化學誘導對2株海洋真菌次級代謝產物及其生物活性的影響
2022-01-19 02:27:06劉亞月薛欣怡黎卓怡馬小翔梁金月胡雪瓊
廣東海洋大學學報 2022年1期
劉亞月,薛欣怡,黎卓怡,馬小翔,梁金月,胡雪瓊,張 翼,2,3
化學誘導對2株海洋真菌次級代謝產物及其生物活性的影響
劉亞月1,3,薛欣怡1,黎卓怡1,馬小翔1,梁金月1,胡雪瓊1,3,張 翼1,2,3
(1. 廣東海洋大學食品科技學院//廣東省水產品加工與安全重點實驗室//廣東省海洋生物制品工程實驗室//廣東省海洋食品工程技術研究中心//水產品深加工廣東普通高等學校重點實驗室//湛江市腦健康海洋藥物與營養品重點實驗室//廣東海洋大學海洋藥物研究所,廣東 湛江 524088;2. 廣東海洋大學深圳研究院海洋醫藥研發中心, 廣東 深圳 518120;3. 海洋食品精深加工關鍵技術省部共建協同創新中心, 大連工業大學, 遼寧 大連 116034)
【目的】以2株海洋真菌為研究對象,探究不同種類及濃度的化學表觀遺傳修飾劑對其次級代謝產物化學多樣性和生物活性的影響。【方法】基于化學表觀遺傳修飾策略,采用兩種不同的化學表觀遺傳修飾劑5-氮雜胞嘧啶核苷(5-azaC)和丁酸鈉,分別對sp. 019和ZN4-5-4兩株海洋來源真菌進行表觀遺傳修飾。通過觀察真菌在培養基中的生長狀態變化,并根據薄層層析(TLC)指紋圖譜、高效液相色譜(HPLC)圖譜、乙酰膽堿酯酶(AChE)抑制、1,1-二苯基-2-三硝基苯肼(1,1-Diphenyl-2-picryl-hydrazyl, DPPH)自由基清除和鹵蟲致死活性篩選模型等技術手段,從中篩選出代謝產物豐富、AChE抑制活性與抗氧化活性產物豐富且毒性小的發酵培養條件。【結果】添加1 mmol/mL的丁酸鈉誘導劑可使菌株019和ZN4-5-4的次生代謝產物的種類增加,且同時能提高該菌株次級代謝產物的AChE的抑制活性和DPPH自由基清除活性。因此,確定菌株019和ZN4-5-4的最優發酵條件均是添加濃度為1 mmol/mL丁酸鈉的PDB培養基。【結論】化學表觀遺傳修飾策略指導下,添加誘導劑5-azaC或者丁酸鈉對2株海洋真菌次生代謝產物的產生有影響。
海洋真菌;化學誘導;乙酰膽堿酯酶抑制活性;抗氧化;鹵蟲致死活性
海洋真菌作為海洋生物類群的重要組成部分,其次生代謝產物具有多樣的結構類型和顯著的藥理活性,已成為海洋天然藥物研究重要生物資源和研究熱點[1]。海洋真菌次級代謝產物分離得到的新化合物多具有抗病毒、抗腫瘤、抗菌活性和抗心腦血管疾病、抗氧化、酶抑制劑等顯著的藥理活性[2]。從中分離得到二蓖麻酚單醚化合物,其中化合物versiorcinols A顯示中等強度的抑菌活性[3]。從海洋藻類內生真菌EH 399中分離獲得六個化合物均對大腸桿菌、黃褐微球菌和哈維弧菌顯示一定的抗菌活性,其中cladosporol H對A549、Huh7和LM3細胞系表現出明顯的細胞毒性,cladosporol C則對H446細胞系表現出細胞毒性[4]。但是現有的研究只涉及海洋天然資源中的一小部分。海洋中大量的微生物,尤其是內生微生物中活性先導化合物的篩選工作尚少,具有巨大的開發潛力。化學表觀遺傳修飾是指通過添加一些小分子的化學表觀遺傳修飾劑,特異性地改變化學表觀遺傳修飾酶的活性,激活某些未表達次生代謝產物的沉默基因簇,誘導真菌產生不同結構骨架或生物活性更豐富的次級代謝產物[5-6]。研究發現,真菌中存在大量編碼化合物的基因,目前從中分離得到的化合物數量遠少于真菌擁有的這些基因所能編碼的化合物數量,這證明其中很多基因都處于沉默狀態。激活沉默基因,可以使微生物表達出新的次級代謝產物,增加微生物的化學多樣性,從而有可能產生數量更多、結構類型更豐富的代謝產物,對進一步研究海洋微生物次級代謝產物具有重要意義[7-9]。化學表觀遺傳修飾是目前激活沉默基因表達的有效方法之一,具有成本低、效率高和易操作等優點。既可應用于模式真菌的研究,也可以用于修飾遺傳背景不清楚的真菌,不受實驗條件限制,能夠在大部分實驗室中完成[6]。
