新冠病毒核衣殼蛋白結構與功能的生物信息學分析及原核表達①
2022-02-10 09:51:02徐本錦李卓禧陳曉聰吳惠文玲楊婭男湯文婷寇妍祺余駿驍山西醫科大學汾陽學院醫學檢驗系汾陽032200
中國免疫學雜志 2022年24期
徐本錦 范 蕾 杜 淼 宣 焱 李 璟 李卓禧 陳曉聰 吳惠文 門 杰 劉 玲楊婭男 湯文婷 寇妍祺 余駿驍 (山西醫科大學汾陽學院醫學檢驗系,汾陽 032200)
2019年12 月,武漢暴發了新冠病毒肺炎疫情,患者出現發熱、胸悶、呼吸急促等癥狀,重癥患者因急性呼吸窘迫綜合征(acute respiratory distress syndrome,ARDS)、多器官功能衰竭而死亡[1-2]。該病毒是20世紀以來第3種可感染人類的高致病性病原[3]。2020年2月11日,國際病毒分類委員會將其命名為“SARS-CoV-2”[4]。據WHO 報道,截至 2021年 8 月23日,全球已有211 730 035例確診病例。
SARS-CoV-2為單股正鏈RNA病毒,是目前已知的第七種冠狀病毒[5-6]。SARS-CoV-2含有2個大的開放閱讀框(ORF1a和ORF1b),二者在不同冠狀病毒中高度保守,編碼4種同源結構蛋白:刺突蛋白(S)、包膜蛋白(E)、膜蛋白(M)、核衣殼蛋白(N)及8種輔助蛋白[1,4,6-9]。
N蛋白是SARS-CoV-2的重要組成,是一種高免疫原性蛋白,在感染過程中大量表達[10-12]。N蛋白具有較高的保守性和RNA分子伴侶活性,是干擾素拮抗劑和病毒編碼的RNA干擾抑制因子[13-14]。N蛋白與病毒基因組RNA相互纏繞形成核衣殼,在病毒RNA合成過程中發揮重要作用[15]。此外,N蛋白還參與病毒mRNA轉錄和復制,組織細胞骨架和免疫調節,調控細胞代謝和細胞周期,誘導感染后體液和細胞免疫應答[5-6,13,16-18]。因此,N蛋白可作為病毒檢測的標志性蛋白,已被廣泛用于疫苗研制和血清學檢測[19-20]。但目前有關SARS-CoV-2 N蛋白的報道較少,急需對該蛋白的更新認識。
本研究對SARS-CoV-2 N蛋白進行了系統生物信息學分析及原核表達,為闡明N蛋白在SARSCoV-2感染宿主細胞中的作用機制奠定了基礎,為靶向該蛋白的抗病毒藥物篩選提供了依據。
1 材料與方法
1.1 材料 pET-22b為本實驗室保存,蛋白和DNA marker購自寶生物公司;E.coli感受態細胞(Top10和BL21)、質粒提取及膠回收試劑盒購自全式金有限公司;XhoⅠ與NdeⅠ購自NEB;氨芐西林、IPTG、氯化鈉等購自國藥集團。
1.2 方法
1.2.1 表達載體pET-22b-N構建 酶切N基因片段和空載體pET-22b,回收酶切后的片段,鏈接載體與N基因片段后轉化感受態細胞Top10,將驗證后的重組載體轉化感受態細胞BL21,IPTG誘導蛋白表達。
