達差異(3A)與KEGG 信號通路富集分析(3B)
2.3 RAW264.7 巨噬細胞β3 AR 及氧化應激水平
Western blot 檢測結果顯示,常氧組、低氧組和低氧+米拉貝隆組β3 AR 灰度值分別為0.40±0.03、0.51±0.04 和0.90±0.01,低氧組β3 AR 表達水平高于常氧組,但低于低氧+米拉貝隆,差異均有統計學意義(P均<0.05),見圖4A、4B。常氧組、低氧組和低氧+米拉貝隆組p22phox 灰度值分別為0.70±0.05、0.86±0.07 和0.72±0.07,低氧組p22phox 表達水平高于常氧組和低氧+米拉貝隆組,差異均有統計學意義(P均<0.05),見圖4A、4C。ABTS 法檢測發現,低氧組總抗氧化力水平低于常氧組和低氧+米拉貝隆組細 胞[(116.39±88.33) μmol/mg vs.(287.11±65.43)μmol/mg;(116.39±88.33) μmol/mg vs.(470.86±197.05)μmol/mg,P均<0.05],見圖4D。
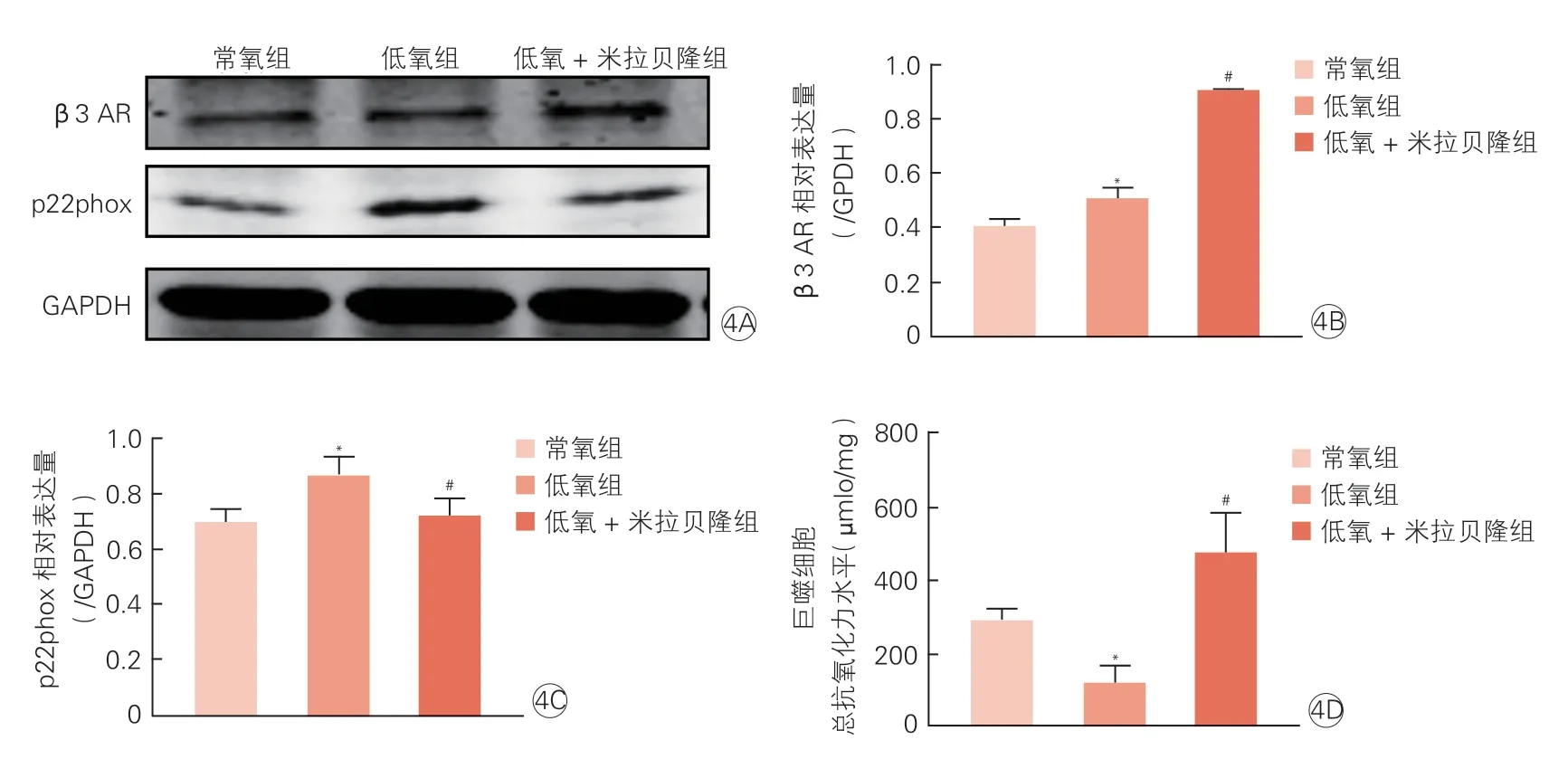
圖4 RAW264.7 巨噬細胞β3 AR 及氧化應激水平(n=3)
3 討論
既往有研究發現β3 AR 活化可減緩常氧環境下動脈粥樣硬化的進展[4-6],本研究在CIH 環境下驗證了β3 AR 活化對動脈粥樣硬化的保護作用,特異性激動β3 AR 可減緩CIH 誘導的ApoE-/-小鼠動脈粥樣硬化發生,其發生機制與抑制巨噬細胞氧化應激反應、增強巨噬細胞抗氧化能力密切相關。
CIH 促進動脈粥樣硬化進展的過程可能涉及炎癥反應、血管新生、氧化應激反應等多條通路。既往研究在C57BL/6J 小鼠[13]和ApoE-/-小鼠[14]中均發現CIH 可促進動脈粥樣硬化進展,本研究選用ApoE-/-小鼠,通過8 周CIH 處理,快速誘導動脈粥樣硬化模型,同樣觀察到CIH 對ApoE-/-小鼠動脈粥樣硬化進展的促進作用。同時,通過檢測粥樣硬化斑塊內細胞成分,發現CIH 可顯著增加斑塊內巨噬細胞數量。巨噬細胞在斑塊進展和斑塊破裂中均發揮重要作用[15],已有研究報道,CIH 可降低巨噬細胞吞噬脂質/膽固醇的功能,增加炎癥因子的分泌[16]。在斑塊進展過程中,氧化修飾的低密度脂蛋白(oxLDL)同樣可降低巨噬細胞的吞噬功能,進而促進斑塊內壞死細胞形成壞死核心[17]。oxLDL 影響巨噬細胞吞噬功能的機制之一是增強氧化應激反應、產生過量的活性氧[18],NOX 是合成活性氧的關鍵酶,p22phox 是其重要的亞基[19],本研究發現,在CIH 促進動脈粥樣硬化進展的過程中,小鼠頸動脈組織活性氧升高,主動脈根部p22phox 表達水平顯著升高,同時小鼠血清總抗氧化力下降,提示在CIH 促進ApoE-/-小鼠動脈粥樣硬化進展的過程中,氧化應激反應加劇在其中發揮重要作用,抑制氧化應激反應有望減緩CIH 對動脈粥樣硬化的促進作用。