本研究中以2株海洋真菌為研究對象,探究不同種類及濃度的化學表觀遺傳修飾劑對其次級代謝產物化學多樣性和生物活性的影響。分別選用丁酸鈉和5-azaC作為誘導劑對兩株海洋真菌進行化學誘導調控,選出各菌株次級代謝產物豐富、抗AD相關活性明顯且毒性小的最優發酵條件,以期為后續深入挖掘2株真菌的活性次級代謝產物奠定基礎。
1 材料與方法
1.1 材料
1.1.1 菌株來源 海洋真菌sp. 019采集于湛江紅樹林自然保護區的紅樹植物楊葉肖槿(),經ITS rDNA全序列分析確定為青霉菌(Genbank登錄號:MN826202);海洋真菌ZN4-5-4 (簡稱:ZN)則是采集于瓊州海峽的沉積泥,經ITS rDNA全序列分析確定為曲霉菌(Genbank登錄號:MN826203)。目前兩株真菌菌株均保存于廣東海洋大學海洋藥物研究所。
1.1.2 培養基 PDB培養基:葡萄糖20 g,蛋白胨5 g,海鹽3 g,加入純水至1 L,于1×105Pa滅菌30 min。
1.1.3 主要試劑與儀器 主要試劑:誘導劑5-azaC和丁酸鈉分別配成5個濃度梯度的貯液(250 mmol/L、25 mmol/L、250 μmol/L、25 μmol/L、2.5 μmol/L)。其中5-azaC用DMSO溶解,丁酸鈉用無菌水溶解,經0.22 μm濾膜除菌后待用。AChE (C3389)、碘代硫代乙酰膽堿 (Acetylthiocholine iodide, ATCI,DA0048)、牛血清蛋白 (Bovine serum albumin, BSA,A1933)、1, 1-二苯基-2-三硝基苯肼 (DPPH,D9132),均購自Sigma-Aldrich;5, 5′-二硫代二硝基苯甲酸 (Dithiobisnitrobenzoic acid, DTNB,D8130),購自Ruibio公司。其余試劑均為國產分析純試劑。
主要儀器:GF254硅膠高效薄層層析板購自德國默克公司。鹽水蝦幼蟲卵購自中國愛家水族館。1260 Infinity Ⅱ高效液相色譜儀,美國Agilent公司;Epoch2酶標儀,美國BioTek公司;R-300旋轉蒸發儀,瑞士BüChi公司;多功能紫外透射儀WFH-201B,上海精科實業有限公司。
1.2 方法
1.2.1 菌株誘導前種子的制備 將待誘導的兩株海洋真菌放進28 ℃、濕度為80%的霉菌培養箱中活化過夜;在無菌條件下,將菌種接種到預先裝有15 mL滅菌馬鈴薯葡萄糖固體培養基的平板中;于28 ℃恒溫培養箱中培養3~4 d,待菌落孢子生長茂盛備用。
1.2.2 菌株的化學誘導 將上述培養好的菌株用0.85%的無菌生理鹽水洗滌、搖勻,形成孢子懸液。采用12孔板,每孔加入2 mL PDB培養基和200 μL孢子懸液,28 ℃培養3 d后,分別加入化學誘導劑,并設置5個誘導劑終濃度梯度 (10 mmol/L、1 mmol/L、100 μmol/L、10 μmol/L和1 μmol/L),每組設置2個平行;將12孔板置于28 ℃、濕度為80%的霉菌培養箱中靜置培養14 d,觀察菌株生長狀態的變化并拍照記錄。
1.2.3 菌株次生代謝產物的提取 從12孔板中取出菌絲體和發酵液,3 000 r/min離心、濃縮干燥4 h,加入200 μL甲醇,功率500 W超聲提取30 min,10 000 r/min離心30 min,取出上清液備用。減壓濃縮、干燥、稱量,得到粗提取物的質量(mg)。
