黃果厚殼桂人工林幼樹群體生存狀態(tài)及譜分析
2022-01-21 07:39:10黃松殿向宇繁陳碧珍鄧祥勝李明蔓
廣西林業(yè)科學(xué) 2021年6期
黃松殿,向宇繁,陳碧珍,鄧祥勝,何 鵬,李明蔓,程 飛,田 湘
(1.廣西壯族自治區(qū)南寧樹木園,廣西南寧 530032;2.廣西大學(xué) 林學(xué)院,廣西南寧 530004)
黃果厚殼桂(Cryptocarya concinna)是樟科(Lau?raceae)厚殼桂屬常綠樹種,主要分布于中國(guó)的廣東、廣西、江西和臺(tái)灣等地,是南亞熱帶季風(fēng)性氣候典型的植被類群之一。黃果厚殼桂的木材紋理美觀、木材顏色鮮明、干形飽滿通直、材質(zhì)堅(jiān)硬且韌且分枝少[1-2],可作為建筑材料和家具原料,具有很高的栽種和研究?jī)r(jià)值。
由于分布區(qū)域的限制,目前對(duì)黃果厚殼桂的研究主要集中在廣東和廣西種群。張池等[3]采用He?gyi 指數(shù)模型分析廣東省自然保護(hù)區(qū)黃果厚殼桂林分的種內(nèi)競(jìng)爭(zhēng),發(fā)現(xiàn)黃果厚殼桂種群的競(jìng)爭(zhēng)強(qiáng)度與林木徑級(jí)、林木間距離有關(guān)。李林等[4]對(duì)黃果厚殼桂種群的死亡動(dòng)態(tài)格局進(jìn)行監(jiān)測(cè),發(fā)現(xiàn)死亡數(shù)量明顯大于個(gè)體新增數(shù)量,需嚴(yán)防重度干擾對(duì)于該種群的影響。張?jiān)伱返萚5]用成對(duì)雙樣本均值分析的方法對(duì)錐栗(Castanea henryi)-荷木(Schima superba)-黃果厚殼桂群落進(jìn)行對(duì)比,了解該群落在時(shí)間上的變化。彭少磷等[6]發(fā)現(xiàn)黃果厚殼桂是我國(guó)季風(fēng)性氣候地區(qū)常綠闊葉林的優(yōu)勢(shì)種群,也是部分南方地區(qū)地帶性林分的建群種。這些研究均以天然林為研究對(duì)象,對(duì)黃果厚殼桂人工林的更新特征仍需進(jìn)一步了解。
種群數(shù)量變化的規(guī)律和動(dòng)態(tài)模型需要對(duì)該物種種群進(jìn)行統(tǒng)計(jì),主要體現(xiàn)方式是生命表。編制靜態(tài)生命表要對(duì)物種種群年齡的分布頻率在一段特定時(shí)間內(nèi)取樣,將數(shù)據(jù)進(jìn)行統(tǒng)計(jì)并描述。靜態(tài)生命表可反映種群出生率和死亡率隨年齡變化的規(guī)律。從靜態(tài)生命表里可解讀出種群結(jié)構(gòu),以生命表為基礎(chǔ)可對(duì)瀕危植物采取措施,改變對(duì)目標(biāo)種群的管理策略[7-9]。
