牛支原體病流行病學及其診斷技術研究進展
2022-01-27 01:19:20王天宇李繼東張志誠郭亞男
畜牧與獸醫 2021年12期
王天宇,李繼東,張志誠,郭亞男
(1. 寧夏大學農學院,寧夏 銀川 750021;2. 中國動物衛生與流行病學中心,山東 青島 266032;3. 寧夏農林科學院動物科學研究所,寧夏 銀川 750002)
牛支原體是柔膜體綱、支原體目、支原體科、支原體屬、革蘭氏染色呈陽性的原核生物,是一種能引起牛肺炎、乳房炎和關節炎等疾病的病原微生物[1]。牛支原體最早于1961年在美國發現,隨后在全球傳播、流行。在我國2008年辛九慶等[2]首次從患肺炎的犢牛肺臟中分離出牛支原體;2017年在新西蘭奶牛群中暴發牛支原體病,造成多達15.8萬頭牛被撲殺,截止目前,挪威是唯一沒有被牛支原體感染的養牛國家。牛支原體常與多殺性巴氏桿菌和溶血曼海姆氏菌等混合感染,給養牛業造成了嚴重的經濟損失。牛支原體和無乳支原體、絲狀支原體絲狀亞種等的病原結構以及其致病的臨床癥狀很相似,難以鑒別診斷及預防控制[3]。牛支原體基因組具有的可變脂蛋白家族基因及眾多的可插入序列導致牛支原體極易突變,能在不同的地理環境中獨立進化,在國際上存在多種基因分型的菌株,表現出高度的多樣性[4]。
1 病原學與流行病學
1.1 病原學
牛支原體與其他支原體相似,具有多種形態,在顯微鏡下呈球形、橢圓形和棒形等;細胞膜由三層膜構成,內、外層由電子密度高的蛋白質與糖類構成,中間層主要由電子密度低的脂質構成,細胞質內含有雙鏈DNA和核糖體,沒有內質網、高爾基體等細胞器[1]。牛支原體是能在無活細胞培養物中存活的最小微生物,其基因組大小僅為大腸桿菌基因組的1/5,能夠通過0.45 μm微孔濾膜,在加壓情況下能通過0.22 μm的微孔濾膜[1]。牛支原體的基因組大小為9.48~10.39 kb,C+G含量較低,僅占27.8%~32.9%。以牛支原體國際參考菌株PG45為例,其基因組包含826個開放閱讀框(ORF),編碼密度為89%,能編碼眾多基因,包括6個rRNA、 34個tRNA基因和96個假定的脂蛋白基因(包括13個vsp基因簇)等。染色體上還含有54個插入序列(IS)和2個整合共軛元件(ICE)即ICEB-1和ICEB-2,大小分別為2.33 kb和3.74 kb,ICEB-1與無乳支原體PG2的ICEA非常相似,所以,牛支原體和無乳支原體被認為是由同一個祖先進化而來的[5]。牛支原體兼性厭氧,能在無活細胞的培養基上生長,適宜在5%~10%的CO2環境下生存,因為基因組較小,缺乏完整的酶系統,代謝活性受到限制,因此不能分解葡萄糖和精氨酸,而是利用有機酸、乳酸和丙酮酸為其生長能源,生長的菌液通常透亮不渾濁,低倍鏡下菌落呈典型的煎蛋外觀[1, 6]。牛支原體對環境的抵抗力較強,4 ℃的海綿或牛奶中能存活2個月,-20 ℃下能存活半年,-70 ℃下能存活一年以上;對高溫敏感,65 ℃熱處理2 min或70 ℃熱處理1 min活性喪失。
