雙歧桿菌和乳桿菌對水蘇糖的利用特性
2022-01-13 08:52:48鄔佳穎陳敏暄金天賜王順合崔樹茂唐鑫毛丙永
食品與發酵工業 2021年24期
鄔佳穎,陳敏暄,金天賜,王順合,崔樹茂,唐鑫,毛丙永
(江南大學 食品學院,江蘇 無錫,214122)
水蘇糖是一種由半乳糖(α1→6)半乳糖(α1→6)葡萄糖(α1→2β)果糖組成的功能性低聚糖,具有促進腸道有益菌的增殖、潤腸通便、保護肝臟等功能[1]。但由于人的小腸不能合成分解水蘇糖所必需的α-半乳糖苷酶,因此水蘇糖無法被小腸消化吸收,會直接進入大腸,從而被腸道細菌發酵[2]。
人體攝入過量水蘇糖可能會導致脹氣等不良反應,而這些負面影響是劑量依賴性的,中等劑量的水蘇糖在腸內發酵有利于益生菌如雙歧桿菌的代謝[3]。腸道內的益生菌通過利用水蘇糖增殖,從而抑制腐敗菌生長,起到調節腸道菌群、維持腸道微生態穩定等功能。
大量研究表明雙歧桿菌和乳桿菌都可以利用水蘇糖。早在1978年,YAZAWA等[4]曾報道,除兩歧雙歧桿菌外,所有人類來源的雙歧桿菌都能利用水蘇糖。HAYAKAWA等[5]研究大豆低聚糖對人類糞便菌群的影響時發現,雙歧桿菌屬、乳桿菌屬等7個屬的58株菌能夠利用水蘇糖。PAN等[6]發現水蘇糖通過誘導植物乳桿菌產生更多的α-半乳糖苷酶水解水蘇糖,增加其糖代謝和細胞活性來高度促進自身增殖。但是目前對于雙歧桿菌和乳桿菌利用水蘇糖的相關研究大多集中在利用情況上,利用方式的研究相對較少。
本研究從人類糞便來源的32株雙歧桿菌和乳桿菌中篩選出能夠利用水蘇糖的菌株,通過測定培養液上清液中糖的組成、α-半乳糖苷酶活力并結合基因組草圖分析等方法,研究雙歧桿菌、乳桿菌對水蘇糖的利用特性,這對于水蘇糖促進人體健康的研究具有一定的指導意義。
1 材料與方法
1.1 實驗材料
1.1.1 實驗試劑
水蘇糖,上海源葉生物科技有限公司;棉籽糖,生工生物工程(上海)股份有限公司;酵母提取物、胰蛋白胨,英國Oxoid公司。
1.1.2 實驗菌株
實驗所用32株雙歧桿菌和乳桿菌,由本實驗室菌種保藏中心(Culture Collection of Food Microorganisms,CCFM)提供,具體信息如表1所示。
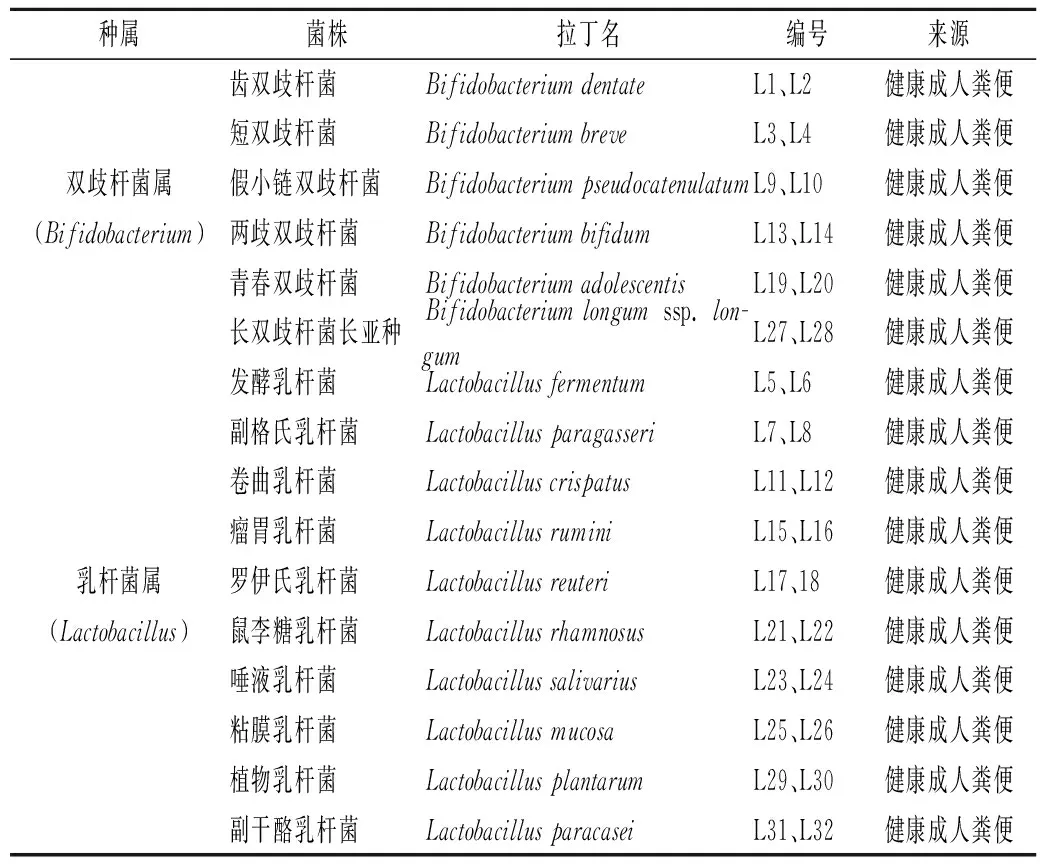
表1 實驗菌株信息Table 1 The information of the experimental strains
1.2 實驗方法
1.2.1 利用水蘇糖的雙歧桿菌和乳桿菌的確定