1.2.2 N蛋白誘導表達 37 ℃、220 r/min培養11.5 h,采用終濃度為1 mmol/L的IPTG誘導,25 ℃、160 r/min繼續培養8 h,10%SDS-PAGE檢測誘導前后目的蛋白表達[21]。
1.2.3 生信分析 依據文獻中的網站對N蛋白的理化性質等特性進行分析[21]。
1.2.4 多序列比對與進化分析 UniProt網站(https://www.uniprot.org/blast/)下載與N蛋白序列相似度較高的12種病毒蛋白,分別用Clustal X2和MEGA7.0執行多序列比對和進化樹構建。
2 結果
2.1 理化性質 N蛋白是由419個氨基酸(共19種)組成的堿性蛋白,不含半胱氨酸。含負電荷氨基酸(Asp+Glu)36個,其中Asp 24個,Glu 12個;正電荷氨基酸(Arg+Lys+His)64個,其中Arg 29個,Lys 31個,His 4個。含量最多的為甘氨酸(10.30%),其次為丙氨酸和絲氨酸(均為8.80%,表1)。N蛋白分子量為45.62 kD,等電點為10.07,分子式為C1971H3137N607O629S7,消光系數為43 890 L/(mol·cm),不穩定指數為55.09;在哺乳動物細胞中的半衰期為30 h,在大腸桿菌體內>10 h,脂肪族系數為52.53,總平均親水系數為-0.971。
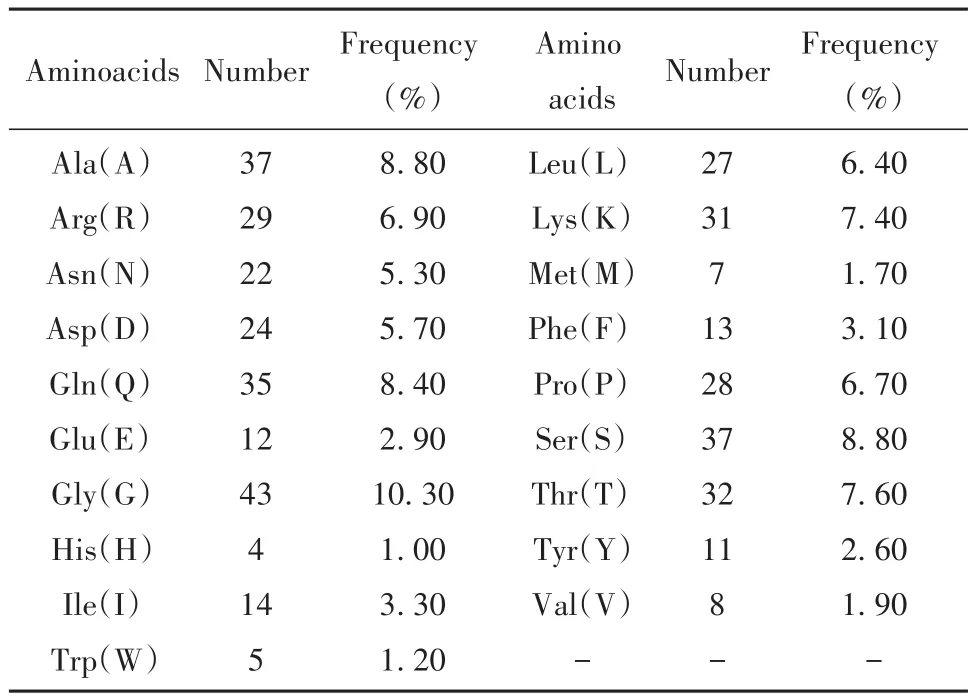
表1 新冠病毒N蛋白的氨基酸組成Tab.1 Amino acid composition of SARS-CoV-2 N protein
2.2 跨膜結構預測 預測結果顯示,SARS-CoV-2 N蛋白不存在跨膜螺旋區(圖1),不屬于跨膜蛋白。
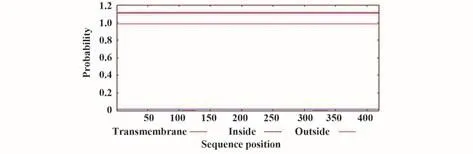
圖1 新冠病毒N蛋白的跨膜結構預測Fig.1 Transmembrane structure prediction of SARSCoV-2 N protein
2.3 親水/疏水性分析 親水/疏水性預測結果顯示,D371親水性最強,Score值為-3.556;A220和L221疏水性最強,Score值為2.322;親水殘基數遠多于疏水殘基數(圖2)。因此推測SARS-CoV-2 N蛋白為親水性蛋白。
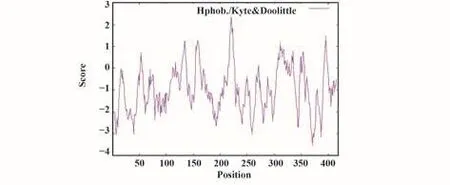
圖2 新冠病毒N蛋白親水/疏水性分析Fig.2 Hydrophobic/hydrophobic analysis of SARS-CoV-2 N protein