1.2 流行病學
1961年,Hale等從美國一例嚴重的牛乳腺炎病例中首次分離出牛支原體,由于血清學、生化特性與無乳支原體相似,最初命名為無乳支原體牛變種,被認為是造成牛乳腺炎的病原體之一。這種支原體自發現以來,在世界范圍內廣泛傳播。20世紀60年代在亞洲的以色列和歐洲的西班牙發現了該支原體,隨后在20世紀70年代擴展到大洋洲的澳大利亞以及法國等歐洲國家[3]。1975年,Thomas等在英國率先從患有肺炎癥狀的3~5月齡的荷斯坦小牛中分離出無乳支原體牛變種,動物接種試驗發現該支原體可以導致牛發生肺炎,1976年該支原體被正式命名為牛支原體。隨后,由牛支原體引起的肺炎與乳腺炎相繼在世界各國暴發并報道,包括丹麥(1981)、日本(1982)、瑞士(1983)、摩洛哥(1985)、韓國(1989)、巴西(1989)、愛爾蘭(1994)、智利(1997)、南非(2005)、捷克共和國(2007)等。我國于2008年由辛九慶等[2]首次從患肺炎的犢牛肺臟中分離到牛支原體,現除海南省未發現牛支原體肺炎疫情外,各地相繼出現牛支原體感染病例[7-8]。目前,牛支原體呈全球傳播,之前沒有牛支原體感染的芬蘭(2012)和新西蘭(2017)也確診暴發了牛支原體疫病[9-10]。
1.3 流行菌株
1961年美國首次分離出的牛支原體菌株PG45已于2011年完成了全基因組測序,成為牛支原體國際參考株[5]。Rosales等[11]對全球的多個牛支原體分離株進行VNTR分析,發現存在兩個主要種群即A和B組,各組包含的分離株與其地理起源高度相關。應用MLST分析發現,源自歐洲的牛支原體分離株主要為CC1組,以色列、澳大利亞和中國的牛支原體分離株主要為CC2組。在CC1組中鑒定出兩個常見祖先菌株序列型(ST2和ST19型),ST2型是代表性牛支原體中最常見的ST型,ST19型是僅來自歐洲大陸(西班牙、德國、法國和立陶宛)的代表性分離株,后者被證明與美國的PG45菌株具有極大的相似性,這一發現支持了牛支原體PG45與歐洲分離株之間存在共同祖先的假說,可能與過去兩大洲之間的活牛及其產品貿易有直接關系。CC2組包括了所有中國分離株以及大多數來自以色列和澳大利亞的分離株ST10、ST33和ST34型,該組分離株在地理起源方面表現出高度的多樣性,與大多數歐洲分離株分開,支持了牛支原體分離株在地理上獨立進化的理論。Yair等[12]和Menghwar等[13]研究發現,中國、以色列和澳大利亞的流行分離株主要為ST10型,提出了中國與以色列的分離株可能來源于澳大利亞的結論,這與中國和以色列從澳大利亞大量進口牛只有關。