1.2.4 菌株次生代謝產物的TLC和HPLC圖像分析 將上述獲得的各粗提物用甲醇統一配制成10 mg/mL的濃縮液,分別采用GF254TLC硅膠薄層層析板和HPLC對各粗提物的化學多樣性進行分析。選擇氯仿(CHCl3):甲醇(MO)= 7∶1 (∶)二元溶劑體系作為TLC分析的展開劑,分別記錄254和365 nm波長下代謝產物的紫外圖像。HPLC分析中,進樣量為10 μL,洗脫條件為:0 ~ 15 min,50% ~ 100% 的甲醇-水;15 ~ 25 min,100%甲醇; 25 ~ 27 min,100% ~ 50% 的甲醇-水;27 ~ 30 min,50%的甲醇-水,流速0.6 mL/min。DAD檢測器信號采集波長為190 ~ 400 nm,檢測波長為254 nm。
1.2.5 菌株次生代謝產物的AChE抑制活性測定
通過優化的Ellman’s比色法[10]測定粗提物對AChE的體外抑制活性。
初篩:往96孔板中每孔加100 μL用甲醇溶解的質量濃度為5 mg/mL的樣品,等待樣品中有機溶劑揮干。再依次往96孔板中每孔加入1 μL的DMSO,49 μL PBS、10 μL 0.2 U/mL AChE和20 μL DTNB,將96孔板置于37 ℃培養箱保溫10 min。然后往每個孔加入20 μL ATCh,繼續于37 ℃培養箱孵育。20 min后,通過酶標儀測定405nm 處的光密度值405nm,鹽酸化多奈哌齊作為陽性對照[11]。通過下述公式計算化合物對AChE的抑制率。
抑制率=[(control?blank)?(sample?sample blank)]/(control?blank)。 (1)
其中sample blank為加入樣品和BSA,但不加入AChE的光密度,sample為加入樣品和AChE的光密度,control為加入AChE但不加入樣品的光密度,blank為加入BSA但不加入樣品和AChE的光密度。
復篩:經上述初篩后,選擇AChE抑制率大于50%的樣品進行活性復篩實驗。采用倍半梯度稀釋法對樣品進行稀釋后,按照上述方法測定各粗提物的IC50值,IC50值是化合物對AChE抑制為50%所對應的濃度。將抑制率和濃度對數 (lnC)導入Origin 9.1軟件。通過三次多項式回歸方程擬合得到濃度對數曲線,并計算得到ln(IC50)。最后使用Microsoft Excel計算得到IC50。
1.2.6 菌株次生代謝產物的DPPH自由基清除活性測定 使用Sharma和Bhat[12]描述的DPPH自由基清除法,在96孔板中測定粗提物的抗氧化能力。
初篩:總反應體系為100 μL,sample為含有50 μL的0.16 mmol/L DPPH和50 μL溶解濃度為5 mg/mL粗提物的DMSO。與此同時,50 μL 0.16 mmol/L DPPH和50 μL DMSO用作對照(control),50 μL MeOH和50 μL溶解不同濃度化合物的DMSO作為樣品本底(sample blank),50 μL MeOH和50L DMSO 作為對照(blank),將96孔板放置在暗處30 min。通過酶標儀測定λ517nm處的光密度517nm。維生素C作為陽性對照[13]。通過下述公式計算化合物的清除率。
清除率= [(control?blank)?(sample?sample blank)]/(control?blank)。 (2)
復篩:經上述初篩后,選擇DPPH自由基清除率大于50%的樣品進行活性復篩實驗。采用倍半梯度稀釋法對樣品進行稀釋后,按照上述方法測定各粗提物的EC50值,EC50值是化合物對DPPH自由基清除率為50%所對應的濃度。將清除率和濃度對數 (lnC)導入Origin 9.1軟件。通過三次多項式回歸方程擬合得到濃度對數曲線,并計算得到ln (EC50)。最后使用Microsoft Excel計算得到EC50。