2.4 功能位點預測 SARS-CoV-2 N蛋白含有1個RNA結合結構域,位于N末端A50-G175位,該結構域包含12個RNA結合位點,分別為A50-A55、R107、Y109、Y111、R149、A156和E174。此外,C末端P258-A359位還存在1個二聚體界面,包含51個二聚體相互作用位點,分別為P258、Q260-A264、V270、F274、R277、G284-F286、L291、T296、W301、I304-Q306、A308-I320、M322、V324、G328-D341、N345、F346、L353和I357。
2.5 磷酸化修飾預測 結果顯示,SARS-CoV-2 N蛋白共有可能的磷酸化修飾57個,其中蘇氨酸位點22個,絲氨酸位點31個及酪氨酸位點4個(圖3)。上述位點及對應激酶如表2所示,通常取0.5為閾值,磷酸化勢能越高置信度越高。
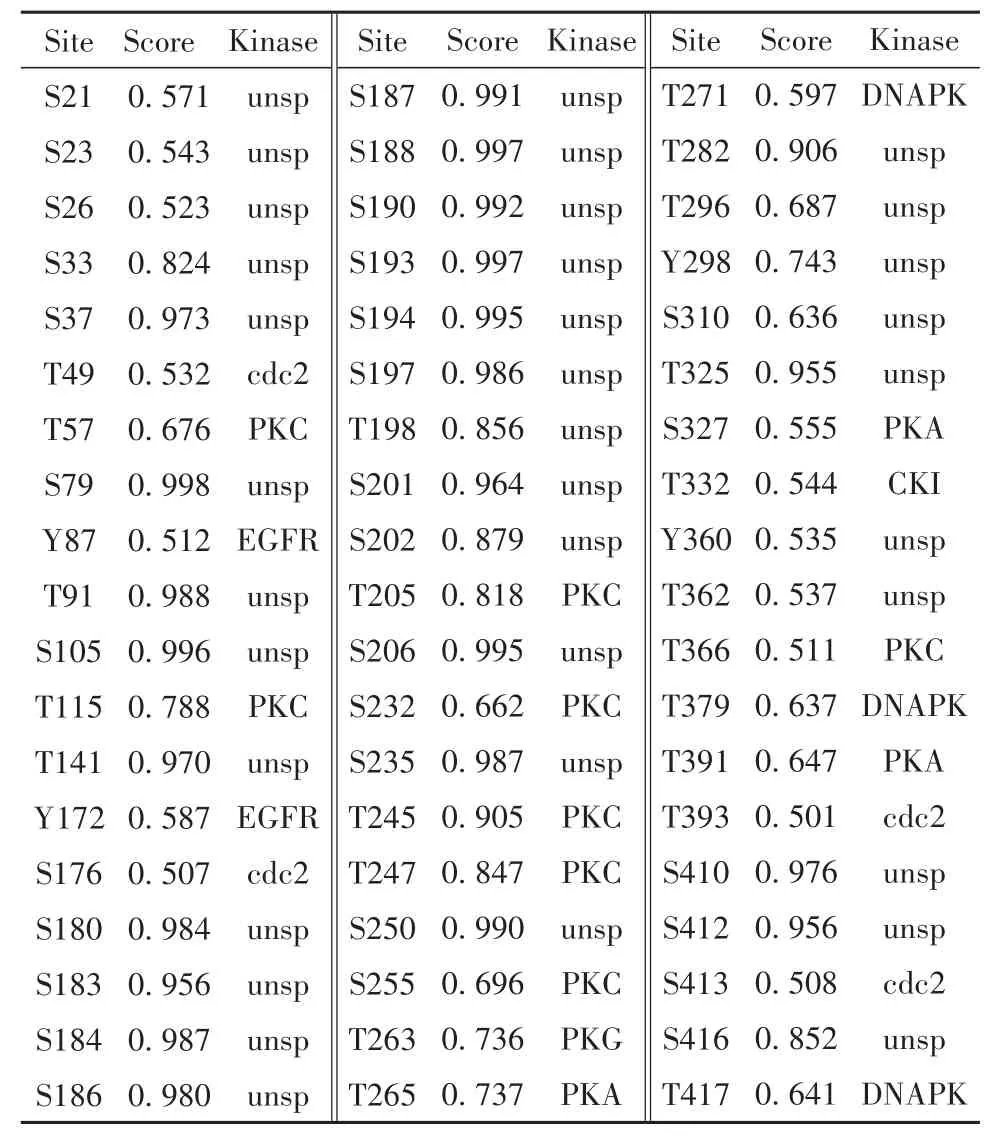
表2 N蛋白磷酸化修飾及對應激酶Tab.2 Phosphorylation sites of N protein and corresponding kinases
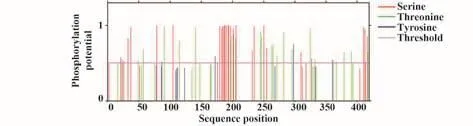
圖3 新冠病毒N蛋白磷酸化修飾Fig.3 Phosphorylation sites of SARS-CoV-2 N protein
2.6 糖基化修飾預測
2.6.1 N-糖基化修飾預測 結果顯示,N蛋白有2個潛在的N-糖基化位點,分別為N47和N269(圖4)。
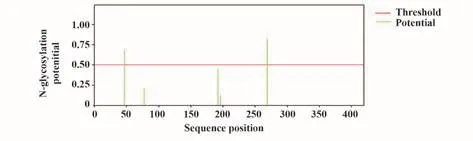
圖4 新冠病毒N蛋白N-糖基化修飾Fig.4 N-glycosylation site of SARS-CoV-2 N protein
2.6.2 O-糖基化修飾預測 結果顯示,N蛋白有23個潛在的O-糖基化位點,分別為S33、S176、S180、S183、S184、S186、S187、S188、S190、S193、S194、S197、T198、S201、S202、S206、T247、T271、T379、S410、S412、S413和T417(圖5)。
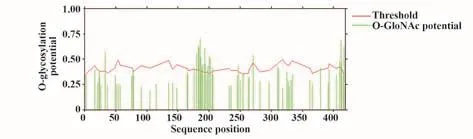
圖5 新冠病毒N蛋白O-糖基化修飾Fig.5 O-glycosylation site of SARS-CoV-2 N protein
2.7 信號肽預測 蛋白有無信號序列的判斷方法是通過預測N端前70個氨基酸中是否有潛在的酶切位點,結果顯示,N蛋白不含信號序列(圖6)。
圖6 新冠病毒N蛋白信號肽預測Fig.6 Prediction of SARS-CoV-2 N protein signal peptide