將菌株接種于以水蘇糖作唯一碳源、并添加溴甲酚紫指示劑的腸道微生物培養基(GMM)平板上,置于厭氧工作站37 ℃培養24~48 h;挑取使培養基變黃的菌株,劃線純化2次后,轉接到水蘇糖GMM液體培養基中,置于厭氧工作站37 ℃培養24~48 h,通過HPLC分析上清液中水蘇糖含量(方法見1.2.2),以未接種菌株的GMM液體培養基為空白對照,根據水蘇糖消耗量確定能夠利用水蘇糖的菌株。
1.2.2 細菌培養上清液中水蘇糖的HPLC分析
菌株在以水蘇糖作唯一碳源的GMM培養基中37 ℃厭氧培養24 h,菌液以8 000 r/min離心5 min收集上清液,采用HPLC測定上清液中水蘇糖的含量。色譜條件:采用SugarPak1糖柱(300 mm,id 3.5 μm),柱溫85 ℃,流動相為純水,流量為0.4 mL/min,使用外標法確定水蘇糖的含量[7]。
1.2.3 細菌利用水蘇糖生長的表征
將菌株活化后,接種于以水蘇糖作唯一碳源的GMM培養基中37 ℃厭氧培養。自初始接種時起,每隔1 h取1 mL待測菌液于1.5 mL 離心管以8 000 r/min離心5 min,收集上清液和菌泥備用。同時,測定菌液的pH和600 nm處的吸光度(OD600)。待菌株生長進入穩定期后,停止實驗。
1.2.4 α-半乳糖苷酶活力測定
α-半乳糖苷酶活力測定參照SCALABRINI等[8]和鄔佳穎等[9]的方法:上述生長過程中得到的上清液和菌泥,其中菌泥用檸檬酸-檸檬酸鈉緩沖液洗滌2次后重懸,加入100 μL氯仿,振蕩30 s,制成粗酶液。將250 μL粗酶液或上清液與對硝基苯α-D-氨基半乳糖苷(p-nitrophenyl β-D-glucopyranoside,PNPG)混合,37 ℃反應30 min,加入500 μL 0.2 mol/L碳酸鈉溶液終止反應,測定420 nm下的吸光值。菌泥測定結果對應胞內酶活力,上清液測定結果對應胞外酶活力。
酶活力定義:在測定條件下,1 min釋放1 μmol的對硝基苯酚所消耗的酶量(mL)為1個酶活力單位。
1.2.5 基因組草圖的測定
將菌株以4%的接種量接種至5 mL MRS液體培養基中進行活化,置于厭氧工作站37 ℃恒溫培養24~48 h。活化2次的菌株以4%的接種量接種至裝有100 mL MRS液體培養基的藍蓋瓶中,置于厭氧工作站培養24~48 h。菌液經8 000 r/min離心10 min得到菌泥,加入30%的甘油保存菌株,送至上海美吉生物醫藥科技有限公司完成基因組草圖測序。