1.2.7 菌株次生代謝產物的鹵蟲致死活性測定[14]采用96孔板法,使用鹽水蝦幼蟲測試化合物的致死能力。每孔加199 μL含鹵蟲幼體的溶液(每孔約含20只鹵蟲),制成測試培養板。將粗提物用DMSO配制成10 mg/mL的溶液,加入1 μL到每個培養板孔中,每個粗提物樣品設置成50 μg/mL終濃度,設3個平行樣。在28 ℃的培養箱中保持光照培養24 h后,記錄每個孔中死亡幼蟲的數量和總數。同時,用DMSO和秋水仙堿[15]分別作為陰性和陽性對照。鹵蟲致死活性采用鹵蟲致死率表示:
鹵蟲致死率= [(加藥組致死率-對照組致死率)/對照組存活率]。 (3)
2 結果與分析
2.1 菌株誘導前后表觀形態對比
采用12孔板對2株海洋真菌進行微量培養,結果如圖1所示。添加誘導劑5-azaC進行誘導前后,菌株019和ZN的表觀生長形態相比空白組均發生了顯著變化。如圖1(B1)所示,添加誘導劑5-azaC進行誘導前后,菌株019的菌絲體由黑褐色變成棕色,菌液變為黃色。并且隨著誘導劑濃度增加,菌液顏色逐步加深。菌株ZN的菌絲體則由棕色變成白色,菌液變為亮黃色(圖1(C1))。由此可以推斷,誘導劑5-azaC的加入對菌株019和ZN的次生代謝產物可能產生了影響。而添加丁酸鈉進行誘導前后,2株菌株的菌絲體顏色雖未表現出明顯變化,但菌絲體厚度和菌液顏色深淺隨著加入誘導劑濃度不同也表現出一定變化。

B1、B2、C1和C2分別代表菌株019和ZN在添加化學誘導劑5-azaC和丁酸鈉的PDB培養基中培養14 d生長形態;依次是B1為019添加5-azaC,B2為019添加丁酸鈉,C1為ZN添加 5-azaC,C2為ZN添加丁酸鈉;12孔板中每個孔添加的誘導劑濃度如圖A所示
2.2 菌株誘導前后次生代謝產物的產量分析對比
微量培養14 d后,用甲醇對2株菌株的次生代謝產物進行提取結果如圖2所示。結果表明,添加誘導劑誘導前后,2株菌株的次生代謝產物產量均發生了相應變化。以菌株019為例,在添加誘導劑進行誘導后,菌株次生代謝產物產量明顯增加;尤其是在添加誘導劑5-azaC濃度為10 μmol/L和1 μmol/L的PDB培養基條件下的產量增加顯著 (0.000 1),大于同等濃度下添加誘導劑丁酸鈉誘導后的產量。菌株ZN則是隨誘導劑加入,代謝產物質量逐漸增加,當添加濃度為10 μmol/L 5-azaC的PDB培養基條件下的產量增加最明顯。但是隨著添加誘導劑5-azaC濃度進一步增大,次級代謝產物的產量卻明顯下降。可推測高濃度的誘導劑5-azaC的加入對菌株ZN的代謝產生了抑制作用。
2.3 菌株誘導前后次級代謝產物的TLC和HPLC圖像分析對比
利用TLC和HPLC對化學誘導前后不同菌株的次生代謝產物進行分析可以發現:對菌株019而言,觀察其TLC圖像可知,5-azaC誘導組 (1 μmol/L除外)相比對照組的代謝產物在波長254 nm和365 nm下吸收斑點明顯不同(圖3(B1));丁酸鈉誘導組相比對照組的代謝產物沒有明顯區別,但是在添加誘導劑濃度為100 μmol/L時,其主產物吸收斑點的強度明顯增強,這意味著在該培養條件下,菌株019代謝某些化合物的能力增強(圖3(B2))。進一步觀察其HPLC圖像可知,在254 nm波長下添加不同濃度的5-azaC誘導組相比對照組的吸收峰明顯增多(圖4(B1));而低濃度丁酸鈉誘導組相比對照組的代謝產物主吸收峰沒有明顯區別,但是隨著添加誘導劑濃度增大,在濃度為10 mmol/L和1 mmol/L時,其吸收峰的個數明顯增多,這意味著誘導劑的加入明顯改變了菌株019的次生代謝產物,菌株的代謝產物更豐富(圖4(B2))。而對菌株ZN而言,觀察其TLC圖像可知,高濃度的5-azaC誘導組和丁酸鈉誘導組相比對照組的代謝產物在波長254 nm和365 nm下吸收斑點都有所增加(圖3(C1、C2))。進一步分析其HPLC圖像可得出相同結論,在添加誘導劑濃度為1 μmol/L和10 μmol/L時,不管是添加5-azaC還是丁酸鈉,菌株代謝產物均沒有明顯變化。但隨著添加誘導劑濃度進一步增大,菌株的吸收峰明顯增多,部分代謝產物的峰吸收強度也有一定增強(圖4(C1、C2))。這表明對菌株ZN而言,高濃度誘導劑的加入更能刺激菌株次生代謝產物的變化。由此推測,在以上特定誘導條件下,菌株可能產生了新的代謝產物。