譜分析可將一系列的數(shù)據(jù)從時(shí)域轉(zhuǎn)換到頻域進(jìn)行分析,有效地研究這一系列數(shù)據(jù)是否存在周期性,從而清楚地觀察并了解這段數(shù)據(jù)的動(dòng)態(tài)特征。所有的植被都存在波動(dòng)性,譜分析可揭示種群數(shù)量變動(dòng)的周期性波動(dòng)。通過(guò)譜分析來(lái)觀察種群數(shù)量的動(dòng)態(tài)變化,對(duì)瀕危植被的保護(hù)有重要意義,還可為種群的群落演替、穩(wěn)定性評(píng)價(jià)、物種多樣性保護(hù)和開發(fā)利用提供理論依據(jù)。
本研究以廣西良鳳江國(guó)家森林公園內(nèi)的黃果厚殼桂人工林天然更新的幼樹群體為研究對(duì)象,統(tǒng)計(jì)幼樹地徑的徑級(jí)數(shù)量分布,編制靜態(tài)生命表,并進(jìn)行譜分析,研究黃果厚殼桂人工林幼樹群體的數(shù)量變化特征,為更好地了解黃果厚殼桂人工林的變化規(guī)律和林木生長(zhǎng)特性提供參考,為黃果厚殼桂資源的開發(fā)利用、種質(zhì)資源保護(hù)提供科學(xué)依據(jù)。
1 材料與方法
1.1 試驗(yàn)地概況
試驗(yàn)地位于廣西南寧市南郊的良鳳江國(guó)家森林公園內(nèi)(108°15'~108°22'E,22°34'~22°46'N),地處北回歸線以南,屬亞熱帶季風(fēng)性氣候,冬季較短,夏季較長(zhǎng)。全年日照充足,≥10 ℃年積溫約7 600 ℃,年均氣溫21.6 ℃。干濕季節(jié)分明,雨季集中在4— 9月。地形以丘陵地貌為主,也有少量的階地;土壤呈酸性,多為棕紅色或磚紅色,由砂頁(yè)巖發(fā)育而成,有機(jī)質(zhì)含量較少。
1.2 調(diào)查方法
該黃果厚殼桂人工林于1981年引種種植,面積約0.2 hm2,郁閉度在0.8 以上,林下有較多天然更新的幼苗和幼樹。2020年8月下旬,對(duì)黃果厚殼桂人工林進(jìn)行全面踏查。在能夠代表林分特征和立地條件相似的地段設(shè)置3 個(gè)10 m×10 m 的樣方,樣方之間間隔5 m。采用卷尺(精確至0.01 m)和測(cè)高器測(cè)量樣方內(nèi)每株幼樹的高度,用游標(biāo)卡尺(精確至0.01 mm)測(cè)量幼樹地徑(距離地面高度10 cm 處)。幼樹定義為樹高大于30 cm 且胸徑小于5 cm 的個(gè)體。
1.3 分析方法
1.3.1 生命表的編制
本研究生命表的編制用徑級(jí)結(jié)構(gòu)替代年齡結(jié)構(gòu)[10]。本研究共獲得713 株黃果厚殼桂幼樹數(shù)據(jù),以2.5 mm的地徑為起測(cè)徑級(jí),每增加5 mm為1個(gè)徑級(jí),可將幼樹地徑分為6個(gè)徑級(jí),即Ⅰ(2.5 mm ≤D<7.5 mm)、Ⅱ(7.5 mm ≤D<12.5 mm)、Ⅲ(12.5 mm ≤D<17.5 mm)、Ⅳ(17.5 mm ≤D<22.5 mm)、Ⅴ(22.5 mm ≤D<27.5 mm)和Ⅵ(D ≥27.5 mm)。生命表中各項(xiàng)參數(shù)的計(jì)算公式如下[10-11]:
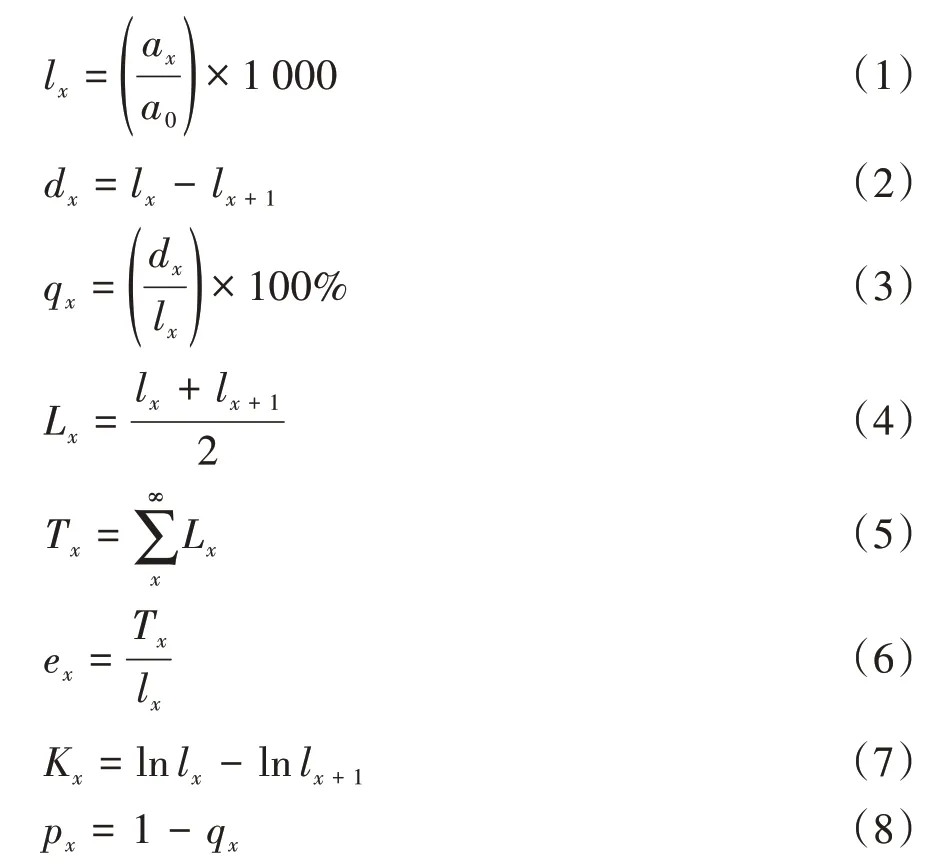
