化學物質誘導對馬氏珠母貝眼點幼蟲附著的影響
2022-01-19 02:40:00楊創業章佳斌曾業濤廖永山鄧岳文王慶恒
廣東海洋大學學報 2022年1期
楊創業,章佳斌,曾業濤,廖永山,鄧岳文,3,王慶恒,3
化學物質誘導對馬氏珠母貝眼點幼蟲附著的影響
楊創業1,章佳斌1,曾業濤1,廖永山2,鄧岳文1,3,王慶恒1,3
(1. 廣東海洋大學水產學院 // 2. 廣東海洋大學珍珠研究所 // 3. 廣東省珍珠養殖與加工工程技術研究中心,廣東 湛江 524088)
【目的】研究血清素(5-HT)、蛻皮激素、γ-氨基丁酸(GABA)、腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤對馬氏珠母貝眼點幼蟲附著的影響。【方法】以不同濃度的5-HT、蛻皮激素、GABA、腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤溶液處理馬氏珠母貝眼點幼蟲,不同時間后,分析眼點幼蟲附著率的變化。【結果】在24 ~ 96 h作用時間里,5-HT、蛻皮激素、GABA對眼點幼蟲附著的誘導效果整體上均呈上升趨勢;在24 ~ 120 h作用時間里,腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤對眼點幼蟲附著的誘導效果整體上呈上升趨勢。眼點幼蟲在10-5mol/L的5-HT和GABA處理24、48、72和96 h后,附著率均顯著高于對照組(< 0.01);10-5mol/L的蛻皮激素處理24 h和72 h,10-6mol/L的蛻皮激素處理24 h和96 h,10-7mol/L的蛻皮激素處理24 h,附著率均顯著高于對照組(< 0.01);10 μmol/L腺嘌呤核苷處理24、48、72和120 h,1 μmol/L腺嘌呤核苷處理24、48、72、96和120 h,附著率均顯著高于對照組(< 0.01);10 μmol/L次黃嘌呤核苷處理48和72 h,1 μmol/L次黃嘌呤核苷處理48、72、96和120 h,附著率均顯著高于對照組(< 0.01);10 μmol/L的次黃嘌呤處理72、96和120 h,附著率均顯著高于對照組(< 0.01)。【結論】5-HT、蛻皮激素、GABA、腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤溶液在適宜的濃度和處理時間均可誘導馬氏珠母貝眼點幼蟲附著,其中10-5mol/L的5-HT處理24 h對附著率的誘導效果最佳,可達對照組的6.3倍。
馬氏珠母貝;眼點幼蟲;附著;化學物質