1.4 傳播途徑
牛支原體能在牛群中廣泛傳播,包括水牛、肉牛、奶牛、野牛在內的所有種類,以及所有年齡段的牛都易感[14]。牛支原體病在牛群中暴發主要是由輸入牛只與運輸工具攜帶病原體傳播導致的,患病牛和病原攜帶牛是主要的傳染源,被感染的牛可能成為無癥狀的攜帶者,持續排出病原體長達數年之久[7]。氣溶膠傳播是最主要的傳播方式,牛支原體通過呼吸道排出,能在氣溶膠內存活數月甚至長達幾年之久,牛之間的直接接觸、排出的分泌液、唾液和糞便都能攜帶病原造成健康牛的感染。牛支原體也可以發生垂直傳播,懷孕母牛能通過胎盤傳播給胎兒,還可以通過初乳傳播給新生犢牛,在公牛的精液中也檢測到了牛支原體的存在[15-16]。牛支原體具有黏附性,寄生于黏膜、纖毛等部位的細胞,可隨部分脫落的細胞排出體外,這個排泄過程呈周期性,一般在感染后一周脫落,最快能在24 h后脫落,氣候變化、過度擁擠或運輸等應激因素可能會加速這一過程使牛更容易受到感染,并引起疾病暴發。除了直接傳播,牛支原體還能通過環境因素進行傳播,能通過運輸車輛、飼料、圍墻欄桿、飼喂用具和墊料等傳播。有研究表明,牛支原體能在4 ℃的牛奶中存活2個月,在肥料中存活37 d,棉花上存活18 d,稻草上存活13 d,木材和不銹鋼上存活1~2 d[3]。除牛之外的一些其他動物也是牛支原體的攜帶者,如綿羊、山羊、豬、雞和鹿等都是牛支原體的儲存宿主,牛支原體跨物種感染其他動物后又反過來感染牛,造成疾病的循環往復[17-20]。牛支原體通常是不感染人的,但由于長期接觸感染牛的牛奶、糞便,農場員工體內檢測到了牛支原體[21]。
2 診斷方法
由于牛支原體能與多種病原混合感染,因此對于牛支原體病的鑒別診斷顯得尤為重要。目前主要的實驗室診斷方法有:病原的分離鑒定、分子生物學方法和免疫學方法。其中分子生物學方法主要包括聚合酶鏈式反應(PCR)、等溫擴增技術、DNA微陣列技術、遺傳鑒定;免疫學方法主要包括免疫組織化學技術(IHC)和酶聯免疫吸附試驗(ELISA)。
2.1 分離鑒定
由于支原體結構簡單,因此無法合成氨基酸,并且無法完全或部分合成脂肪酸。為了滿足這種高營養需求,支原體培養基通常富含牛心浸出液、血清、酵母提取物和蛋白胨等,培養基pH值為7.3~7.8[1]。接種后的培養基在37 ℃、5% CO2條件下,孵育7~10 d,光學顯微鏡下可見牛支原體菌落出現“煎蛋”樣形態。牛支原體革蘭氏染色呈陽性,因為沒有細胞壁,革蘭氏染色困難,吖啶橙染色效果更好,染色后菌體呈亮紅色或橘紅色[22]。雖然通過分離培養法鑒定牛支原體相對簡單且成本低廉,但存在局限性。如在采集樣品時動物必須有活菌排出,這和牛支原體的排泄模式有關。在慢性和亞臨床支原體乳腺炎病例中,常有牛支原體呈間歇性排出的情況,其間隔時間甚至能長達56 d,這可能導致單個樣本的診斷失敗,因此建議應至少采集3份樣品并間隔3~4 d,收集后進行培養鑒定[23]。牛支原體的儲存對環境要求較高,采集的病料需冷藏并在24 h內送往實驗室,也可以凍存運輸,但凍存會導致活菌數量降低1或2個數量級。此外,牛支原體的培養經常受到其他柔膜體綱病原的污染,分離難度大、分離周期長,不適用于牛支原體的快速檢測[24]。