利用Illumina HiSeq 2000(Illumina Inc USA)測序平臺對送測菌株進行雙末端測序,獲得原始數據,基因組覆蓋度達到100倍[10]。使用短序列組裝軟件SOAPdenovo2進行從頭組裝,獲得菌株基因組序列[11]。運用GeneMark1.1和Glimmer 3.02軟件對組裝結果進行基因denovo預測,獲取開放閱讀框(Open Reading Frame,,ORF)等相關信息[12-13]。在預測得到的編碼基因基礎上進行基因的功能注釋,將草圖基因的蛋白質序列與KEGG(Kyoto Encychopedia of Genes and Genomes)、蛋白相鄰類的聚簇(Cluster of Orthologous Groups of proteins,COG)等數據庫進行BLASTP對比,保留前5條相似性較高序列的結果,獲得相應的功能注釋信息,以此初步確定該蛋白序列與數據庫中的蛋白質具備相似的功能。上述步驟均由上海美吉生物醫藥科技有限公司完成。
1.2.6 α-半乳糖苷酶的亞細胞定位分析
利用LOCTREE對菌株中的α-半乳糖苷酶進行亞細胞定位分析。在LOCTREE中輸入該酶的氨基酸序列,選擇物種細菌,系統將對該酶位置進行預測。
1.2.7 數據分析
實驗中pH、 OD600、 糖含量以及酶活力的測定均為3次平行,結果以平均值±標準差表示。作圖采用Origin 9.0,對數據進行分析與處理。
2 結果與分析
2.1 能夠利用水蘇糖的菌株確定
腸道細菌利用水蘇糖生長后會產酸,使指示劑變色,即溴甲酚紫變黃。根據指示劑變色結果,初步篩選出能利用水蘇糖的雙歧桿菌和乳桿菌24株(表2)。
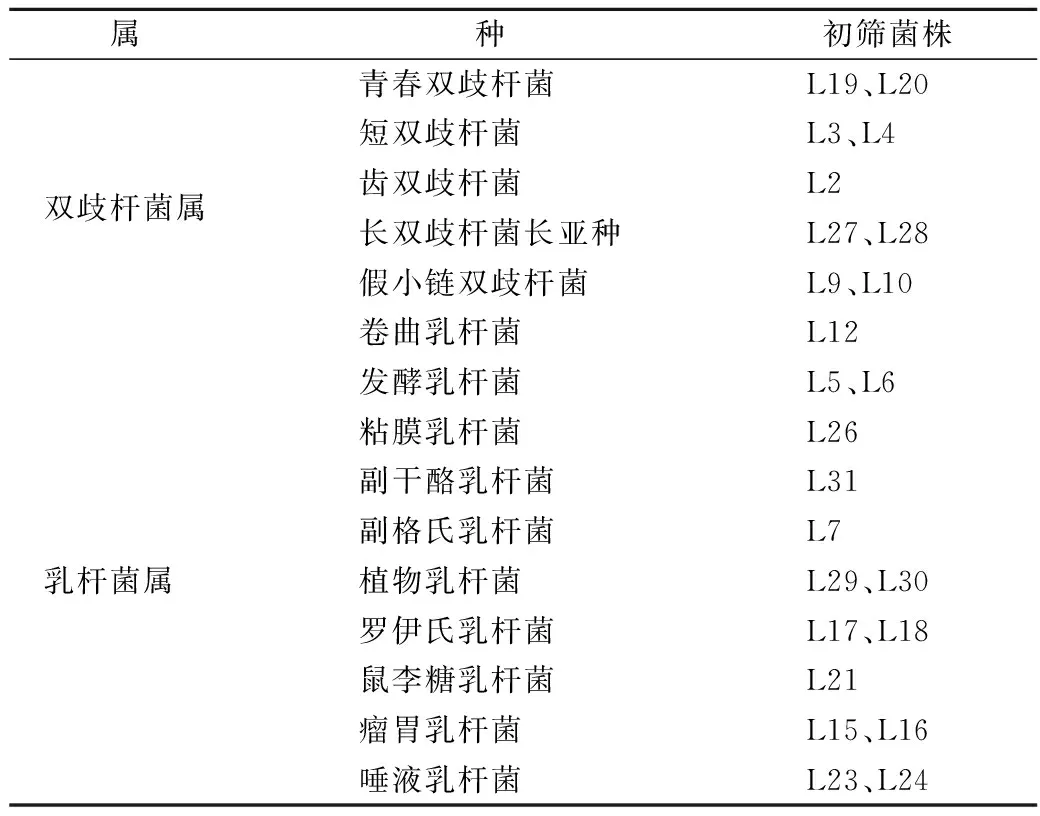
表2 能利用水蘇糖細菌的初步篩選Table 2 The primary screening of bacteria capable of using stachyose
HPLC法測定上述菌株上清液中水蘇糖含量,與初始液體培養基中的水蘇糖含量(8 g/L)進行比較,確定該菌株是否利用水蘇糖。結果如表3所示,共有15株菌能夠利用水蘇糖,分別為短雙歧桿菌、齒雙歧桿菌、長雙歧桿菌長亞種、假小鏈雙歧桿菌、發酵乳桿菌、副格氏乳桿菌、羅伊氏乳桿菌、瘤胃乳桿菌以及唾液乳桿菌共9個種。
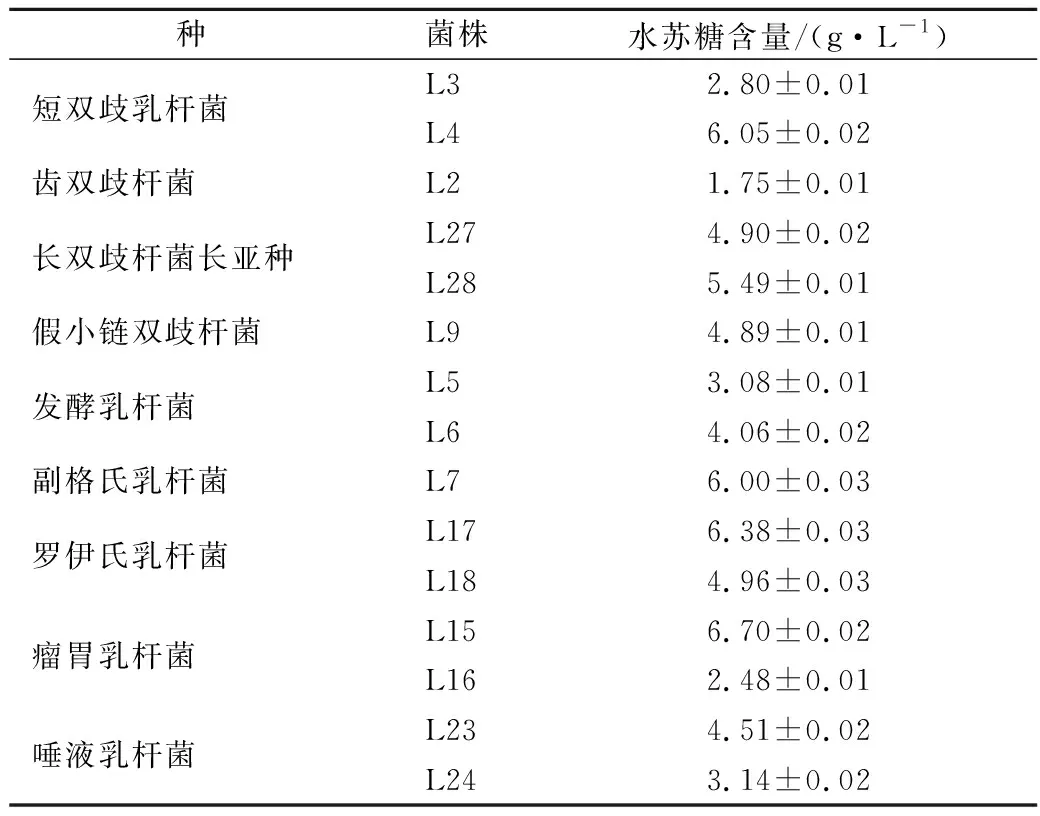
表3 細菌培養上清液中水蘇糖含量的測定Table 3 The determination of stachyose in the supernatant of bacterial culture