2.8 二級結構分析 結果顯示,N蛋白含有α-螺旋96個(22.20%),延伸鏈66個(16.47%),β-轉角34個(6.92%),無規則卷曲223個(54.42%)。見附圖1(www.immune99.com)。
2.9 三級結構分析 通過SWISS-MODEL數據庫將目的蛋白與已有蛋白進行序列比對,選擇相似度高或同源蛋白進行自動比對建模,預測未知蛋白的三級結構,SARS-CoV-2 N蛋白三級結構預測結果如圖7。
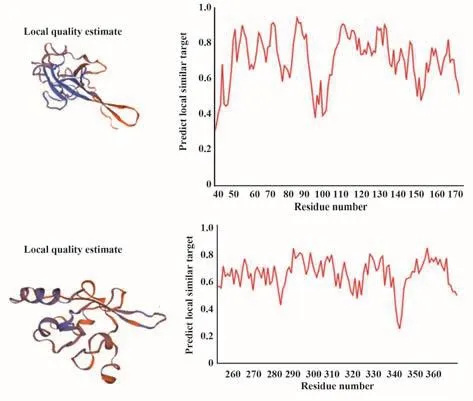
圖7 新冠病毒N蛋白三級結構預測及同源蛋白相似性波形圖Fig.7 Tertiary structure prediction of SARS-CoV-2 N protein and similarity waveform of its homologous proteins
2.10 抗原表位預測
2.10.1 T細胞抗原表位預測 IEDB預測結果顯示,N蛋白共有15個T細胞抗原表位,分別為G5-S21、F66-N77、D81-D98、K100-L113、G116-G120、K127-D128、I130-N140、A220-L230、K257-T271、Y298、H300-W301、A305-A336、D340、H356和 K361-K369(圖8)。
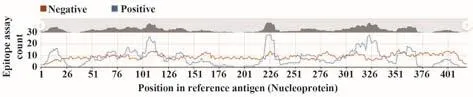
圖8 新冠病毒N蛋白T細胞抗原表位預測Fig.8 Prediction of T cell epitopes of SARS-CoV-2 N protein
2.10.2 B細胞抗原表位預測 IEDB預測結果顯示,取閾值為0.5,N蛋白共有11個線性B細胞抗原表位,分別為N4-I15、F17-N48、H59-S105、A119-K127、G137-Q163、T165-D216、R226-A267、R276-K299、D343-D348、D358-D402和S404-S416。根據表位分布圖最終確定N蛋白的優勢抗原表位區段為I15~D216(圖9)。
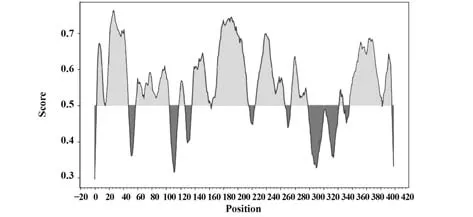
圖9 新冠病毒N蛋白B細胞抗原表位預測Fig.9 Prediction of B cell epitopes of SARS-CoV-2 N protein
2.11 相互作用網絡分析 N蛋白與6種蛋白存在二元相互作用,分別為70 kD熱休克蛋白1A、抗病毒天然免疫應答受體RIG-Ⅰ、信號轉導和轉錄激活因子1-α/β、信號轉導和轉錄激活因子2、蛋白酶體激活物復合體亞單位3和富含絲氨酸和精氨酸的蛋白特異性激酶1(圖10)。