β3 AR 已被證實廣泛分布于棕色脂肪組織和血管等組織中[4],在既往研究中,β3 AR 通過調節棕色脂肪組織而發揮的抗動脈粥樣硬化作用受到廣泛關注,其在其他部位的作用機制尚不明確;另外,既往研究均是在常氧環境下形成動脈粥樣硬化斑塊,CIH 環境下,β3 AR 在動脈粥樣硬化進展中發揮何種作用尚未可知。CIH 環境下交感神經系統興奮性顯著升高,有研究表明長期興奮的交感神經系統會使β3 AR出現脫敏現象,即β3 AR活性下降[4];本研究在在體實驗和體外實驗中均發現,低氧組β3 AR 表達水平較常氧組明顯上調,這可能是對脫敏現象的代償性調節。與Nagai 等[20]報道的在CIH環境下大鼠肺組織中巨噬細胞β3 AR 表達水平上升的結果一致。給予米拉貝隆刺激后,小鼠粥樣硬化斑塊內β3 AR 表達水平顯著下降,而RAW264.7巨噬細胞中β3 AR 表達水平呈增高,這種不一致的現象可能與在體實驗和體外實驗中米拉貝隆的藥物濃度和刺激時間不同有關。在體實驗中活化β3 AR后顯著減緩CIH 誘導的動脈粥樣硬化,同時減少斑塊內巨噬細胞數量。進一步在巨噬細胞水平探討β3 AR 活化改善CIH 誘導的動脈粥樣硬化的可能機制,發現米拉貝隆激活巨噬細胞的β3 AR 后,可顯著降低巨噬細胞p22phox 表達水平,同時增強細胞的總抗氧化力。Hadi 等[5]在子宮肌層的巨噬細胞中發現β3 AR 活化可抑制氧化酶2(NOX2)活性進而減少活性氧的產生。本研究在既往研究的基礎上,結合轉錄組測序分析,進一步明確:與常氧環境相比,CIH 可上調促進氧化應激的相關基因表達,活化β3 AR 后可使上述基因表達下調。同時發現β3 AR 活性與NOX 的亞基p22phox 的表達之間具有重要聯系,提示β3 AR 活化后抑制巨噬細胞氧化應激反應、增強總抗氧化力可能是其延緩CIH 誘導的動脈粥樣硬化進展的機制,但β3 AR 與上述氧化應激相關基因之間的具體調節機制還有待于進一步探究。
綜上所述,本研究在ApoE-/-小鼠中發現β3 AR活化可減緩CIH 誘導的動脈粥樣硬化,其具體機制可能是降低巨噬細胞p22phox 表達、抑制氧化應激反應。β3 AR 有望成為OSA 患者防治動脈粥樣硬化疾病的有效靶點。
利益沖突:所有作者均聲明不存在利益沖突
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
學苑創造·A版(2020年9期)2020-10-13 09:41:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:56
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
西南軍醫(2016年6期)2016-01-23 02:21:19
新疆醫科大學學報(2015年10期)2015-12-26 12:33:30
癌變·畸變·突變(2015年3期)2015-02-27 06:15:09
西南軍醫(2015年2期)2015-01-22 09:09:37
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
食品科學(2013年15期)2013-03-11 18:25:48