式中,x為徑級(jí);ax為在x徑級(jí)內(nèi)的幼樹個(gè)體數(shù);a0為第Ⅰ徑級(jí)內(nèi)的幼樹個(gè)體數(shù);lx為第x徑級(jí)的標(biāo)準(zhǔn)化存活個(gè)體數(shù);lnlx為標(biāo)準(zhǔn)化存活個(gè)體數(shù)lx的對(duì)數(shù)值;dx為從x到x+1 徑級(jí)間隔期內(nèi)的死亡數(shù);qx為從x到x+1 徑級(jí)間隔期內(nèi)的死亡率;Lx為從x到x+1 徑級(jí)間隔期內(nèi)的存活數(shù);Tx為從第x徑級(jí)到超過(guò)第x徑級(jí)的個(gè)體數(shù);ex為第x徑級(jí)個(gè)體的平均期望壽命;Kx為消失率;px為從x到x+1徑級(jí)間隔期內(nèi)的存活率[11]。
1.3.2 曲線繪制及分析
以徑級(jí)(x)為橫坐標(biāo)、死亡率(qx)和消失率(Kx)為縱坐標(biāo)繪制黃果厚殼桂幼樹群體死亡率和消失率曲線;以徑級(jí)為橫坐標(biāo)、標(biāo)準(zhǔn)化存活個(gè)體數(shù)(lx)的對(duì)數(shù)值(lnlx)為縱坐標(biāo)繪制存活曲線。
存活曲線可以直觀地表達(dá)出幼樹群體的存活過(guò)程。Deevey 存活曲線分為3 種類型[12]。Deevey-Ⅰ型又稱為A型,曲線整體呈凸型,表示該幼樹群體在初期時(shí),幼體死亡率非常低,絕大多數(shù)個(gè)體都能存活至平均壽命,當(dāng)達(dá)到平均壽命時(shí),短期內(nèi)大量個(gè)體死亡;Deevey-Ⅱ型又稱為B 型,曲線整體呈對(duì)角直線,表示該群體各徑級(jí)的死亡率相等;Deevey-Ⅲ型又稱為C型,曲線整體呈凹型,表示該群體在初期時(shí)死亡率高,隨著徑級(jí)的增長(zhǎng),死亡率逐漸降低且趨于穩(wěn)定。
采用SPSS 統(tǒng)計(jì)軟件對(duì)黃果厚殼桂幼樹群體的存活曲線進(jìn)行擬合,檢驗(yàn)其是否符合Deevey-Ⅱ型或Deevey-Ⅲ型[13],建立的數(shù)學(xué)模型如下:
Deevey-Ⅱ型:

Deevey-Ⅲ型:

式中,Nx為第x徑級(jí)內(nèi)個(gè)體的存活數(shù);N0為該群體形成初期的個(gè)體存活數(shù);b為死亡率。
1.3.3 生存分析
為了更好地分析黃果厚殼桂幼樹群體的生存規(guī)律,引入4種函數(shù)對(duì)其進(jìn)行生存分析,生存率函數(shù)S(i)、積累死亡率函數(shù)F(i)、死亡密度函數(shù)f(i)和危險(xiǎn)率函數(shù)λ(i),計(jì)算公式如下[14]:

式中,pi為存活率;qx為死亡率;hi為徑級(jí)寬度。
根據(jù)4 個(gè)生存函數(shù)的計(jì)算結(jié)果繪制生存率曲線、累計(jì)死亡曲線、死亡密度曲線和危險(xiǎn)率曲線。
1.3.4 群體數(shù)量預(yù)測(cè)