貝類附著變態階段幼蟲對環境因素異常敏感,常因受到不良環境因素的影響,出現變態延遲或大量死亡,死亡率有時可以高達80% ~ 90%,給生產造成巨大損失[1]。在貝類苗種培育過程中,以優化育苗技術方案來促進浮游幼蟲大量同步定向附著和變態,可顯著提高育苗的產量和效益。研究表明,物理環境誘導(如基質、水流、溫度和鹽度等)[2-4]、生物誘導[1, 5-6]及化學物質誘導[7-9]等均影響著貝類幼蟲的附著變態。然而,同一種藥物對于雙殼貝類,甚至同屬并不具備有普適性。例如,腎上腺素在厚殼貽貝()眼點幼蟲具誘導作用[10],而在紫貽貝()上卻沒有作用[11]。因此,尋找促進特定貝類附著變態的有效解決方案具有重要意義。
馬氏珠母貝()是我國南方沿海特色養殖貝類,主要用于培育海水珍珠,同時具有較高營養和藥用價值[12],然而,苗種生產中的幼蟲附著變態率低的問題一直是限制馬氏珠母貝產業發展的主要因素之一。為縮短幼蟲附著變態時間,提高幼蟲附著變態率,本團隊前期利用代謝組結合轉錄組技術研究了馬氏珠母貝幼蟲附著變態過程調控特征,篩選了參與附著變態過程的差異代謝物,包括血清素(5-HT)、γ-氨基丁酸(GABA)、蛻皮激素、腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤等[13]。本研究在此基礎上探討了以上差異代謝物對馬氏珠母貝眼點幼蟲附著的影響,以期為高效苗種培育提供技術指導。
1 材料和方法
1.1 實驗材料
從本團隊培育的馬氏珠母貝黑殼色品系中挑出性腺成熟的個體,清洗貝體表面的附著物,剖取馬氏珠母貝的精子和卵子,將收集到的精子與卵子在砂濾海水中混合,使其受精(溫度25 °C ± 1 °C,鹽度30 ± 1)。完成受精后,使用砂濾海水洗滌受精卵3 ~ 4次,去除多余的精子和未成熟的卵子,并將卵移入新的砂濾海水中進行早期培育。受精后21 d,幼蟲后腹部口與肛門之間出現足和鰓的原基,在足原基的基部前方兩側靠近貝殼的位置出現圓形或橢圓形的眼點,選取此時期幼蟲作為實驗材料。
1.2 實驗方法
實驗開始前,使用過濾海水分別配制成5-HT、蛻皮激素 [助溶劑為0.9 μL/L二甲基亞砜 (DMSO)溶液]、GABA、腺嘌呤核苷、次黃嘌呤核苷和次黃嘌呤的儲備液。5-HT設計3個濃度實驗梯度,分別是10-5、10-6、10-7mol/L和1個空白對照組,作用時間為24、48、72和96 h;蛻皮激素設計3個濃度實驗梯度,分別是10-5、10-6和10-7mol/L,對照組為1個空白對照組和0.9 μL/L DMSO溶液組,作用時間為24、48、72和96 h;GABA設計3個濃度實驗梯度,分別是10-5、10-6、10-7mol/L和1個空白對照組,作用時間為24、48、72和96 h;腺嘌呤核苷設計3個濃度實驗梯度,分別是10-4、10-5、10-6mol/L和1個空白對照組,作用時間為24、48、72、96和120 h;次黃嘌呤核苷設計3個濃度實驗梯度,分別是10-4、10-5、10-6mol/L和1個空白對照組,作用時間為24、48、72、96和120 h;次黃嘌呤設計3個濃度實驗梯度,分別是10-4、10-5、10-6mol/L和1個空白對照組,作用時間為24、48、72、96和120 h。