圖10 新冠病毒N蛋白的二元相互作用分析Fig.10 Binary interaction analysis of SARS-CoV-2 N protein
2.12 同源性分析 UniProt網站下載與SARSCoV-2 N蛋白序列相似度較高12個N蛋白序列,分別為SARS冠狀病毒、SARS冠狀病毒PUMC03、SARS 冠 狀 病 毒 PUMC02、BtRs-β-冠 狀 病 毒/HuB2013、蝙蝠非典型冠狀病毒WIV1、蝙蝠冠狀病毒Rp/Shaanxi2011、蝙蝠冠狀病毒Rp3/2004、蝙蝠非典型冠狀病毒YNLF_31C、SARS冠狀病毒WH20、蝙蝠冠狀病毒Cp/Yunnan2011、蝙蝠冠狀病毒HKU3和蝙蝠冠狀病毒279/2005。多序列比對結果顯示,與SARS-CoV-2 N蛋白序列相似度最高的為BtRs-β-冠狀病毒/HuB2013和蝙蝠冠狀病毒Rp/Shaanxi2011(均為90.3%),其次為SARS冠狀病毒、SARS冠狀病毒PUMC02和SARS冠狀病毒PUMC03(均為89.8%)。相似度最低的是蝙蝠冠狀病毒HKU3(88.8%)。此外,13條序列中完全相同(*表示)的殘基有284個(67.8%);性質及其相似(:表示)和性質微弱相近(.表示)的殘基均為19個(4.5%),見附圖2(www.immune99.com)。
2.13 進化分析 采用MEGA7.0軟件對包括SARS-CoV-2在內的13種病毒N蛋白進行多序列比對并構建進化樹,結果顯示,SARS-CoV-2與蝙蝠非典型冠狀病毒YNLF_31C親緣關系最近,二者聚為一支,置信度為24,提示其可能具有共同祖先;其次,SARS冠狀病毒與蝙蝠非典型冠狀病毒WIV1聚為一支,置信度為20;蝙蝠冠狀病毒Cp/Yunnan2011與蝙蝠冠狀病毒HKU3聚為一支,置信度為49;BtRsβ-冠狀病毒/HuB2013與蝙蝠冠狀病毒Rp/Shaanxi 2011聚為一支,置信度為25;蝙蝠冠狀病毒Rp3/2004與蝙蝠冠狀病毒279/2005聚為一支,置信度為36(圖11)。

圖11 新冠病毒N蛋白的系統進化樹Fig.11 Phylogenetic tree of SARS-CoV-2 N protein
2.14 原核表達載體的構建
2.14.1 空載體的酶切驗證 pET-22b空載體圖譜如圖12A,大小約5 500 bp。經NdeⅠ、XhoⅠ單酶切及雙酶切后用瓊脂糖凝膠電泳檢測,結果顯示大小符合預期(圖12B)。
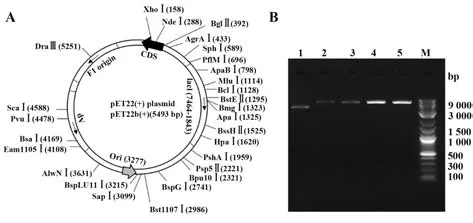
圖12 pET-22b空載體檢測及酶切驗證Fig.12 Detection and restriction endonuclease digestion of pET-22b empty vector