目前,有些菌株如短雙歧桿菌、發酵乳桿菌、羅伊氏乳桿菌等已被報道能夠利用水蘇糖[5, 14-15]。有文獻報道,副干酪乳桿菌、植物乳桿菌可以利用水蘇糖[16],而本實驗中所篩選的副干酪乳桿菌和植物乳桿菌幾乎不能利用水蘇糖生長,表明即使是同一個種,不同菌株之間也存在差異。
2.2 菌株生長曲線的測定
為研究篩選出來的腸道菌對水蘇糖的利用情況,我們從14種菌中選擇24 h內消耗水蘇糖最多的菌株即短雙歧桿菌L4、齒雙歧桿菌L2、長雙歧桿菌長亞種L28、假小鏈雙歧桿菌L9、發酵乳桿菌L6、副格氏乳桿菌L7、羅伊氏乳桿菌L17、瘤胃乳桿菌L15以及唾液乳桿菌L23,測定其在以水蘇糖作唯一碳源的GMM培養基中的生長曲線(圖1)。
由圖1可知,不同菌株在以水蘇糖作唯一碳源的GMM液體培養基中的生長存在顯著差異。菌株生長快慢與初始接種量有關,但更重要的是菌株自身的特性以及對水蘇糖利用情況。大多數菌株在培養8 h左右進入穩定期,OD600值能夠達到1.0以上,pH值下降至4.0左右。瘤胃乳桿菌L15在到達穩定期時,OD600值高達2.0,表明菌株對水蘇糖利用情況較好;短雙歧桿菌L4、副格氏乳桿菌L7菌株長勢較好,OD600值在1.6左右;齒雙歧桿菌L2、發酵乳桿菌L6生長較慢,其中發酵乳桿菌L6生長最慢,有大約8 h的延滯期,10 h之后,OD600值快速增長,一直到第18 h才進入穩定期。
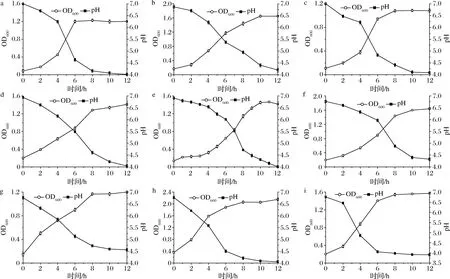
a-短雙歧桿菌L4;b-齒雙歧桿菌L2;c-長雙歧桿菌長亞種L28;d-假小鏈雙歧桿菌L9;e-發酵乳桿菌L6;f-副格氏乳桿菌L7;g-羅伊氏乳桿菌L17;h-瘤胃乳桿菌L15;i-唾液乳桿菌L23圖1 菌株在以水蘇糖作唯一碳源的 GMM 培養基中的生長曲線Fig.1 The growth curves of the strains in GMM medium with stachyose as the sole carbon source
2.3 細菌培養上清液中糖組成分析
通過測定菌株生長過程中上清液中糖組成及含量變化,探究其對水蘇糖的利用特性。隨著時間的增長,菌株對水蘇糖的消耗量增加(表4、表5)。
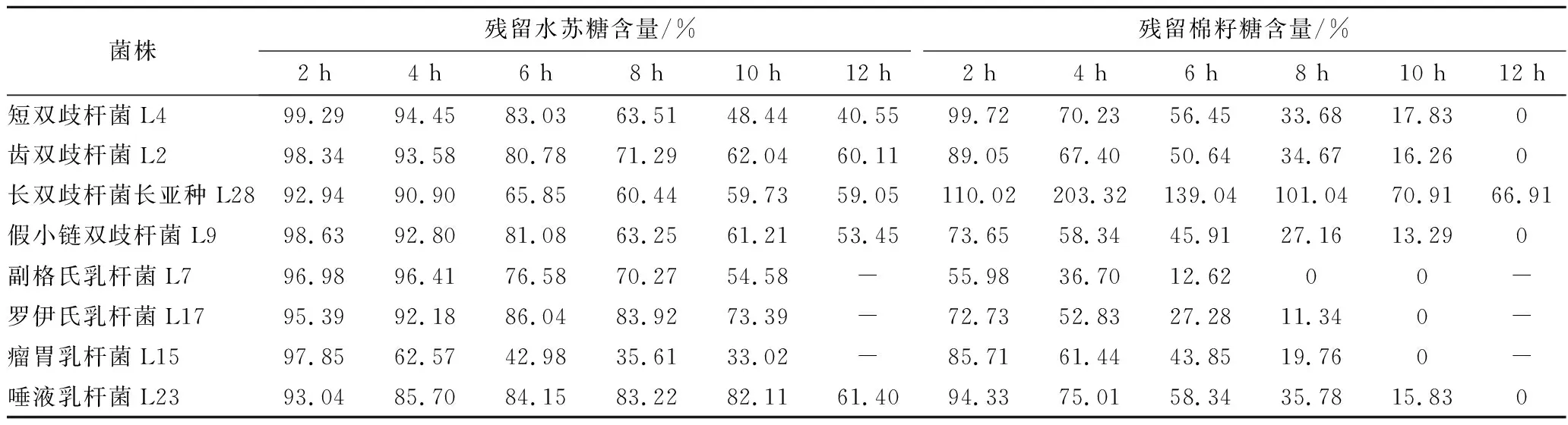
表4 不同時間菌株上清液中殘留糖含量Table 4 The content of residual stachyose in the supernatant of different strains at different times