2.2 分子生物學方法
為了克服常規分離培養的不足,目前已經開發了多種基于核酸的分子檢測方法,這些方法具有檢測速度快、靈敏性和特異性高等優點。由于基于核酸的分子檢測方法是物種特異性的,所以能夠區分不同種類的支原體以及牛支原體的不同基因型。常用牛支原體核酸診斷技術比較見表1。
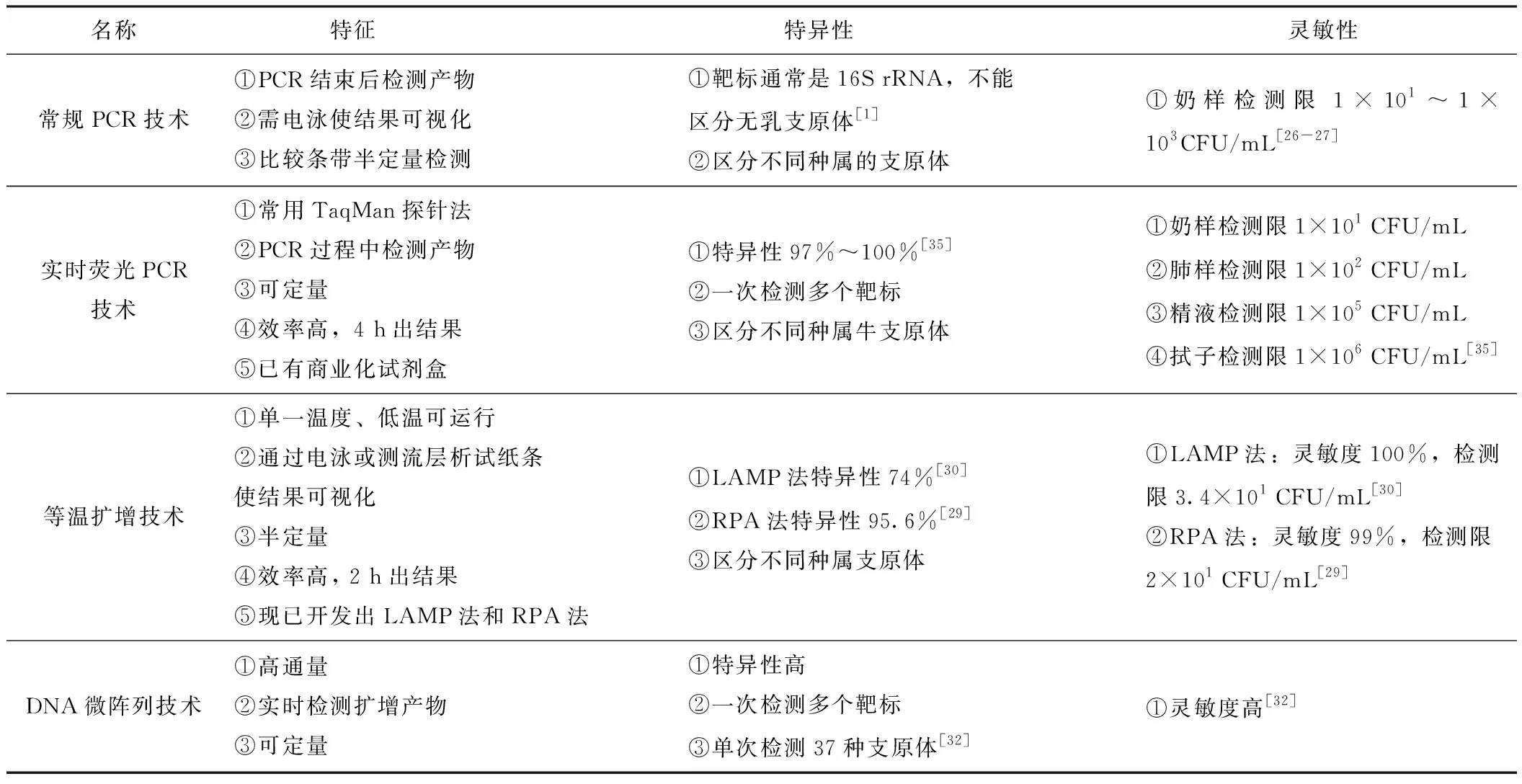
表1 牛支原體核酸診斷技術
2.2.1 PCR技術
應用PCR技術檢測來自各種樣品類型的支原體具有更高的效率、特異性和敏感性。早期主要利用16S rRNA基因做靶標,但由于牛支原體與無乳支原體的16S rRNA序列一致性高達99%,所以經常發生交叉擴增的現象[25]。為了區分不同病原體,設計了針對16S~23S rRNA間隔區的PCR方法,可以區分支原體與無膽甾原體;使用幾個引物組設計的多重PCR,能夠同時鑒別感染牛的幾種不同支原體,在牛奶樣品中的檢測下限低至1×101CFU/mL[26-27]。目前,實時熒光PCR技術也逐漸應用于牛支原體的檢測。由于與無乳支原體16S rRNA高度的同源性,人們選取了特異性更好的DNA修復基因uvrC基因、ATP結合蛋白oppD/F基因和p81基因等作為靶基因進行特異性擴增,Clothier等[28]針對uvrC基因設計開發了實時熒光PCR檢測方法,檢測下限低至2.4×102CFU/mL,靈敏度和特異性良好。
2.2.2 等溫擴增技術
Zhao等[29]針對牛支原體uvrC基因設計開發了重組酶聚合酶擴增技術與側流層析試紙分析(RPA- LFD)相結合的檢測方法,能在30 min內檢測到牛支原體,檢測下限為20個拷貝,靈敏度和特異性分別為99%和95.6%。Bai等[30]針對牛支原體uvrC基因設計開發了LAMP法,與普通PCR方法相比較,在純培養物中測定的靈敏度比PCR高10倍,檢測下限為34個拷貝,該方法的靈敏度和特異性分別為100%和74%,靈敏度高,特異性較準確,適宜于在生產中檢測牛支原體。Yumiko等[31]針對oppD/F基因區域設計6對引物,改進后的LAMP法,其靈敏度、特異性分別為97.2%和90.9%(kappa系數為0.823 1)。中國農業大學吳文學團隊研發的牛支原體環介導等溫擴增檢測試劑盒于2020年獲得了國家新獸藥注冊證書((2020)新獸藥證字35號)。
2.2.3 DNA微陣列技術