樣品的編號為xxx-y-z,xxx代表菌株編號簡稱;y代表添加誘導劑的類型,1代表添加誘導劑5-azaC,2代表添加誘導劑丁酸鈉,z代表添加誘導劑濃度,1-6分別表示10 mmol/L、1 mmol/L、100 μmol/L、10 μmol/L、1 μmol/L、空白; “****”表示與空白組比較,P < 0.0001
2.4 菌株誘導前后次級代謝產物生物活性分析對比
各菌株粗提物的AChE抑制活性、DPPH自由基清除活性和鹵蟲致死活性如圖5。對菌株019而言,無論是添加誘導劑5-azaC或丁酸鈉誘導后,菌株次生代謝產物的AChE抑制活性、DPPH自由基清除活性和鹵蟲致死活性均明顯增強。尤其是添加誘導劑5-azaC誘導后,其代謝產物的AChE抑制活性均大于40%,其中添加濃度為1 mmol/L的5-azaC誘導組菌株次生代謝產物的AChE抑制率高達 96.4%,而空白組不顯示抑制活性。添加誘導劑丁酸鈉(1 mmol/L)誘導后,其菌株次生代謝產物的AChE抑制率為95.6%,但其鹵蟲致死率也高達63.6%。對菌株ZN而言,添加誘導劑5-azaC誘導后,菌株次生代謝產物的AChE抑制活性相比對照組明顯減弱甚至消失;同時菌株次級代謝產物的鹵蟲致死活性有所增強(濃度為10 μmol/L時,抑制率為60.0%);而添加誘導劑丁酸鈉(10 mmol/L、1 mmol/L和100 μmol/L)誘導后,菌株次級代謝產物的AChE抑制活性明顯增強。
經初篩后,抑制率或者清除率大于50%的樣品活性復篩結果如表1所示。菌株019添加5-azaC (1 mmol/L)的誘導組其次生代謝產物顯示較強的AChE抑制活性(IC50為31.9 μg/mL)。但在添加誘導劑丁酸鈉(1 mmol/L)進行誘導后其次生代謝產物同時顯示較強的AChE抑制活性(IC50值為32.2 μg/mL)和清除活性(EC50為80.6 μg/mL)。因此,選擇具有雙重活性的丁酸鈉誘導組(1 mmol/L)作為最優發酵條件。菌株ZN4-5-4在添加誘導劑丁酸鈉(1 mmol/L和100 μmol/L)的PDB培養基條件下的次級代謝產物也具有顯著的AChE抑制活性,IC50值均為24.9 μg/mL。結合兩個條件下菌株次級代謝產物的產量因素,選擇產量更多的丁酸鈉誘導組(1 mmol/L)作為最優發酵條件。

254 nm和365 nm表示測定時選用的紫外波長條件;10 mmol/L、1 mmol/L、100 μmol/L、10 μmol/L、1 μmol/L表示添加誘導劑的濃度;B1、B2、C1和C2分別代表菌株019和ZN添加化學誘導劑5-azaC和丁酸鈉的次級代謝產物在254 nm波長下的TLC指紋圖譜;B3、B4、C3和C4分別代表菌株019和ZN添加化學誘導劑5-azaC和丁酸鈉的次級代謝產物在365 nm波長下的TLC指紋圖譜;依次是B1、B3為019添加5-azaC,B2、B4為019添加丁酸鈉,C1、C3為ZN添加 5-azaC,C2、C4為ZN添加丁酸鈉。

圖4 誘導前后菌株019和ZN發酵產物的HPLC圖譜

圖5 誘導前后菌株粗提物的AChE抑制活性、DPPH自由基清除活性和鹵蟲致死活性生物活性
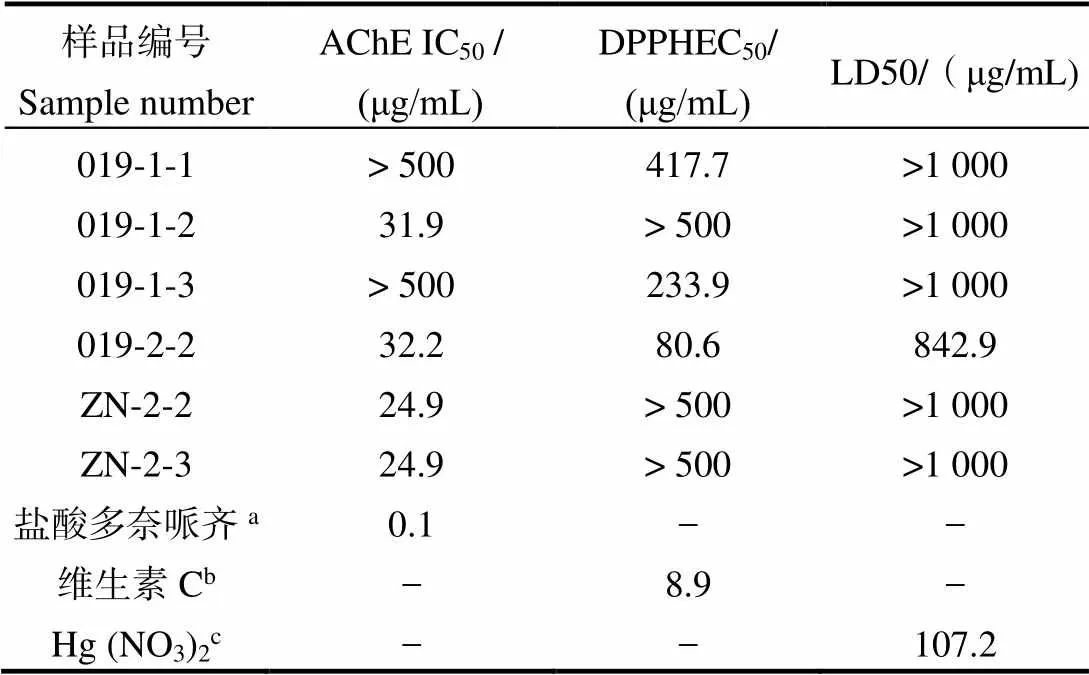
表1 部分樣品的IC50、EC50和LD50活性值
注:a, b, c表示陽性對照。“-”表示沒有測試。
Note: a/b/c means positive control,“-”means not test.