表5 不同時間發酵乳桿菌L6上清液中殘留糖含量Table 5 The content of residual stachyose in the supernatant of L.fermentum L6 at different times
由于實驗所用的水蘇糖中含有少量棉籽糖,因此,在HPLC測定細菌上清液時,會出現棉籽糖的峰[保留時間約為(7.8±0.1)min](圖2-a)。實驗結果表明,大多數菌株隨著時間的增加,上清液中棉籽糖含量不斷減少,至不再有棉籽糖檢出。僅長雙歧桿菌長亞種L28例外,其上清液中棉子糖含量呈現了先增加后減少的趨勢,同時在4 h出現了半乳糖[保留時間約為(12.8±0.1)min](圖2)。以上結果表明菌株對水蘇糖的利用方式存在差異。
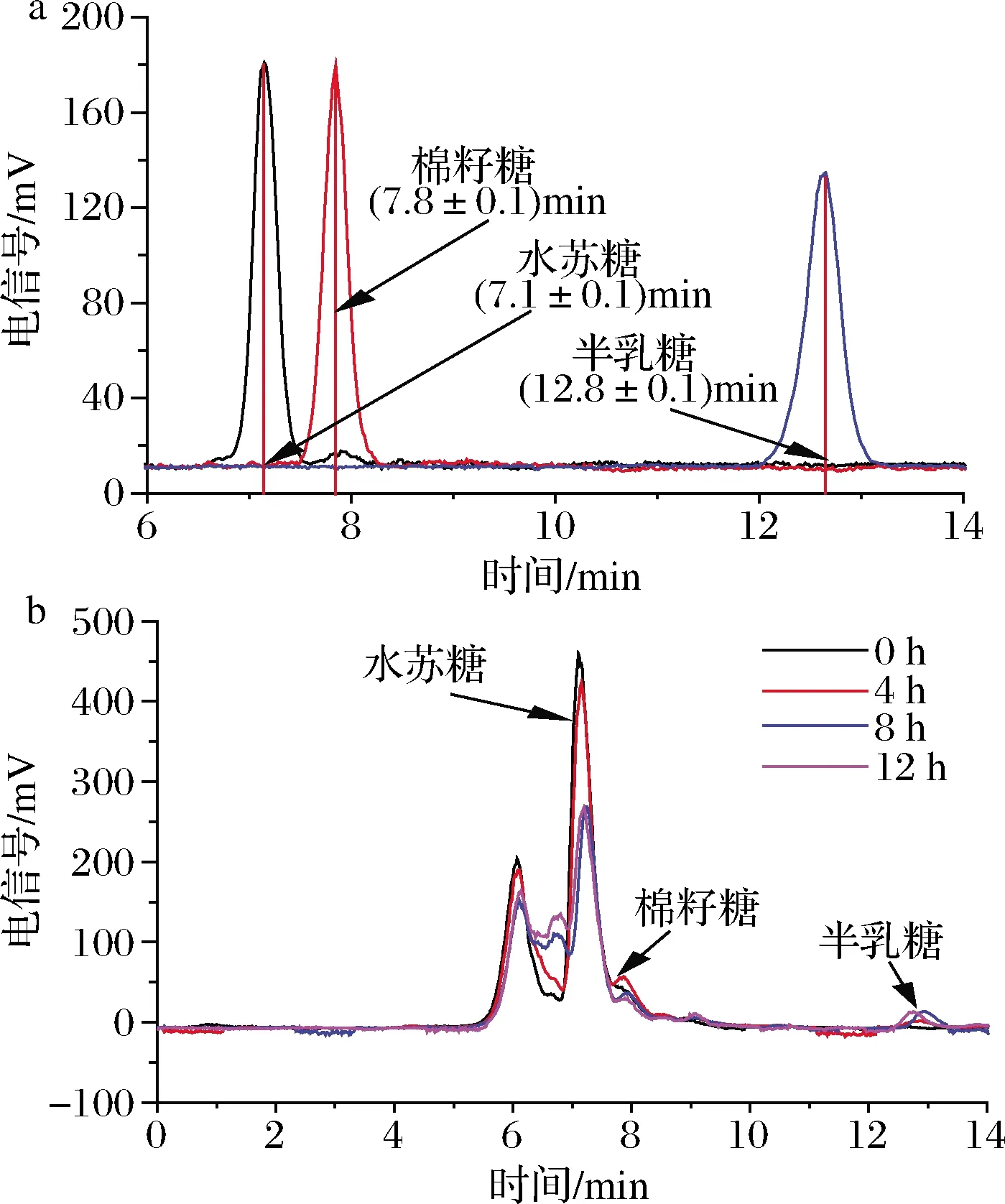
a-不同糖在HPLC中的出峰時間;b-不同時間長雙歧桿菌長亞種L28上清液中糖含量變化圖2 長雙歧桿菌長亞種L28培養上清液中糖組成的HPLC分析Fig.2 The composition of sugars in the supernatant of B.longum spp.longum L28 by HPLC analysis
2.4 α-半乳糖苷酶酶活力的測定
腸道細菌水解水蘇糖需要α-半乳糖苷酶,DONKOR等[17]測定了雙歧桿菌、乳桿菌胞內α-半乳糖苷酶酶活力,發現乳酸雙歧桿菌B94酶活力最高,水蘇糖水解率也最高。實驗中瘤胃乳桿菌L15的α-半乳糖苷酶活力最高,24 h內消耗的水蘇糖也最多,約為67%。本實驗通過比較胞內外酶活力,以確定酶所在位置,進一步探究其對水蘇糖的利用特性。選擇生長至穩定期的菌泥與上清液進行酶活力測定,由表6可知,實驗菌株以胞內α-半乳糖苷酶為主。短雙歧桿菌L4胞內、外酶活力相差最大,胞內約是胞外的2 973倍;齒雙歧桿菌L2胞內、外酶活力相差最小,胞內約為胞外的8倍;長雙歧桿菌長亞種L28、瘤胃乳桿菌L15、發酵乳桿菌L6、唾液乳桿菌L23胞內酶活力約為胞外的100~360倍。
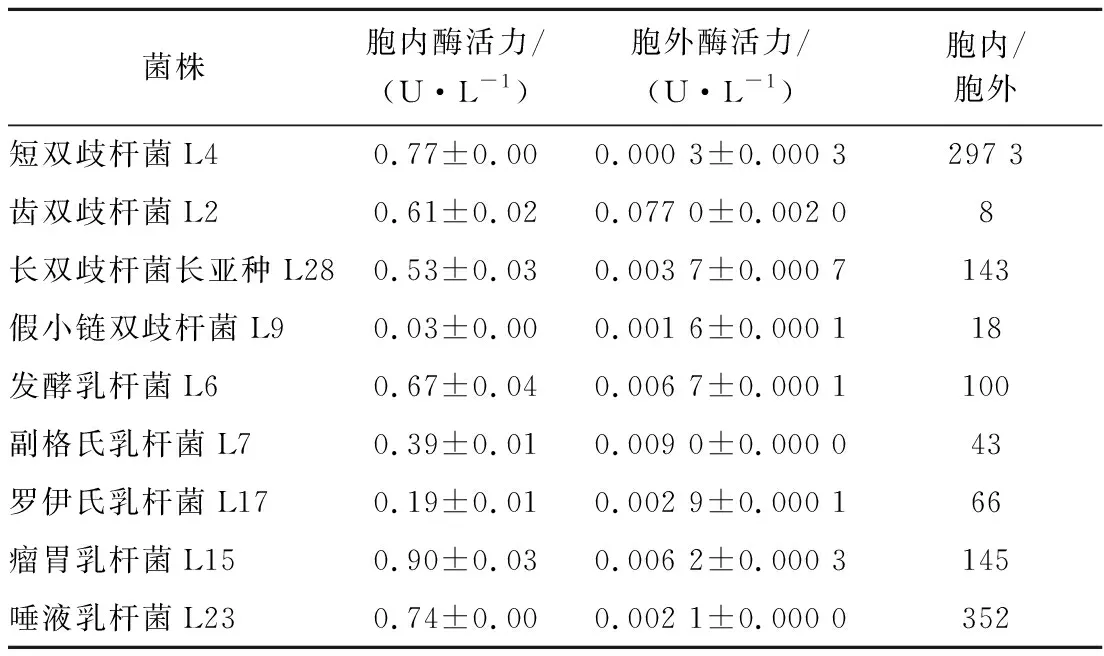
表6 α-半乳糖苷酶酶活力的測定Table 6 The determination of α-galactosidase enzyme activity
2.5 基因組草圖分析