Schnee等[32]根據基因組高度保守區16S~23S rRNA間隔區和可變區交替分布的23S rRNA序列,對38個支原體、3個無膽甾原體和3個解脲脲原體設計了70個寡核苷酸探針;為了擴展微陣列的區分能力,將tuf基因視為另一個靶標,設計了86個滿足基本選擇標準的寡核苷酸探針,經過PCR擴增和生物素化后,在陣列上探針與DNA的雜交可特異性鑒定目標病原,一次最多可以檢測出37種支原體。微陣列測定法的主要優點包括操作簡便、快速和信息量高,分析靈敏度與實時熒光PCR相當,并且無需培養即可檢查野外樣品。
2.2.4 遺傳鑒定
由于牛支原體基因組相對于病毒較大,全基因測序比較困難,人們使用脈沖場凝膠電泳(PFGE)、可變串聯重復序列(VNTR)、多基因座可變串聯重復分析(MLVA)和多基因座序列分型(MLST)等對支原體基因組的幾個管家基因進行獨特的序列分析,用于牛支原體的遺傳鑒定與基因分型。最近提出了3種MLST方案用于研究牛支原體的種群結構、進化和傳播:Manso-Silvan等[33]開發的MLST方案基于4個管家基因(fusA、gyrB、lepA和rpoB),分辨力為0.833;Register等[34]開發的MLST方案基于7個管家基因(adh-1、gltX、gpsA、gyrB、pta-2、tdk、tkt),分辨力為0.923;Rosales等[11]開發改進的MLST方案基于7個管家基因(dnaA、metS、recA、tufA、atpA、rpoD和tkt),分辨力為0.91(95%置信區間為0.88~0.93),后兩種改進方案均優于第一種方案。
2.3 免疫學方法
不同于牛支原體的分離與PCR檢測,免疫學檢測方法無需在牛支原體的排泄期采集樣本,因為動物機體感染牛支原體所產生的抗體可以保持幾個月之久。在病料采集與檢測難度方面,免疫學方法具有極大優勢,通過優化,免疫學方法在檢測牛支原體方面具有良好的靈敏性與特異性。
2.3.1 免疫組織化學技術(IHC)
Hermeye等[36]對膿腫的細支氣管或終末呼吸道、肺臟和腎小管上皮細胞等組織進行了染色檢測,在顯微鏡下組織切片可見棕黃色顆粒,呈彌漫性出現于細胞周圍,即為牛支原體陽性。
2.3.2 酶聯免疫吸附試驗(ELISA)
目前有多種類型的ELISA用于檢測牛支原體,包括間接ELISA、競爭ELISA和阻斷ELISA等。利用VSP重組蛋白或膜脂蛋白作為包被抗原以檢測血清中的IgG,所建立的間接ELISA方法是檢測牛血漿、血清和牛奶等樣本的首選方法,應用最為廣泛。