實驗在60 mL的一次性培養皿中進行,各培養皿中放置40 mL過濾海水,每個培養皿幼蟲數為80 ~ 130只,并通過微型蠕動泵進行增氧,每12 h投喂湛江等鞭金藻(),各組處理設置3個重復。
通過對附著在培養皿中的幼蟲進行計數并統計附著率(附著率= 附著個體數/實驗個體總數),具體技術方法參照Yu等[14]。
1.3 數據處理
數據運用EXCEL進行處理,將獲得的附著率百分比數值進行平方根的反正弦轉換,以增加數據的正態和方差齊性。采用SPSS 26. 0對所得數據進行單因素方差分析,若差異達到顯著,則進行Tukey多重比較,顯著性水平= 0.05。
2 結果
2.1 5-HT對馬氏珠母貝眼點幼蟲附著率的影響
在24 ~ 96 h作用時間里,5-HT對馬氏珠母貝眼點幼蟲附著的誘導效果整體呈上升趨勢(圖1)。其中,10-5mol/L的5-HT對幼蟲附著的誘導效果最佳,10-5mol/L的5-HT處理24、48、72和96 h后,幼蟲的附著率均顯著高于對照組(< 0.05),附著率分別是對照組的6.3倍、6.0倍、3.9倍和2.0倍;10-6mol/L和10-7mol/L的5-HT對馬氏珠母貝眼點幼蟲附著率無顯著提高(0.05)。
2.2 蛻皮激素對馬氏珠母貝眼點幼蟲附著率影響
在24 ~ 96 h作用時間里,蛻皮激素對馬氏珠母貝眼點幼蟲附著的誘導效果整體上呈上升趨勢(圖2)。10-5mol/L的蛻皮激素處理24 h和72 h后,眼點幼蟲附著率均顯著高于對照組和DMSO組(< 0.05),附著率分別是對照組的2.4倍和2.6倍;10-6mol/L的蛻皮激素處理24 h和96 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組的2.0倍和2.1倍;10-7mol/L的蛻皮激素處理24 h后,幼蟲的附著率顯著高于對照組(< 0.05),附著率是對照組的2.5倍。
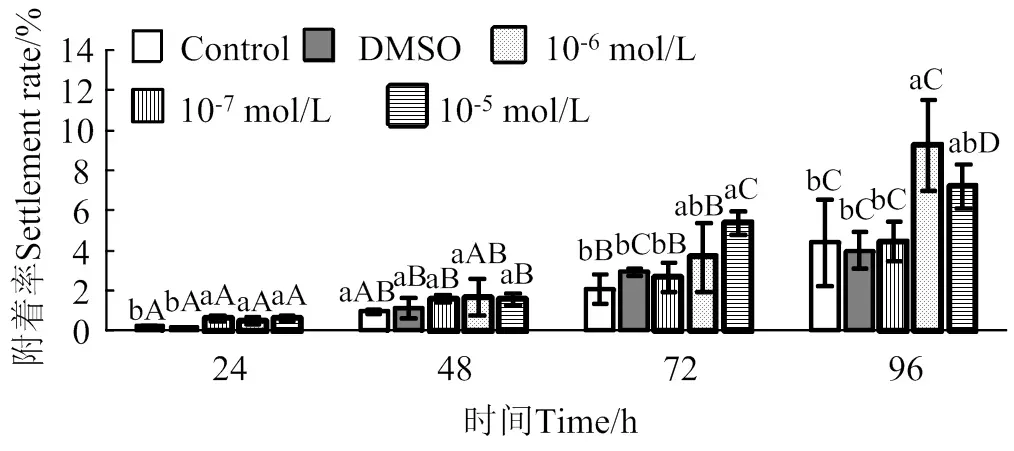
相同濃度處理組在不同時間之間,凡含一個相同大寫字母,表示差異不顯著(P > 0.05),相同時間里不同濃度處理組之間,凡含一個相同小寫字母表示差異不顯著(P > 0.05)
2.3 GABA對馬氏珠母貝眼點幼蟲附著率的影響
在24 ~ 96 h作用時間里,GABA對馬氏珠母貝眼點幼蟲附著的誘導效果整體呈上升趨勢(圖3)。10-5mol/L的GABA對幼蟲附著的誘導效果最佳,10-5mol/L的GABA處理24、48、72和96 h后,幼蟲的附著率均顯著高于對照組(< 0.05),附著率分別是對照組的5.5倍、3.0倍、2.8倍和2.2倍;10-6mol/L和10-7mol/L的GABA對馬氏珠母貝眼點幼蟲附著率無顯著提高(> 0.05)。
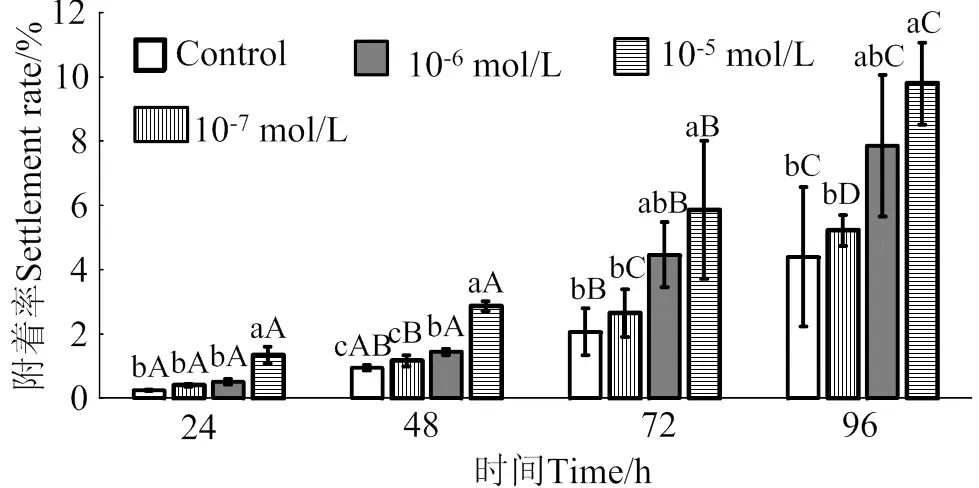