運(yùn)用時(shí)間序列分析一次移動(dòng)平均法預(yù)測(cè)黃果厚殼桂幼樹群體的年齡結(jié)構(gòu),公式如下[15]:

式中,t為徑級(jí);n為需要預(yù)測(cè)的時(shí)間;Mt為經(jīng)過(guò)n個(gè)徑級(jí)時(shí)間后t徑級(jí)種群存活數(shù)的預(yù)測(cè)值;Xk為第k徑級(jí)的種群存活數(shù)。本研究將預(yù)測(cè)經(jīng)過(guò)2、3、4、5和6個(gè)徑級(jí)時(shí)間后各徑級(jí)的存活數(shù)。
1.3.5 譜分析
譜分析可用于研究林木種群年齡結(jié)構(gòu)更替過(guò)程的周期性及波動(dòng)性,是Fourier 級(jí)數(shù)的展開,可以寫成正弦波形式[16]:
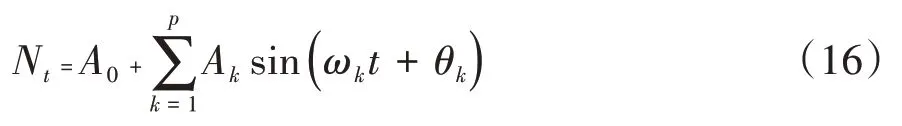
式中,A0為周期變化的均值;Ak為各諧波振幅值;ωk和θk分別為諧波頻率和相角;Nt為t時(shí)刻種群大小。
譜分析正弦波形式中的各個(gè)參數(shù)由下列數(shù)學(xué)模型得出:
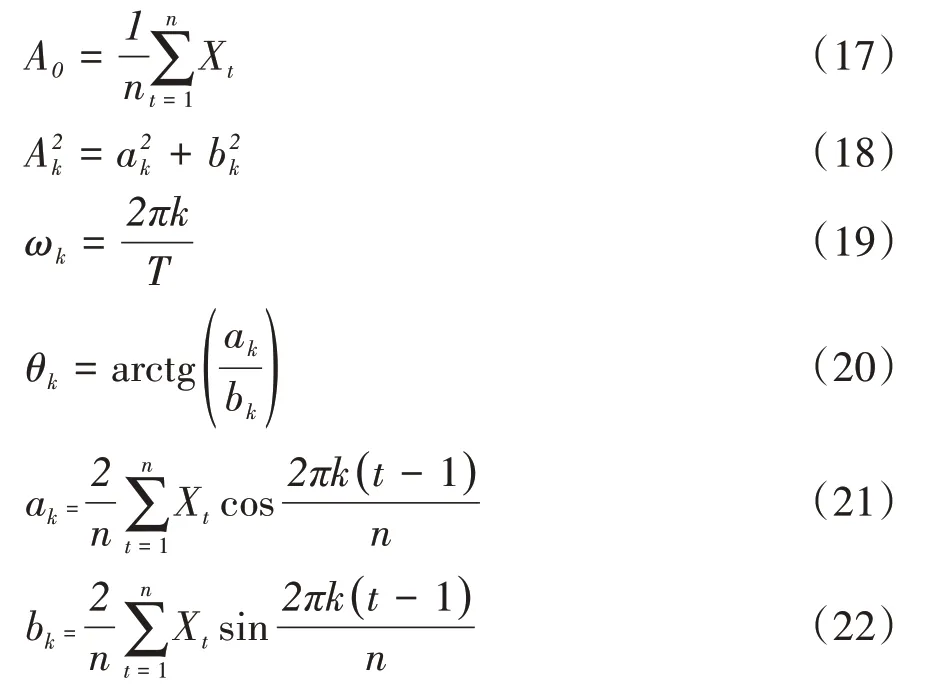
式中,ak和bk為參數(shù)估計(jì)值;將種群各徑級(jí)個(gè)體數(shù)分布視為時(shí)間序列t,Xt表示t年齡序列時(shí)的個(gè)體數(shù);n為系列總長(zhǎng)度;P=n∕2為諧波的總個(gè)數(shù);T為正弦波的基本周期,即時(shí)間序列t的最長(zhǎng)周期,即資料總長(zhǎng)度,即T=n為已知。
本研究中,黃果厚殼桂幼樹群體共有6個(gè)徑級(jí),將群體各徑級(jí)個(gè)體數(shù)分布視為時(shí)間序列t(t= 1,2,3,4,5,6);系列總長(zhǎng)度n為6;諧波的總個(gè)數(shù)為P=n∕2=6∕2=3,需計(jì)算振幅值A(chǔ)k(k=1,2,3);T=n=6。諧波對(duì)應(yīng)徑級(jí)的計(jì)算方法如下:

式中,D為地徑;n為徑級(jí)總數(shù);hi為徑級(jí)寬度;k= 1,2,3。通過(guò)D值所處的范圍得出諧波對(duì)應(yīng)的徑級(jí)。
由于黃果厚殼桂幼樹群體各徑級(jí)個(gè)體存活數(shù)相差較大,在利用譜分析公式進(jìn)行計(jì)算時(shí)采用對(duì)數(shù)處理,用Xt'代替公式中的Xt(即為表1 中第x徑級(jí)內(nèi)的幼樹個(gè)體數(shù)ax)[17]。處理后對(duì)各波形的振幅值A(chǔ)k(k=1,2,3)進(jìn)行計(jì)算。
2 結(jié)果與分析
2.1 徑級(jí)結(jié)構(gòu)
黃果厚殼桂幼樹群體的徑級(jí)結(jié)構(gòu)呈倒金字塔型。各徑級(jí)個(gè)體存活數(shù)表現(xiàn)為Ⅰ>Ⅱ>Ⅲ>Ⅳ>Ⅴ>Ⅵ(圖1)。第Ⅰ徑級(jí)的個(gè)體數(shù)最多,占總數(shù)的38.43%;其次為第Ⅱ徑級(jí),占總數(shù)的31.98%;第Ⅲ~Ⅵ徑級(jí)共占總數(shù)的29.59%。隨徑級(jí)增加,幼樹個(gè)體數(shù)減少。