間接ELISA檢測方法依賴于牛對牛支原體的免疫反應及其產生的抗體。研究顯示,動物機體產生的牛支原體抗體可以持續數月,且不受牛支原體間歇性排泄的影響,這表明間接ELISA適用于牛支原體抗體的檢測,但幾例實驗性感染表明,血清IgG抗體轉化需要2~3周,在該時間段內進行檢測可能顯示陰性結果[37]。間接ELISA方法還適用于對牛奶樣品的檢測,可應用于大罐奶的檢測以大規模分析牛群中牛支原體的感染情況,進行流行病學調查,但單個乳樣本檢測結果往往并不理想[38]。這可能是因為IgG抗體是在血液中產生的,當轉移到牛奶中時導致濃度較低的緣故,并且抗體在牛奶中存在的時間沒有在血液中持久[39]。CABI的入侵物種綱要對ELISA檢測患病牛的鼻拭子、肺、牛奶、關節液和滑膜、精液和生殖器分泌物等樣本與血清樣本的檢測效果進行了比較分析,發現在血清中都能檢測到抗體,而其他樣品只有相關部位發生炎癥時才能檢測出來。由此可知,血清樣本最適用于做牛支原體的間接ELISA檢測樣本,但需要注意在血清抗體轉化后進行檢測診斷,或增加樣本檢測量以提高準確度。
目前已有開發的牛支原體抗體ELISA檢測商品化試劑盒,但陽性檢出率差,靈敏度不足[34,39]。最近Wawegama等[40]建立了一種基于MilA蛋白重組片段的間接ELISA,檢測實驗性感染犢牛,最佳臨界值估計為68.6抗體單位(AU);對于成年牛群,最佳臨界值估計為58.7 AU;在自然條件下,育肥牛的最佳臨界值估計為105 AU。在此臨界值時,診斷靈敏度為94.3%,95%概率區間(PI為89.9%~99.6%),特異性為94.4%, PI為90.3%~99.6%)。與目前的商品化試劑盒相比,Wawegama等[41]開發的間接ELISA方法診斷靈敏性和特異性更佳,更適用于牛支原體的血清學診斷與流行病學調查。中國農業科學院哈爾濱獸醫研究所辛九慶團隊研發的牛支原體ELISA抗體檢測試劑盒于2017年獲得了國家新獸藥注冊證書((2017)新獸藥證字48號)。
3 常用診斷技術比較
不同診斷技術有各自的優點和不足,對牛支原體常用診斷技術比較的結果見表2[35]。

表2 牛支原體常用診斷技術比較[35]
4 小結
我國是養牛大國,牛支原體病嚴重危害我國養牛業,掌握其流行病學特征以及診斷技術是控制和預防該病的關鍵。同時我國也是牛及其產品的進口大國,目前國際上存在多種基因型牛支原體,基因組測序和基因分型鑒定有利于對牛支原體進行分析溯源,有助于降低牛支原體通過國際貿易傳入的風險。不同的診斷方法具有一定的優缺點,使用者需針對不同使用環境、不同的診斷需求,選擇最合適的方法。目前還需要將新材料、新技術和計算機技術結合起來,研究快速和高通量檢測技術,以提高疾病的診斷能力。
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
兒童故事畫報(2019年5期)2019-05-26 14:26:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