相同濃度處理組在不同時間之間,凡含一個相同大寫字母,表示差異不顯著(P > 0.05),相同時間里不同濃度處理組之間,凡含一個相同小寫字母表示差異不顯著(P > 0.05)
2.4 腺嘌呤核苷對馬氏珠母貝眼點幼蟲附著率影響
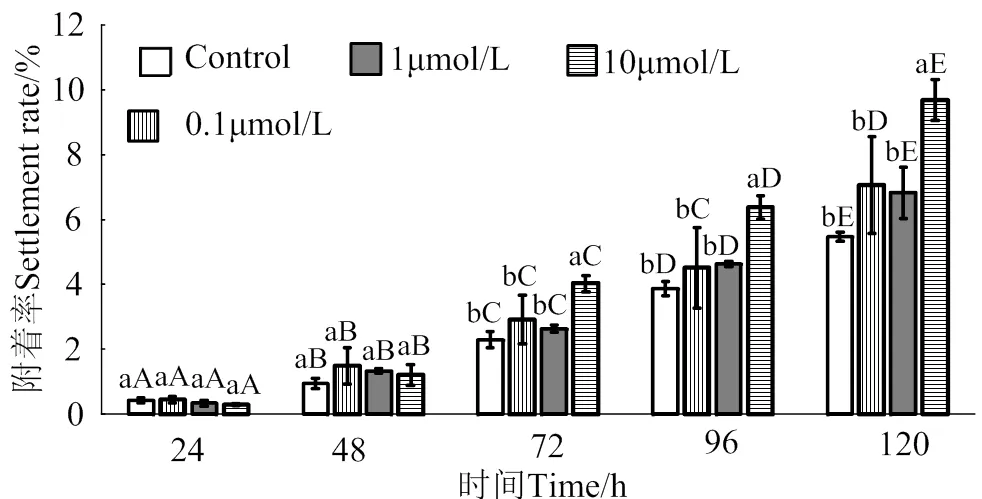
在24 ~ 120 h作用時間里,腺嘌呤核苷對馬氏珠母貝眼點幼蟲附著的誘導效果整體呈上升趨勢(圖4)。10 μmol/L的腺嘌呤核苷處理24、48、72和120 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組的2.1倍、3.5倍、2.7倍和2.1倍;1 μmol/L的腺嘌呤核苷處理24、48、72、96和120 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組的1.9倍、3.0倍、2.6倍、2.1倍和2.3倍;0.1 μmol/L的腺嘌呤核苷對馬氏珠母貝眼點幼蟲附著率無顯著提高(> 0.05)。
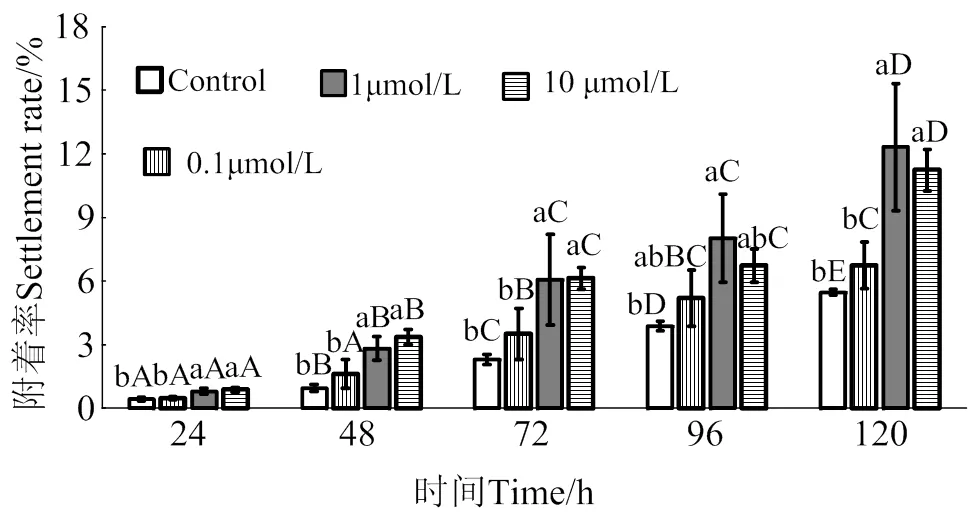
相同濃度處理組在不同時間之間,凡含一個相同大寫字母,表示差異不顯著(P > 0.05),相同時間里不同濃度處理組之間,凡含一個相同小寫字母表示差異不顯著(P > 0.05)
2.5 次黃嘌呤核苷對馬氏珠母貝眼點幼蟲附著率影響
在24 ~ 120 h作用時間里,次黃嘌呤核苷對馬氏珠母貝眼點幼蟲附著誘導效果整體呈上升趨勢(圖5)。10 μmol/L次黃嘌呤核苷處理48 h和72 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組2.4倍和1.8倍;1 μmol/L次黃嘌呤核苷處理48、72、96和120 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組1.8倍、1.5倍、1.3倍和1.3倍;0.1 μmol/L次黃嘌呤核苷對馬氏珠母貝眼點幼蟲附著率無顯著提高(> 0.05)。
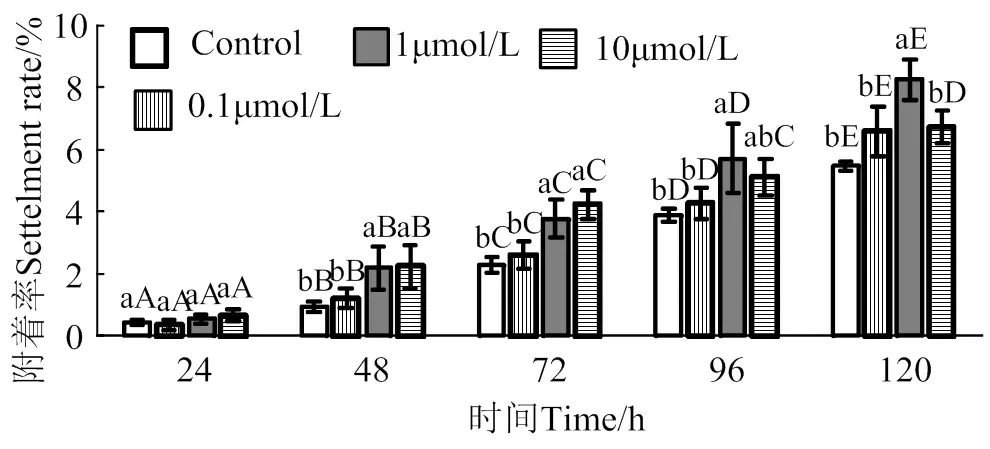
相同濃度處理組在不同時間之間,凡含一個相同大寫字母,表示差異不顯著(P > 0.05),相同時間里不同濃度處理組之間,凡含一個相同小寫字母表示差異不顯著(P > 0.05)
2.6 次黃嘌呤對馬氏珠母貝眼點幼蟲附著率影響
在24 ~ 120 h作用時間里,次黃嘌呤對馬氏珠母貝眼點幼蟲附著的誘導效果整體呈上升趨勢(圖6)。10 μmol/L的次黃嘌呤處理72、96和120 h后,幼蟲附著率均顯著高于對照組(< 0.05),附著率分別是對照組的1.8倍、1.6倍和1.8倍;0.1 μmol/L和1 μmol/L的次黃嘌呤對馬氏珠母貝眼點幼蟲附著率無顯著提高(> 0.05)。