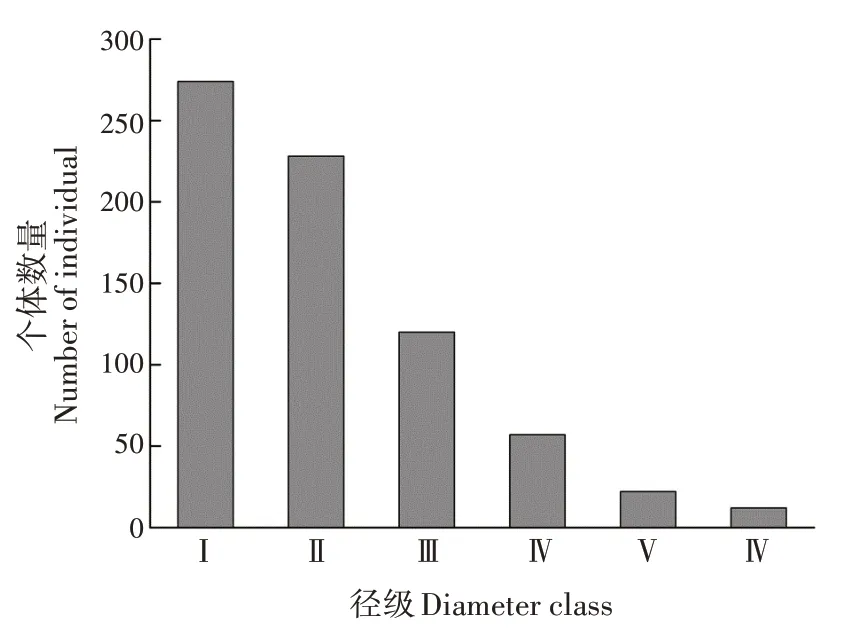
圖1 黃果厚殼桂幼樹群體徑級(jí)結(jié)構(gòu)Fig.1 Diameter class structure of C.concinna sapling popu?lation
2.2 靜態(tài)生命表
黃果厚殼桂幼樹群體中,第Ⅰ、Ⅱ徑級(jí)的ax和lx較大,分別為274、1 000 和228、832;第Ⅵ徑級(jí)的ax和lx最小,分別為12和44(表1)。ax和lx均隨徑級(jí)增大而減小,原因可能是黃果厚殼桂幼樹群體中大部分個(gè)體年齡較小。
dx呈現(xiàn)明顯的波動(dòng)性,第Ⅱ徑級(jí)最大(394),第Ⅴ徑級(jí)最小(36),表明在第Ⅱ~Ⅲ徑級(jí)間隔期內(nèi)死亡數(shù)最多,在大徑級(jí)的間隔期內(nèi)死亡數(shù)最少(表1)。原因可能是隨徑級(jí)增加,幼樹個(gè)體在生長(zhǎng)過(guò)程中受種內(nèi)競(jìng)爭(zhēng)或環(huán)境等因素的影響而被淘汰,變成中、大徑級(jí)的個(gè)體數(shù)量減少。

表1 黃果厚殼桂幼樹群體靜態(tài)生命表Tab.1 Static life table of C.concinna sapling population
ex反映各徑級(jí)的平均生存能力。ex呈下降趨勢(shì),第Ⅰ徑級(jí)最大(2.10),第Ⅵ徑級(jí)最小(0.50)。在第Ⅰ至Ⅱ、第Ⅳ至第Ⅵ徑級(jí),ex明顯降低。黃果厚殼桂幼樹耐陰喜陰[18],林冠下的小徑級(jí)個(gè)體更易存活,但由于種內(nèi)競(jìng)爭(zhēng),群體生存能力下降;大徑級(jí)個(gè)體因?yàn)樯碓蛏婺芰ψ儾睿贁?shù)大徑級(jí)個(gè)體較早分布在林分中,占據(jù)了更好的生長(zhǎng)空間[19],沒(méi)有被淘汰。
2.3 幼樹群體曲線
2.3.1 死亡率和消失率曲線
qx和Kx的變化趨勢(shì)基本一致,存在一定波動(dòng)但整體呈增加的趨勢(shì),符合一般的種群規(guī)律(圖2)。qx和Kx在第Ⅰ徑級(jí)均最小,在第Ⅳ徑級(jí)達(dá)到峰值后均下降,說(shuō)明第Ⅰ~Ⅳ徑級(jí)的幼樹間競(jìng)爭(zhēng)逐漸加劇或生存環(huán)境逐漸惡化,第Ⅳ徑級(jí)后競(jìng)爭(zhēng)減緩或生存環(huán)境改善。
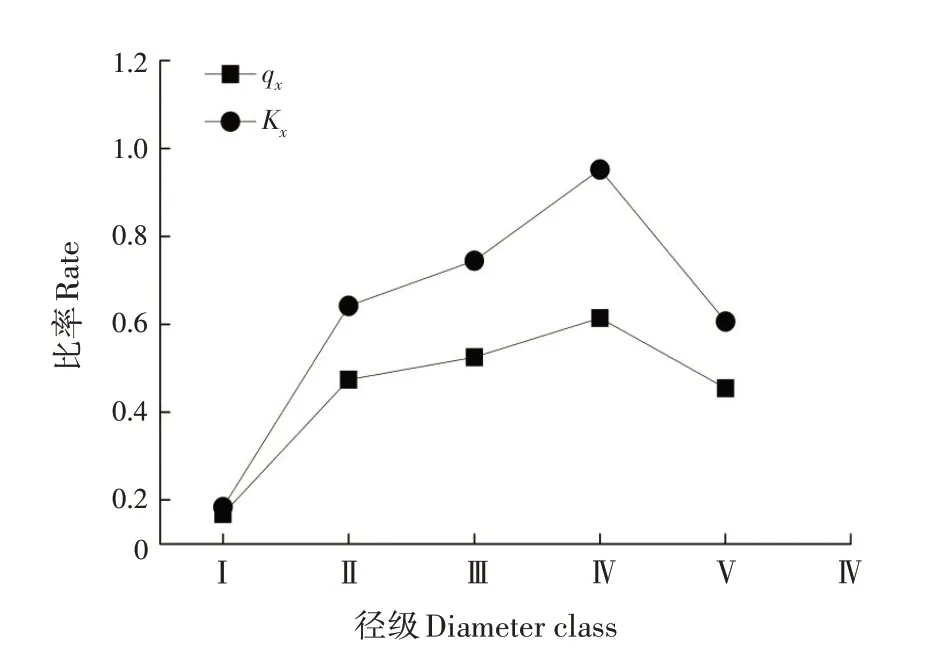
圖2 黃果厚殼桂幼樹群體死亡率(qx)和消失率(Kx)曲線Fig.2 Curves for mortality rate and vanishing rate of C.con?cinna sapling population
2.3.2 存活曲線