實驗菌株均已完成基因組草圖測序,由KEGG注釋結果可知,水蘇糖代謝屬于半乳糖代謝路徑,其代謝通路涉及了α-半乳糖苷酶(galA)、β-呋喃果糖苷酶(sacA)、α-葡萄糖苷酶(malZ)以及低聚-1,6-葡糖苷酶(malL)(圖3)。我們從NCBI上下載了2株能夠代謝水蘇糖的雙歧桿菌和乳桿菌的基因組信息,分別為動物雙歧桿菌乳酸亞種ATCC SD5219和副格氏乳桿菌JCM5343,并與實驗菌株進行比較。結果顯示,所有菌株都具有α-半乳糖苷酶,但并不是所有菌株都含有β-呋喃果糖苷酶、α-葡萄糖苷酶以及低聚-1,6-葡糖苷酶,而且即使是同一種的菌株(副格氏乳桿菌JCM5343和副格氏乳桿菌L7)基因數量也有可能不同(表7)。因此,不同菌株對水蘇糖的利用方式不同。
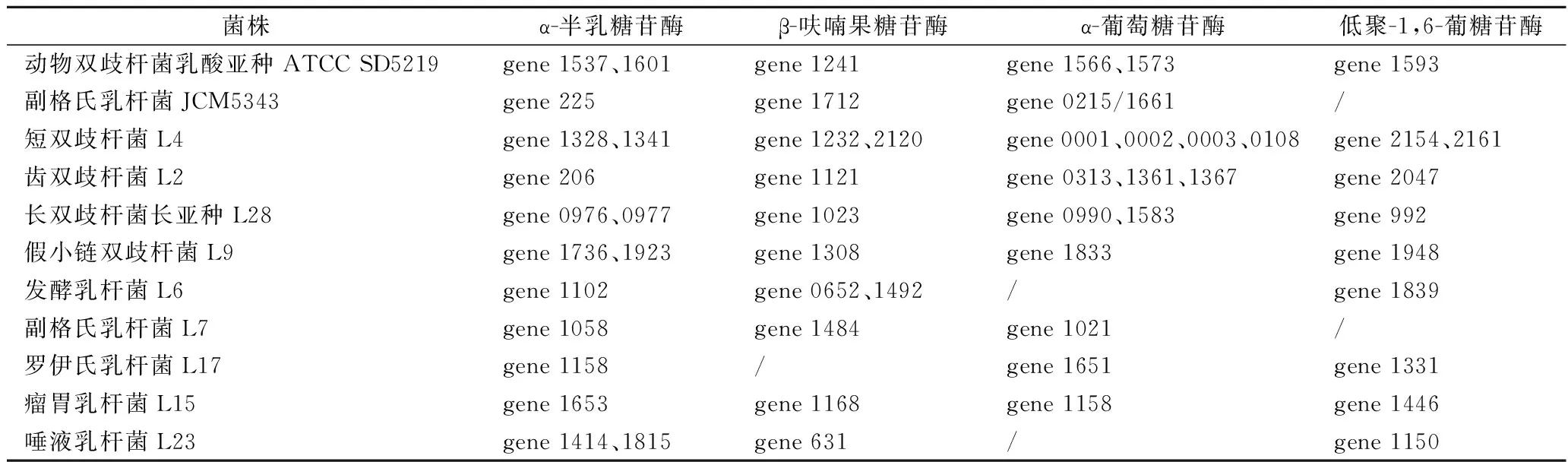
表7 菌株代謝水蘇糖相關酶的基因信息Table 7 The information on enzymes related to stachyose metabolism by strains
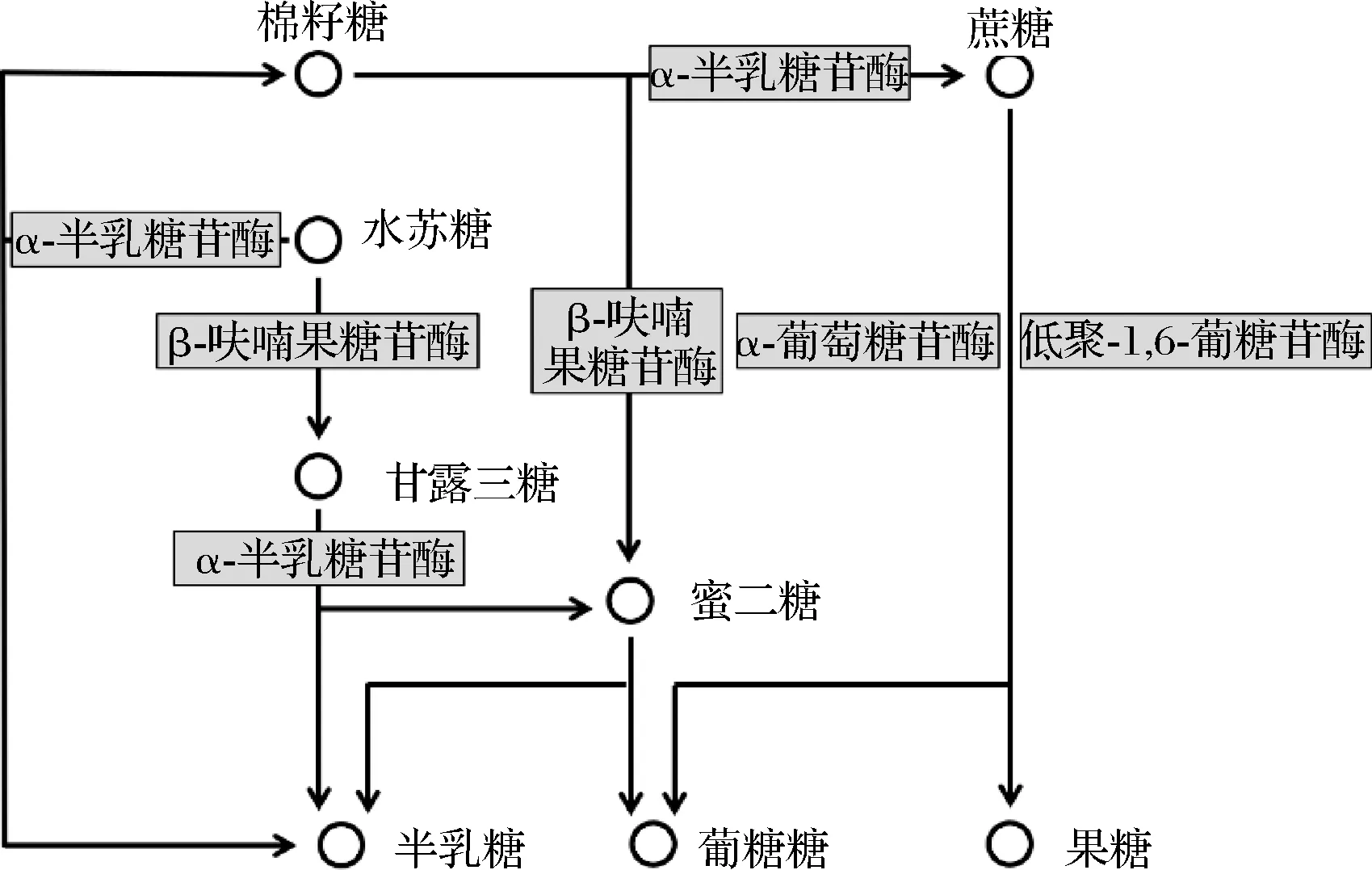
圖3 水蘇糖的代謝路徑Fig.3 The metabolic pathways of stachyose
我們利用LOCTREE對9株菌株的α-半乳糖苷酶進行亞細胞定位分析。除長雙歧桿菌長亞種L28和羅伊氏乳桿菌L17外,大部分實驗菌株的α-半乳糖苷酶均存在于細胞質,與酶活力測定結果基本一致(表8)。長雙歧桿菌長亞種L28有2個α-半乳糖苷酶基因,根據LOCTREE預測結果顯示,1個在細胞質中,1個存在于細胞外。酶活力結果顯示長雙歧桿菌長亞種L28的胞內酶活力是胞外的143倍,可能是胞內α-半乳糖苷酶的含量遠高于胞外。羅伊氏乳桿菌L17僅有1個α-半乳糖苷酶基因,且系統預測該酶存在于細胞外,與酶活力測定結果不符,可能是因為酶活力測定時大量α-半乳糖苷酶尚未分泌到細胞外。
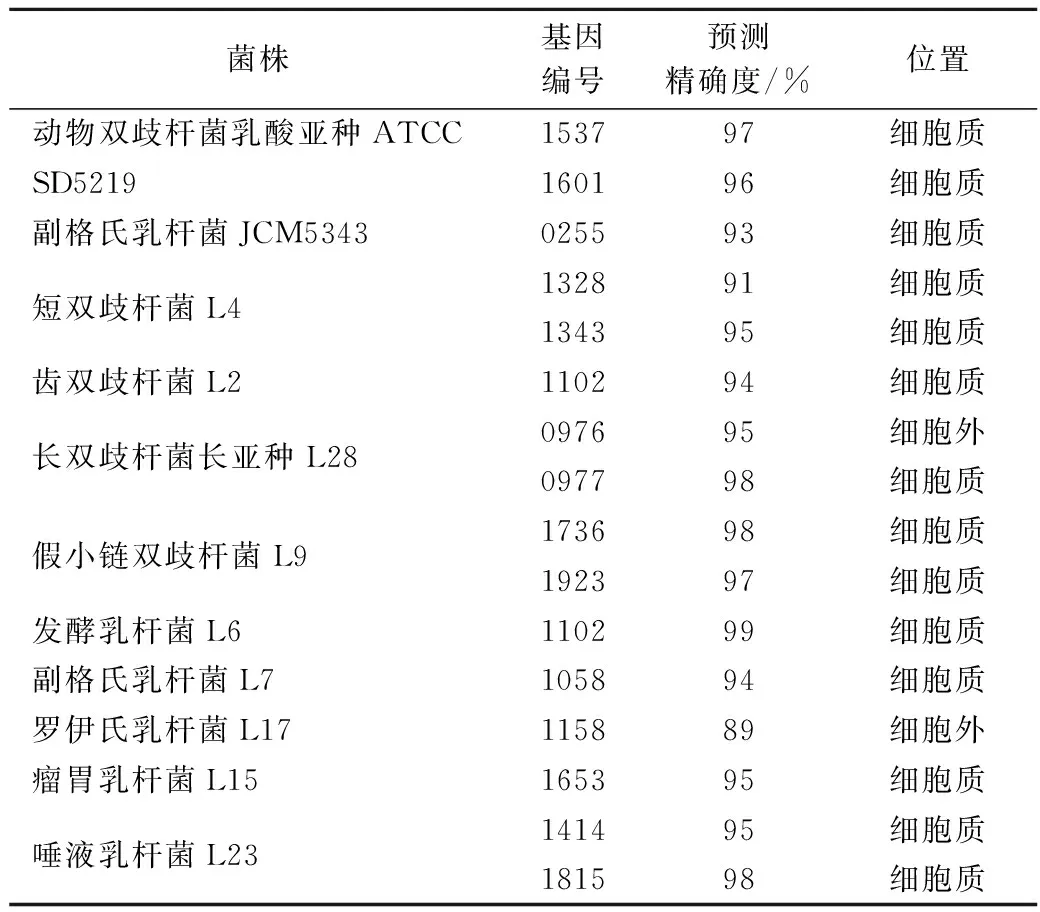
表8 α-半乳糖苷酶的亞細胞定位分析Table 8 The subcellular localization analysis of α-galactosidase