相同濃度處理組在不同時間之間,凡含一個相同大寫字母,表示差異不顯著(P > 0.05),相同時間里不同濃度處理組之間,凡含一個相同小寫字母表示差異不顯著(P > 0.05)
3 討論
在水產經濟貝類的苗種生產中,幼蟲附著變態的成功與否直接決定出苗量和育苗的成敗。提高幼蟲附著變態率是解決這一問題的主要途徑之一。化學物質通過對軟體動物行為方式或生活方式的影響,可使更多幼蟲附著到基質上。因此,幼蟲一旦探測到化學信號,就可能會改變其垂直分布,使幼蟲集中分布在水體的底部或附著在附著基上[15]。
5-HT是一種神經遞質和調節劑,在無脊椎動物的神經系統中起著重要的作用[16]。研究表明,5-HT可以誘導大珠母貝()[17]和羊鮑()[8]等幼蟲的變態發育。本研究中,10-5mol/L的5-HT處理24、48、72和96 h后,馬氏珠母貝眼點幼蟲的附著率均顯著高于對照組,最高可達對照組的6.3倍。該結果與Yu等[14]的誘導結果相似;同時與本團隊馬氏珠母貝眼點幼蟲附著變態前后轉錄組與代謝組研究中血清素及血清素受體相關基因的變化相呼應[13]。
蛻皮激素是節肢動物幼蟲啟動變態過程中最重要的激素之一,蛻皮激素與蛻皮激素受體觸發昆蟲變態的一系列級聯反應[18]。蛻皮激素受體最初被認為是蛻皮動物所特有,但現有基因組和轉錄組數據的分析顯示,蛻皮激素受體以及蛻皮激素級聯反應相關轉錄因子同樣存在于軟體動物,例如,長牡蠣()[19]、青螺()[20]和光滑雙臍螺()[21]。Vogeler等[22]研究發現蛻皮激素在長牡蠣()變態前的面盤幼蟲中高表達,推測了蛻皮激素在雙殼類變態中起到誘導作用。Chelikowsk等[23]在對海灣扇貝()的研究中發現,合成的蛻皮激素與pavlovol(微藻屬的一種甾醇,與蛻皮激素結構相似)能夠誘導海灣扇貝幼蟲的變態。本團隊前期的代謝組分析發現蛻皮激素在進入稚貝期后發生下調;并且甾醇激素生物合成通路在幼蟲附著期到稚貝期之間被富集[13]。本研究結果顯示,適宜濃度的蛻皮激素能夠引起馬氏珠母貝眼點幼蟲附著率上升,進一步證明馬氏珠母貝的附著受到蛻皮激素的調控。
GABA是一種重要的抑制性氨基酸類神經遞質,它是由谷氨酸經谷氨酸脫羧酶催化脫羧基產生,GABA敏感細胞的去極化可能參與幼體的附著變態,并在多種貝類中發揮作用[24-25]。然而,GABA可能對一些貝類的幼蟲產生毒性作用,例如綠唇貽貝()[26];Young等[26]推測該毒性作用是由于GABA受體導致的神經元膜的抑制性超極化。但是GABA在適宜濃度時,其誘導效果顯著,且無毒副作用。本研究中,10-5mol/L的GABA處理24、48、72和96 h后,馬氏珠母貝眼點幼蟲的附著率均顯著高于對照組,最高可達對照組的5.5倍。這與誘導西施舌幼蟲附著變態的GABA適宜濃度相一致[27]。