存活曲線呈近乎線性下降趨勢(shì)(圖3)。Deevey-Ⅱ指數(shù)函數(shù)模型的R2= 0.97,Deevey III 冪函數(shù)模型的R2= 0.84,說(shuō)明指數(shù)函數(shù)模型的擬合效果好,優(yōu)于冪函數(shù)模型(表2)。黃果厚殼桂幼樹群體的存活曲線屬于Deevey-Ⅱ型,表明該群體死亡率相對(duì)穩(wěn)定。

表2 黃果厚殼桂幼樹群體存活曲線的檢驗(yàn)?zāi)P蚑ab.2 Test models of survival curves of C.concinna sapling population

圖3 黃果厚殼桂幼樹群體存活曲線Fig.3 Survival curve of C.concinna sapling population
2.3.3 生存函數(shù)曲線
黃果厚殼桂幼樹群體的F(i)呈單調(diào)上升,S(i)呈單調(diào)下降,兩者互補(bǔ)(表3)。在接近第Ⅱ徑級(jí)時(shí),F(xiàn)(i)等于S(i),說(shuō)明在第Ⅱ徑級(jí)之前群體達(dá)到平衡狀態(tài),隨后F(i)超過(guò)S(i),表明群體進(jìn)入衰減期。群體在第Ⅰ~Ⅲ徑級(jí)有相對(duì)較高的ex,但個(gè)體密度大,生長(zhǎng)競(jìng)爭(zhēng)激烈,導(dǎo)致大批個(gè)體在這3個(gè)徑級(jí)被淘汰,使f(i)較高;第Ⅳ徑級(jí)后,f(i)趨于平緩,源于個(gè)體間競(jìng)爭(zhēng)趨于緩和,淘汰減少。λ(i)在第Ⅴ徑級(jí)出現(xiàn)頂峰,說(shuō)明第Ⅴ徑級(jí)幼樹死亡風(fēng)險(xiǎn)最高,這與生理性死亡等原因有關(guān)。從幼樹群體數(shù)量上看,小、中徑級(jí)數(shù)量多但減少快,大徑級(jí)數(shù)量少但減少趨于平緩,幼樹群體整體呈衰減趨勢(shì)。
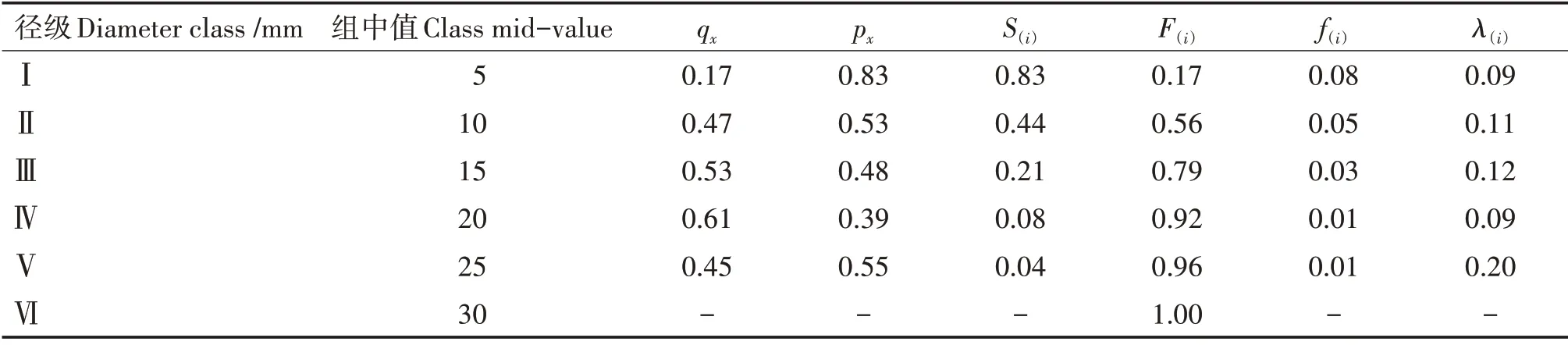
表3 黃果厚殼桂幼樹群體的生存函數(shù)估算值Tab.3 Estimated values of survival functions of C.concinna sapling population
2.3.4 群體數(shù)量的時(shí)間序列分析
以黃果厚殼桂幼樹群體各徑級(jí)個(gè)體數(shù)量為基礎(chǔ),采用時(shí)間序列分析的方法預(yù)測(cè)該群體第Ⅱ~Ⅵ徑級(jí)后的數(shù)量(表4)。除了第Ⅱ徑級(jí)在兩個(gè)徑級(jí)后數(shù)量下降近一半外,其他各徑級(jí)均呈增長(zhǎng)趨勢(shì),第Ⅲ徑級(jí)增長(zhǎng)最明顯,說(shuō)明該群體的小、中徑級(jí)個(gè)體數(shù)量充足,短期內(nèi)占群體主導(dǎo)地位。第Ⅲ徑級(jí)后,主要為大徑級(jí),整體上仍處于數(shù)量增長(zhǎng)期,與小徑級(jí)數(shù)量充足有關(guān)。整體上看,各個(gè)徑級(jí)個(gè)體數(shù)量在之后的各徑級(jí)階段均上升,但在經(jīng)過(guò)6個(gè)徑級(jí)后,群體存活數(shù)由713 個(gè)下降至119 個(gè),說(shuō)明該黃果厚殼桂幼樹群體屬于衰減型群體。