3 討論
大多數微生物在傳統單一的條件下培養只能表現出有限的生物活性。而將實驗室條件下生長的微生物的代謝產物種類與自然環境下生長微生物的代謝產物種類進行對照,發現許多代謝產物的表達被抑制了[16]。因此,如何采用科學合理的方法來削弱環境對于微生物內部表達基因的抑制作用是獲得新型次生代謝產物表達的核心問題。大量科學研究表明,化學表觀遺傳修飾策略能夠很好達到發掘菌株潛能、獲得新型代謝產物的目的[17-19]。如王曉茹等[20]在真菌的培養基中加入濃度為 50 μmol/L的DNA甲基化酶抑制劑 5-azaC,結果發現實驗組與對照組的菌株在生長過程中各自的菌絲體形貌上出現了很大差別,最后通過對其次生代謝產物采用高效液相色譜分析,發現在激活后該菌與對照組相比產生了 10 個新色譜峰。通過對實驗組真菌代謝產物進行進一步分離純化,得到了 2 個新的萜類化合物atlantinone A和 atlantinone B。楊靜明等報道[21]采用組蛋白去乙酰化酶抑制劑丁酸鈉處理一株珊瑚來源的土曲霉C23-3,可以顯著提高其丁內酯類代謝產物的差異性。在本研究中,同時選用去甲基化酶抑制劑5-azaC和組蛋白乙酰化酶抑制劑丁酸鈉兩種誘導劑進行研究,并設立5個濃度梯度(10 mmol/L、1 mmol/L、100 μmol/L、10 μmol/L和1 μmol/L)對sp. 019和ZN4-5-4兩株海洋來源真菌進行表觀遺傳修飾,發現不同種類及濃度的化學表觀遺傳修飾劑對不同海洋來源真菌次生代謝產物的產生和生物活性影響不同。
在菌株次級代謝產物產量和化學多樣性方面,添加誘導劑能在一定程度上增加菌株次生代謝產物的產量。如本研究中,添加濃度為10 μmol/L和1 μmol/L的誘導劑5-azaC對菌株019和ZN次生代謝產物的產量具有顯著的增加作用(增長率> 46.7%)。同時結合菌株的TLC和HPLC圖像可知,菌株的次級代謝產物的種類和個數也隨著誘導劑的加入逐漸增多。如在添加濃度為10 mmol/L、1 mmol/L和100 μmol/L的誘導劑的情況下,菌株019和ZN的次生代謝產物種類明顯增多。通過這些現象推測,誘導劑5-azaC和丁酸鈉的加入可在一定程度上激活或者增強菌株的某些沉默基因,使得菌株的次生代謝產物的種類增加或者產量增加。
在菌株次級代謝產物生物活性方面,添加不同誘導劑對菌株次級代謝產物的生物活性影響不一。如本研究中,添加5-azaC對菌株019中抗AChE抑制和鹵蟲致死活性成分的產生具有顯著激活作用,但是對菌株ZN中抗AChE活性成分的產生具有顯著抑制作用。而添加丁酸鈉可明顯提高菌株019和ZN次生代謝產物的AChE抑制活性。同時發現,這種誘導激活作用并不具備濃度依賴性,如在用5-azaC激活的菌株019次生代謝產物實驗中,在低于濃度為1 mmol/L時,菌株的抗AChE活性隨著丁酸鈉濃度增大而增強,當濃度超過1 mmol/L時,菌株019次生代謝產物的抗AChE活性隨著5-azaC濃度升高而減小。可能的原因有:一是隨著誘導劑5-azaC的加入,菌株019細胞內編碼AChE抑制活性組分的沉默基因被激活,并且隨著濃度增加,這種激活作用越明顯,從而導致大量抗AChE活性的代謝產物被合成了出來,使得粗提物的AChE抑制活性增強。但當濃度增大到一定的時候,由于對主產物的基因激活作用達到飽和,且可能有其他新的基因被激活,導致新的代謝產物被合成,最終導致次生代謝產物的抗AChE活性降低。二有可能是由于DNA甲基化酶抑制劑被過多加入,從而導致DNA甲基化酶被過多抑制,導致生物體內代謝平衡紊亂,從而影響了真菌正常生長代謝過程,最終導致菌株019代謝產物的抗AChE活性的降低。由此可以得出,不同表觀遺傳修飾劑對于不同真菌的抗AChE活性代謝產物的激活是有濃度限制的,這個濃度可認為是表觀遺傳修飾劑作用的最佳濃度。
4 結論
本研究表明,不同種類及濃度的化學表觀遺傳修飾劑對不同海洋來源真菌次生代謝產物的產生和生物活性影響不同。綜合菌株次級代謝產物的產量、種類、AChE抑制活性、DPPH自由基清除活性和鹵蟲致死活性結果分析可知:添加1 mmol/mL的5-azaC和丁酸鈉誘導劑均可使菌株019的次生代謝產物的種類增加;且同時能提高該菌株次級代謝產物的AChE的抑制活性和DPPH自由基清除活性。但添加1 mmol/mL的5-azaC會增大次級代謝產物的鹵蟲致死率。而對菌株ZN4-5-4而言,添加1 mmol/mL和100 μmol/L的丁酸鈉誘導劑均可使該菌株次生代謝產物的種類增加,且同時能提高次級代謝產物的AChE抑制活性和DPPH自由基清除活性。但添加1 mmol/mL的丁酸鈉可使菌株次級代謝產物產量增大程度更多。綜上所述,由此確定了菌株019和ZN4-5-4的最優發酵條件均為添加了濃度為1 mmol/mL丁酸鈉的PDB培養基。