嘌呤類物質可以作為一種信號分子影響魚類與多毛類的生理活動。例如,ATP、ADP、AMP和腺嘌呤核苷可以提高魚類的食欲,而次黃嘌呤3-N-氧化物可以引起魚類的回避行為或警惕行為[28-29];嘌呤代謝物尿酸和次黃嘌呤核苷可以分別作為精子和卵子的釋放信息素[30-31]。He等[32]研究發現腺嘌呤核苷、次黃嘌呤核苷與次黃嘌呤作為信息素,可以誘導沙篩貝()幼蟲的沉降形成聚集,并且三種嘌呤共同作用的誘導效果比單一嘌呤的作用更強,且沙篩貝對特定比例的嘌呤混合物最為敏感。本研究發現腺嘌呤核苷、次黃嘌呤核苷與次黃嘌呤對馬氏珠母貝幼蟲的附著存在不同程度的誘導作用,推測嘌呤可能作為一種信號分子誘導馬氏珠母貝幼蟲附著。但是,是否存在誘導幼蟲附著的最佳特定比例及嘌呤類化合物是如何引起馬氏珠母貝幼蟲變態等問題還需要進一步研究。
本研究中,5-HT、蛻皮激素、GABA、腺嘌呤核苷酸、次黃嘌呤核苷酸和次黃嘌呤溶液在適宜的濃度和處理時間均可誘導馬氏珠母貝眼點幼蟲附著,其中10-5mol/L的5-HT處理24 h對附著率的誘導效果最佳,可達對照組的6.3倍。研究結果為珍珠貝幼蟲附著變態研究奠定基礎,有助于為高效的商業化苗種培育提供技術支撐。
[1] 杜美榮, 方建光, 毛玉澤, 等. 底棲硅藻生物膜附著基對扇貝幼蟲附著和變態的影響[J]. 海洋與湖沼, 2020, 51(1): 125-131.
[2] VELASCO L A, BARROS J. Survival and growth of hatchery-produced postlarvae and spat of the Caribbean scallopsand[J]. Aquaculture Research, 2009, 40(3): 362-375.
[3] 胡冰, 米成志, 閆喜武, 等. 水溫及密度對皺紋盤鮑()幼蟲運輸存活率和變態率的影響[J]. 海洋通報, 2017, 36(3): 320-325.
[4] 梁飛龍, 林偉財, 鄧岳文, 等. 換水頻率和附著基對大珠母貝育苗效果的影響[J]. 中國農學通報, 2015, 31(29): 33-38.
[5] 蔡雨珊, 張秀坤, 竹攸汀, 等. 海假交替單胞菌()鞭毛蛋白對生物被膜形成及厚殼貽貝附著的影響[J]. 海洋學報, 2021, 43(4): 75-83.
[6] VASQUEZ H E, HASHIMOTO K, YOSHIDA A, et al. A glycoprotein in shells of conspecifics induces larval settlement of the Pacific oyster[J]. PLoS One, 2013, 8(12): e82358.
[7] YANG J L, LI S H, BAO W Y, et al. Effect of different ions on larval metamorphosis of the mussel[J]. Aquaculture Research, 2015, 46(1): 155-162.