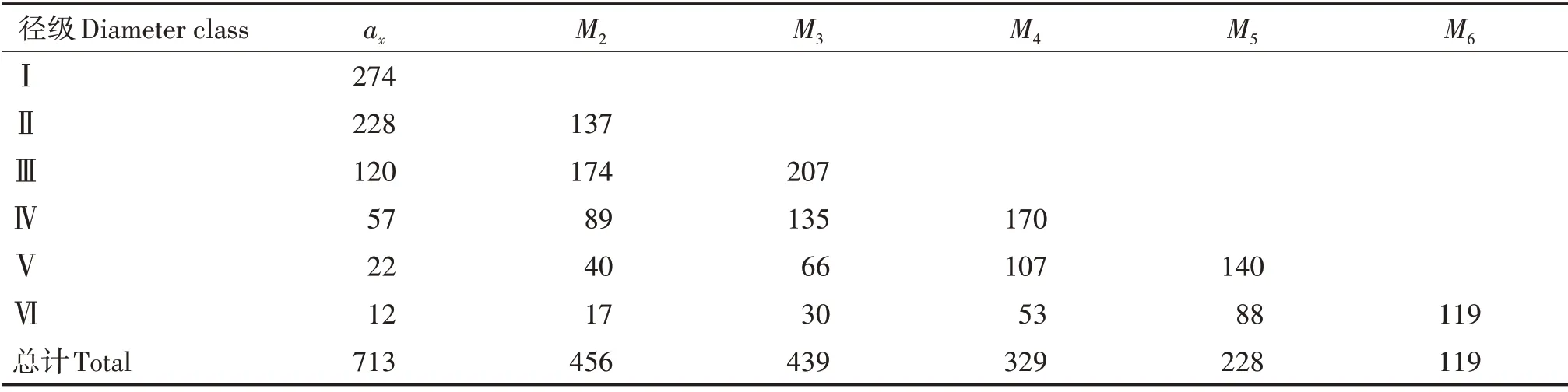
表4 黃果厚殼桂幼樹群體動(dòng)態(tài)變化的時(shí)間序列分析Tab.4 Analysis of time series of C.concinna sapling population
2.4 譜分析
A1至A3的振幅呈單調(diào)遞減的趨勢(shì),各諧波的振幅相差較大,表明該群體數(shù)量變化的波動(dòng)并非單一周期,即大周期內(nèi)包含小周期,且各周期的作用強(qiáng)度不同(表5)。諧波A2和A3對(duì)應(yīng)的周期分別是基波A1的1∕2 和1∕3,整體上隨著周期的增大而增大[20]。基波的周期取決于黃果厚殼桂幼樹群體的特性,能夠表現(xiàn)出該群體數(shù)量變化的波動(dòng)性。除去周期變化的平均值(A0)后,基波A1振幅值最大,表明群體的數(shù)量變化受基波的影響十分顯著。

表5 黃果厚殼桂幼樹群體的周期性波動(dòng)Tab.5 Periodic fluctuation of C.concinna sapling population
黃果厚殼桂幼樹群體的數(shù)量動(dòng)態(tài)除受基波的影響外,還表現(xiàn)出小周期波動(dòng)。第1 次小周期為諧波A2,反應(yīng)在6 × 5∕2 = 15 mm,位于空間序列徑級(jí)12.5~17.5 mm 處,對(duì)應(yīng)第Ⅲ徑級(jí),這一小周期波動(dòng)與幼樹的生理特性或林分的環(huán)境有關(guān)。第2次小周期也是理論上的最小周期(A3),反映在6×5∕3=10 mm,位于空間序列徑級(jí)7.5~12.5 mm 處,對(duì)應(yīng)第Ⅱ徑級(jí),這一小周期波動(dòng)與幼樹間的激烈競(jìng)爭(zhēng)導(dǎo)致部分個(gè)體死亡有關(guān),這種數(shù)量調(diào)節(jié)更有利于群體的穩(wěn)定發(fā)展。
3 討論
種群是構(gòu)成生物群落和生態(tài)系統(tǒng)的基本單位,幼樹的群體結(jié)構(gòu)可反映種群的發(fā)展趨勢(shì)。研究林分特定種群的幼樹結(jié)構(gòu)特征和數(shù)量變化規(guī)律,對(duì)于林分的可持續(xù)經(jīng)營(yíng)有重要意義。了解幼樹群體的存活率和死亡率等變化規(guī)律,是研究種群的基本任務(wù)之一[21-23]。