酶活力測定以及LOCTREE預測結果顯示實驗菌株均以胞內α-半乳糖苷酶為主,因此需要相關轉運蛋白將水蘇糖轉運至胞內。已有研究表明,變形鏈球菌具有多糖代謝轉運體,能夠利用其轉運水蘇糖、棉籽糖、蜜二糖等。該轉運體由底物結合蛋白(msmE)、2個膜蛋白(msmF,msmG)以及ATP-結合蛋白(msmK)構成[18-19]。之前的研究證明一些腸道菌(如屎腸球菌、糞腸球菌)中存在msmEFG基因,當缺失msmK基因時可能并不影響水蘇糖的轉運[9]。事實上,msmEFGK轉運體屬于ABC(ATP-binding cassette)家族的成員[19]。雙歧桿菌屬已被報道具有msmEFG轉運體,并證明該轉運體能夠轉運水蘇糖等寡糖[20-22]。本實驗研究的雙歧桿菌中同樣檢測到了msmEFG基因,但乳桿菌中并不存在。
實驗結果顯示,雙歧桿菌屬在代謝水蘇糖的基因方面比較相似,是由msmEFG轉運體負責將水蘇糖轉運至細胞內,再由胞內的4種酶將水蘇糖水解成單糖進行利用。其中,長雙歧桿菌長亞種L28相對比較特殊。從表觀現象來看,HPLC結果表明該菌株生長過程中胞外上清液棉籽糖含量呈現先增加后減少的趨勢,同時出現了半乳糖。從基因方面來看,長雙歧桿菌長亞種L28代謝水蘇糖的相關基因片段更為復雜(圖4),除重復出現galA和msmEFG基因之外,還有NagC轉錄調節因子和lacI轉錄調節因子,它們是否表達,是否對利用水蘇糖有影響,目前尚不可知。根據現有結果,推測長雙歧桿菌長亞種L28利用水蘇糖的方式可能是:水蘇糖在胞外被水解產生棉籽糖和半乳糖,導致其生長過程中出現了棉籽糖以及半乳糖含量的增加;而后期棉籽糖含量減少可能是被轉運至細胞內利用,而不是繼續在胞外水解,因為并未在上清液中檢測到蜜二糖等物質。另外,在模式菌株動物雙歧桿菌乳酸亞種ATCC SD5219中,galA基因附近除連續的msmEFG基因外,還出現了NagC轉錄調節因子,表明該基因可能參與調節水蘇糖代謝。
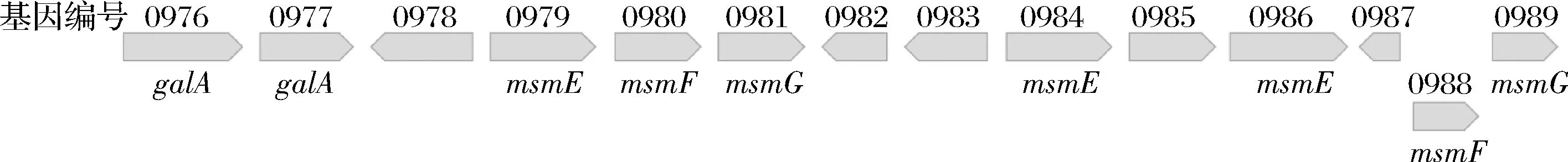
圖4 長雙歧桿菌長亞種L28中與水蘇糖利用相關的基因簇Fig.4 The gene clusters related to stachyose utilization in B.longum ssp. longum L28
在乳桿菌屬5株菌的基因組草圖注釋結果中,并沒有找到msmEFG基因,說明乳桿菌轉運水蘇糖的方式與雙歧桿菌屬不同。分析乳桿菌基因組草圖,列出可能轉運水蘇糖的潛在基因(表9)。副格氏乳桿菌JCM5343和副格氏乳桿菌L7結果相似,擁有類似msmEFGK的體系,由3個假定蛋白組成:ABC.X4.S(ABC轉運系統底物結合蛋白)、ABC.X4.P(ABC轉運系統通透酶)、ABC.X4.A(ABC轉運系統ATP結合蛋白),利用這3個基因指導合成的蛋白質轉運水蘇糖。