[8] 浦舒為, 逯峰, 王佳偉, 等. γ-氨基丁酸、氯化鉀和5-羥色胺對羊鮑()幼蟲存活、附著和變態的影響[J]. 海洋與湖沼, 2020, 51(3): 541-546.
[9] 王濤, 李琪. 腎上腺素對巖牡蠣幼蟲變態的誘導[J]. 水產學報, 2018, 42(11): 1729-1736.
[10] YANG J L, LI S H, LI Y F, et al. Effects of neuroactive compounds, ions and organic solvents on larval metamorphosis of the mussel[J]. Aquaculture, 2013, 396/397/398/399: 106-112.
[11] Galley T H. Induction of metamorphosis and seed attachment in hatchery production of the king scallop(L.) and the blue mussel(L.) [M]. Bangor University (United Kingdom), 2014.
[12] 楊創業, 廖永山, 蔡煒裕, 等. 流沙灣海區不同規格馬氏珠母貝氨基酸組成的研究[J]. 廣東海洋大學學報, 2017, 37(4): 86-91.
[13] ZHANG J B, XIONG X W, DENG Y W, et al. Integrated application of transcriptomics and metabolomics provides insights into the larval metamorphosis of pearl oyster ()[J]. Aquaculture, 2021, 532: 736067.
[14] YU X J, HE W H, GU J D, et al. The effect of chemical cues on settlement of pearl oyster(Dunker) larvae[J]. Aquaculture, 2008, 277(1/2): 83-91.
[15] WANG X B, BAI Y, HUANG B. Effects of chemical cues on larval survival, settlement and metamorphosis of abalone[J]. Chinese Journal of Oceanology and Limnology, 2010, 28(6): 1261-1265.
[16] 李一峰, 楊金龍, 包衛洋, 等. 人工誘導物影響海洋無脊椎動物幼體附著變態的研究[J]. 海洋科學, 2011, 35(8): 102-107.
[17] ZHAO B, ZHANG S, QIAN P Y. Larval settlement of the silver- or goldlip pearl oyster(Jameson) in response to natural biofilms and chemical cues[J]. Aquaculture, 2003, 220(1/2/3/4): 883-901.
[18] KING-JONES K, THUMMEL C S. Nuclear receptors—a perspective from[J]. Nature Reviews Genetics, 2005, 6(4): 311-323.
[19] VOGELER S, GALLOWAY T S, LYONS B P, et al. The nuclear receptor gene family in the Pacific oyster,, contains a novel subfamily group[J]. BMC Genomics, 2014, 15: 369.
[20] KAUR S, JOBLING S, JONES C S, et al. The nuclear receptors ofand: implications for developing new model organisms[J]. PLoS One, 2015, 10(4): e0121259.
[21] LAGUERRE M, VEENSTRA J A. Ecdysone receptor homologs from mollusks, leeches and a polychaete worm[J]. FEBS Letters, 2010, 584(21): 4458-4462.
[22] VOGELER S, BEAN T P, LYONS B P, et al. Dynamics of nuclear receptor gene expression during Pacific oyster development[J]. BMC Developmental Biology, 2016, 16(1): 33.
[23] CHELIKOWSKY D J, KAPAREIKO D, JEFFRESS D, et al. The microalga Pavlova contains an analog for the hormone ecdysone that promotes metamorphosis of larval bay scallops ()[J]. Connect, 2013: 517-561.
[24] HE J, QI J F, HUANG Y Q, et al. Larval settlement and metamorphosis of the invasive biofouler,, in response to ions and neuroactive compounds[J]. Marine Biology Research, 2017, 13(6): 645-658.
[25] MESíAS-GANSBILLER C, BENDIMERAD M E A, ROMáN G, et al. Settlement behavior of black scallop larvae (, L.) in response to GABA, epinephrine and IBMX[J]. Journal of Shellfish Research, 2008, 27(2): 261-264.
[26] YOUNG T, ALFARO A C, ROBERTSON J. Effect of neuroactive compounds on the settlement of mussel () larvae[J]. Aquaculture, 2011, 319(1/2): 277-283.
[27] 高如承, 劉文彪. 幾種化學物質對西施舌幼蟲附著和變態的誘導技術[J]. 水產學報, 2006, 30(5): 597-602.
[28] WAKISAKA N, MIYASAKA N, KOIDE T, et al. An adenosine receptor for olfaction in fish[J]. Current Biology, 2017, 27(10): 1437-1447.e4.