[1] RATEB M E, EBEL R. Secondary metabolites of fungifrommarine habitats[J]. Natural Product Reports, 2011, 28(2): 290-344.
[2] CARROLL A R, COPP B R, DAVIS R A, et al. Marine natural products[J]. Natural Product Reports, 2020, 37(2): 175-223.
[3] GU B B, TANG J, WANG S P, et al. Structure, absolute configuration, and variable-temperature 1H-NMR study of (±)-versiorcinols A-C, three racemates of diorcinol monoethers from the sponge-associated fungus16F-11[J]. RSC Advances, 2017, 7(79): 50254-50263.
[4] LI H, LI X, MANDI A, et al. Characterization of cladosporols from the marine algal-derived endophytic fungusEN-399 and configurational revision of the previously reported cladosporol derivatives [J]. Journal of Organic Chemistry, 2017, 82(19): 9946-9954.
[5] TAKAHASHI J A, TELES A P C, ALMEIDA PINTO BRACARENSE A, et al. Classical and epigenetic approaches to metabolite diversification in filamentous fungi[J]. Phytochemistry Reviews, 2013, 12(4): 773-789.
[6] 張偉, 陳敏, 邵長倫, 等. 化學表觀遺傳修飾方法在真菌次級代謝產物研究中的應用[J]. 中國海洋藥物, 2014, 33(5): 83-91.
[7] 孫坤來. 表觀遺傳修飾增加兩株滸苔真菌的化學多樣性研究[D]. 青島: 中國海洋大學, 2014.
[8] 李德寶, 張雪晴, 張秀麗, 等. 1株花柳珊瑚來源真菌產cytochalasin B發酵優化及其化學表觀遺傳修飾研究[J]. 中國海洋藥物, 2016, 35(5): 14-20.
[9] 丁佩. 化學表觀遺傳修飾劑激活海洋真菌sp.AA007次級代謝產物的研究[D]. 杭州: 浙江工商大學, 2017.
[10] ELLMAN G L, COURTNEY K D, ANDRES V jr, et al. A new and rapid colorimetric determination of acetylcholinesterase activity[J]. Biochemical Pharmacology, 1961, 7(2): 88-95.
[11] PALEACU D, MAZEH D, MIRECKI I, et al. Donepezil for the treatment of behavioral symptoms in patients with Alzheimer's disease[J]. Clinical Neuropharmacology, 2002, 25(6): 313-317.
[12] SHARMA O P, BHAT T K. DPPH antioxidant assay revisited[J]. Food Chemistry, 2009, 113(4): 1202-1205.
[13] PADAYATTY S J, KATZ A, WANG Y H, et al. Vitamin C as an antioxidant: evaluation of its role in disease prevention[J]. Journal of the American College of Nutrition, 2003, 22(1): 18-35.
[14] SOLIS P N, WRIGHT C W, ANDERSON M M, et al. A microwell cytotoxicity assay using(brine shrimp)[J]. Planta Medica, 1993, 59(3): 250-252.