發酵乳桿菌L6、瘤胃乳桿菌L15、唾液乳桿菌L23可能是通過lacS基因編碼的通透酶將水蘇糖運輸至胞內利用。最初我們認為羅伊氏乳桿菌L17極有可能是利用yagU(假定通透酶)轉運水蘇糖,但是結合α-半乳糖苷酶的亞細胞定位分析,該基因所產生的通透酶可能是α-半乳糖苷酶分泌到細胞外的通道或載體;其后續利用可能也與其他菌株不同,因為羅伊氏乳桿菌L17并不存在β-呋喃果糖苷酶,可能是利用α-半乳糖苷酶水解成蔗糖,再由α-葡萄糖苷酶或低聚-1,6-葡糖苷酶進一步分解利用。
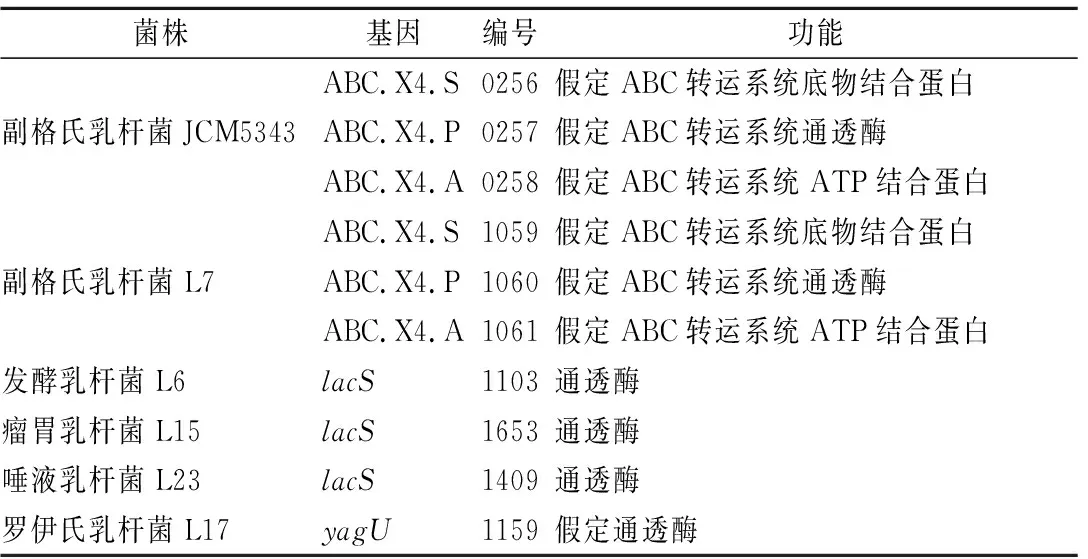
表9 乳酸菌中轉運水蘇糖的潛在基因Table 9 The potential genes for stachyose transport in Lactobacillus
綜上所述,雙歧桿菌、乳桿菌大多通過轉運蛋白將水蘇糖轉運至胞內水解利用,但是不同種屬的菌株利用水蘇糖的基因不同。雙歧桿菌主要是通過msmEFG轉運體轉運水蘇糖,再由胞內的多種酶將其水解;乳桿菌轉運水蘇糖可能是利用了ABC轉運系統或其他通透酶。可能也存在一些特例,例如,長雙歧桿菌長亞種L28可能是利用胞外α-半乳糖苷酶將水蘇糖水解成棉籽糖和半乳糖,再通過msmEFG轉運體棉籽糖轉運至胞內利用。羅伊氏乳桿菌L17分泌胞外α-半乳糖苷酶將水蘇糖水解成蔗糖,再利用α-葡萄糖苷酶或低聚-1,6-葡糖苷酶將其徹底分解利用。
3 結論
本研究分析了人類糞便來源的雙歧桿菌、乳桿菌對水蘇糖的利用情況及相關代謝基因,得到的實驗結果表明,不同菌株對水蘇糖的利用方式存在差異,產氣機理也不同。雙歧桿菌主要通過msmEFG轉運體轉運水蘇糖至胞內,然后由α-半乳糖苷酶、β-呋喃果糖苷酶、α-葡萄糖苷酶或低聚-1,6-葡糖苷酶將其水解利用;乳桿菌主要是通過各種透性酶及其他轉運蛋白轉運水蘇糖至胞內進行利用。此外,如長雙歧桿菌長亞種和羅伊氏乳桿菌,這些菌株可能存在胞外α-半乳糖苷酶,可在細胞外將水蘇糖初步水解以便進一步利用。研究腸道細菌對水蘇糖的利用方式,篩選出能利用水蘇糖但不產氣的益生菌,可以指導可食用菌種與水蘇糖進行合理搭配,為開發功能性產品提供思路。今后可以更深入地研究腸道細菌對水蘇糖的利用機制,揭示攝入過量水蘇糖引起人體不適的機理,進一步闡明水蘇糖與人體健康的關系。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
發明與創新(2016年38期)2016-08-22 03:02:52