[29] SHAMCHUK A L, BLUNT B J, LYONS D D, et al. Nucleobase-containing compounds evoke behavioural, olfactory, and transcriptional responses in model fishes[J]. FACETS, 2018, 3(1): 79-102.
[30] ZEECK E, HARDER T, BECKMANN M. Uric acid: the sperm-release pheromone of the marine polychaete[J]. Journal of Chemical Ecology, 1998, 24(1): 13-22.
[31] ZEECK E, HARDER T, BECKMANN M. Inosine, L-glutamic acid and L-glutamine as components of a sex pheromone complex of the marine polychaete(Annelida:)[J]. CHEMOECOLOGY, 1998, 8(2): 77-84.
[32] HE J, DAI Q, QI Y X, et al. Aggregation pheromone for an invasive mussel consists of a precise combination of three common purines[J]. iScience, 2019, 19: 691-702.
Effect of Chemical Cues on Settlement of Pearl OysterEyespot Larvae
YANG Chuang-ye1, ZHANG Jia-bin1, ZENG Ye-tao1, LIAO Yong-shan2, DENG Yue-wen1,3, WANG Qing-heng1,3
(1.// 2.// 3.,524088,)
【Objective】The objective of the present study was to research the effects of 5-Hydroxytryptamine (5-HT), ecdysone, γ-aminobutyric acid (GABA), adenine nucleotides, hypoxanthine nucleotides and hypoxanthine on theeyespot larvae settlement of. 【Method】By studying thetime-course and changes on settlement rate of eyespot larvae after5 treating with different concentrations of 5-HT, ecdysone, GABA, adenine nucleotides, hypoxanthine nucleotides and hypoxanthine.【Result】During the 24?96 h, the settlement of eyespot larvae was induced by 5-HT, ecdysone and GABA showed an overall upward trend. During the 24-120 h, the settlement of eyespot larvae induced by adenine nucleotides, hypoxanthine nucleotides and hypoxanthine showed an overall upward trend. After treatment with 10-5mol/L 5-HT and GABA for 24 h, 48 h, 72 h and 96 h, the settlement rate of eyespot larvae was significantly higher than that of the control group (< 0.01). The 10-5mol/L ecdysone treatment for 24 h and 72 h, 10-6mol/L ecdysone treatment for 24 h and 96 h, and 10-7mol/L ecdysone treatment for 24 h, the settlement rate of eyespot larvae was significantly higher than that of the control group (< 0.01). The 10 μmol/L adenosine nucleotides treatment for 24 h, 48 h, 72 h and 120 h, 1 μmol/L adenosine nucleotides treatment for 24 h, 48 h, 72 h, 96 h and 120 h, the settlement rate of eyespot larvae was significantly higher than that of the control group (< 0.01). The10 μmol/L inosine treatment for 48 h and 72 h, 1 μmol/L inosine treatment for 48 h, 72 h, 96 h and 120 h, the eyespot larval settlement rate was significantly higher than that of the control group (< 0.01). 10 μmol/L hypoxanthine treatment for 72 h, 96 h and 120 h, the eyespot larvae settlement rate was significantly higher than the control group (< 0.01).【Conclusion】With an appropriate concentration and treatment time, 5-HT, ecdysone, GABA, adenine nucleotides, hypoxanthine nucleotides and hypoxanthine solutions could induce eyespot larvae settlement and a 10-5mol/L 5-HT treatment for 24 h presented the best induction effect on eyespot larvae settlement rate, which showed 6.3 times higher than that of control group.
; eyespot larvae; settlement; chemical cues
S917.4
A
1673-9159(2022)01-0029-06
10.3969/j.issn.1673-9159.2022.01.005
楊創業,章佳斌,曾業濤,等. 化學物質誘導對馬氏珠母貝眼點幼蟲附著的影響[J]. 廣東海洋大學學報,2022,42(1):29-34.
2021-08-22
國家自然科學基金(32102817);廣東省基礎與應用基礎研究基金項目(2019A1515111026);廣東省農業農村廳項目(2020KJ146和粵財農[2020]4號);廣東省教育廳項目(2019KQNCX043和2020ZDZX1045);大學生創新創業訓練計劃項目(CXXL2020045)
楊創業(1990-),男,博士,講師,研究方向為無脊椎動物增養殖及珍珠培育。E-mail:yangcy@gdou.edu.cn
鄧岳文,教授,研究方向為珍珠貝養殖與育珠。E-mail:dengyw@gdou.edu.cn
(責任編輯:劉嶺)