[15] AN C Y, LI X M, LI C S, et al. Prenylated indolediketopiperazine peroxides and related homologues from the marine sediment-derived fungusSD-273[J]. Mar Drugs, 2014, 12(2): 746-756.
[16] 楊康敏, 高向東, 顧覺奮. 激活沉默基因簇發掘微生物次級代謝產物的研究進展[J]. 中國醫藥生物技術, 2015, 10(1): 77-80.
[17] HENRIKSON J C, HOOVER A R, JOYNER P M, et al. A chemical epigenetics approach for engineering thebiosynthesis of a cryptic natural product from[J]. Organic & Biomolecular Chemistry, 2009, 7(3): 435-438.
[18] 張偉. 化學表觀遺傳修飾法研究海葵來源月狀旋孢腔菌代謝產物多樣性初探[D]. 青島: 中國海洋大學, 2014.
[19] 孫鑒昕. 表觀遺傳修飾劑對三株刺松藻內生真菌次級代謝產物生物活性的影響[D]. 青島: 青島科技大學, 2016.
[20] WANG X R, SENA FILHO J G, HOOVER A R, et al. Chemical epigenetics alters the secondary metabolite composition of guttate excreted by an Atlantic-forest-soil-derived[J]. Journal of Natural Products, 2010, 73(5): 942-948.
[21] 楊靜明, 楊文聰, 劉亞月, 等. 化學誘導對一株海洋來源土曲霉C23-3次生代謝產物及其生物活性的影響[J]. 微生物學通報, 2019, 46(3): 441-452.
Influence of Chemical Induction on the Secondary Metabolites and Biological Activities of Two Marine Fungi
LIU Ya-yue1,3, XUE Xin-yi1, LI Zhuo-yi1, MA Xiao-xiang1, LIANG Jin-yue1, HU Xue-qiong1,3, ZHANG Yi1,2,3
(1.,////////////,,524088,; 2.,,518120,; 3.,,116034,)
【Objective】Based on two marine-derived fungi, this study explore the effects of different types and concentrations of chemical epigenetic modifiers on the secondary metabolites and biological activities.【Method】Based on the chemical epigenetic modification strategy, two strains of marine-derived fungi, i.e.sp. 019 andZN4-5-4, were epigenetically modified by two different chemical epigenetic modifiers, i.e. 5-azacytidine (5-azaC) and sodium butyrate. The morphological variations of the mycelia were observed. Then, based on the thin layer chromatography (TLC) fingerprints, high performance liquid chromatography, acetylcholinesterase (AChE) inhibitory, 1,1-Diphenyl-2-picryl-hydrazyl (DPPH) free radical scavenging, and artemia lethality activities screening models, the fermentation conditions with rich metabolites, low toxicity and rich anti-acetylcholinesterase and antioxidant products were initially screened out.【Result】The results showed that different concentrations (i.e. 10 mmol/L, 1 mmol/L, 100 μmol/L, 10 μmol/L, 1 μmol/L) of 5-azaC and sodium butyrate had significant improvementeffects on the production and biological activity of marine fungal secondary metabolites.【Conclusion】The optimal fermentation conditions for strain 019 and ZN4-5-4 were PDB medium plus 1 mmol/mL sodium butyrate. Under the chemical epigenetic modification strategy, the addition of inducer 5-azaC or sodium butyrate exerts some effects on the production of secondary metabolites of three marine fungi.
marine fungi; chemical induction; acetylcholinesterase inhibitory activity; antioxidant; artemia lethal activity
Q936
A
1673-9159(2022)01-0050-09
10.3969/j.issn.1673-9159.2022.01.008
劉亞月,薛欣怡,黎卓怡,等. 化學誘導對2株海洋真菌次級代謝產物及其生物活性的影響[J]. 廣東海洋大學學報,2022,42(1):50-58.
2021-09-09
國家自然科學基金(21807015);廣東海洋大學博士科研啟動項目(R18008);廣東省揚帆計劃引進緊缺拔尖人才項目(201433009);廣東省自然科學基金(2018A030307046);深圳市科創委基礎研究自由探索項目(JCYJ20170306165013264);深圳市科創委基礎研究面上項目(JCYJ20190813105005619);深圳市大鵬新區產業發展資金(KY20180203 和PT201901-05);深圳市大鵬新區科技研發項目(KJYF202001-07)
劉亞月(1989-),女,博士,講師,主要研究方向為海洋微生物活性物質。E-mail: yayue_liu@163.com
張翼,男,教授,博士,研究方向為海洋生物化學。E-mail: hubeizhangyi@163.com
(責任編輯:劉嶺)