徑級(jí)結(jié)構(gòu)可體現(xiàn)種群的結(jié)構(gòu)特征和穩(wěn)定性,靜態(tài)生命表和存活曲線從宏觀角度觀測(cè)群體數(shù)量的變化規(guī)律。黃果厚殼桂人工林幼樹群體的徑級(jí)結(jié)構(gòu)呈倒金字塔型,各年齡結(jié)構(gòu)較為完整,由于黃果厚殼桂的耐蔭特性,小徑級(jí)個(gè)體在林冠下更易存活,這與小徑級(jí)ex較大相一致;由于個(gè)體密度大,隨徑級(jí)增加,個(gè)體間競(jìng)爭(zhēng)加劇,群體生存能力逐漸下降,被淘汰個(gè)體也逐漸增加[24],少數(shù)大徑級(jí)個(gè)體占據(jù)更好的生長(zhǎng)空間而生存下來(lái)。該幼樹群體在第Ⅳ徑級(jí)時(shí)qx和Kx出現(xiàn)峰值,這是由于在第Ⅳ徑級(jí),種內(nèi)競(jìng)爭(zhēng)和環(huán)境篩選導(dǎo)致幼樹在生長(zhǎng)發(fā)育至高徑級(jí)過(guò)程中死亡[25]。存活曲線呈下降趨勢(shì),說(shuō)明林冠下的蔭蔽環(huán)境有利于小徑級(jí)個(gè)體存活;吳敏等[26]發(fā)現(xiàn)光照因素可能導(dǎo)致幼苗難以生長(zhǎng)發(fā)育為幼樹,對(duì)黃果厚殼桂人工林的管理應(yīng)充分考慮該樹種的光適應(yīng)性,為其提供適當(dāng)?shù)恼谑a環(huán)境,保證小徑級(jí)苗木的生長(zhǎng)發(fā)育。存活曲線屬于Deevey-Ⅱ型,隨徑級(jí)增大,個(gè)體存活數(shù)量逐漸減少,說(shuō)明該幼樹群體擁有較多的小徑級(jí)個(gè)體和少量的中、大徑級(jí)個(gè)體,群體構(gòu)成較為穩(wěn)定[27]。在小徑級(jí)階段,種內(nèi)競(jìng)爭(zhēng)激烈,生存空間或資源不足,自疏或它疏作用造成淘汰,僅保留少部分發(fā)育至大徑級(jí)。從生存函數(shù)來(lái)看,幼樹群體在小、中徑級(jí)時(shí)期的生存壓力主要來(lái)自于種內(nèi)競(jìng)爭(zhēng),大徑級(jí)時(shí)期的生存壓力主要來(lái)自于環(huán)境篩選。通過(guò)時(shí)間序列預(yù)測(cè)可知,該幼樹群體小徑級(jí)個(gè)體數(shù)多,未來(lái)有較多幼樹可發(fā)育成大徑級(jí)。對(duì)于該林分的經(jīng)營(yíng)重點(diǎn)應(yīng)放在小徑級(jí)幼樹的管理上,比如改善小徑級(jí)個(gè)體的種內(nèi)競(jìng)爭(zhēng)。隨著種群個(gè)體數(shù)的增加,相鄰的個(gè)體間會(huì)相互影響(密度效應(yīng)),通過(guò)合理的措施調(diào)控密度效應(yīng),可以改善幼樹群體的生存狀況。
譜分析表明,該幼樹群體的數(shù)量變化明顯受生命周期生物特性因素的影響[28],且在數(shù)量變化的大周期內(nèi)有一部分小周期。從整體來(lái)看,該群體仍以小徑級(jí)黃果厚殼桂幼樹個(gè)體為主,未進(jìn)入成熟、衰老的階段。譜分析結(jié)果只能體現(xiàn)當(dāng)前幼樹群體生長(zhǎng)情況的變化規(guī)律,不能體現(xiàn)黃果厚殼桂種群整體在整個(gè)基本周期的數(shù)量變化規(guī)律。