2.14.2 目的片段的酶切及PCR驗證 對N基因片段進行XhoⅠ與NdeⅠ雙酶切(圖13A);PCR擴增N蛋白編碼序列,電泳檢測顯示,條帶大小正確(圖13B)。
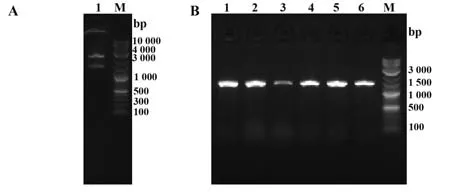
圖13 N蛋白基因片段的酶切及菌落PCR驗證Fig.13 Restriction endonuclease digestion and colony PCR validation of N protein gene fragment
2.14.3 重組質粒酶切驗證及轉化 將pET-22b-N進行雙酶切,電泳檢測結果顯示,大小正確(圖14A)。將pET-22b-N轉化Top10感受態細胞,在LB平板(Amp+)上于37 ℃培養12~13 h(圖14B左);將驗證后的重組質粒轉化BL21,37 ℃培養13 h(圖14B右)。
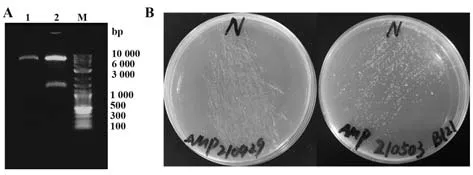
圖14 重組質粒pET-22b-N的酶切驗證與轉化Fig.14 Verification and transformation of recombinant plasmid pET-22b-N
2.15 蛋白誘導表達 挑取2個單菌落,IPTG誘導后10%聚丙烯酰胺凝膠電泳檢測,結果顯示:誘導后N蛋白表達含量比誘導前顯著增多(泳道4高于泳道1,泳道10高于泳道7)。為進一步優化N蛋白提純條件,分別對誘導前/后的全菌、上清、沉淀中N蛋白表達進行檢測,結果如圖15(黑色虛線框):誘導后的N蛋白大量表達于沉淀中(泳道6、12),因此在提純該蛋白時考慮從沉淀進行。
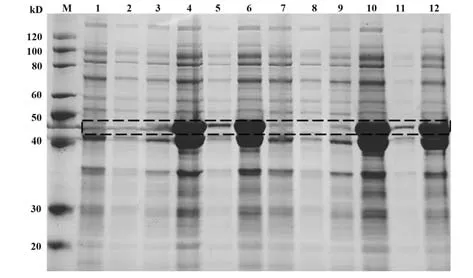
圖15 N蛋白誘導表達檢測Fig.15 Detection of induced expression of N protein
3 討論
N蛋白是病毒檢測的重要靶點和藥物作用靶標,保守性高,是SARS-CoV-2的重要抗原,參與病毒基因組包裝和病毒顆粒釋放[22-23]。血清學診斷發現,SARS患者血清中針對N蛋白的特異性抗體比SARS-CoV其他結構蛋白抗體具有更高的敏感性和持久性[24-25]。此外,N蛋白抗體在感染早期具有較高特異性[26]。因此,建立大量表達N蛋白的方法并對其進行系統生物信息學分析有助于深入了解該蛋白的功能和病毒RNA復制機制。
本研究顯示,SARS-CoV-2 N蛋白呈堿性,富含正電荷,有助于其與病毒基因組RNA結合[27-28]。N蛋白親水性較強,無信號肽,不屬于分泌型蛋白。二級結構分析發現,N蛋白以無規則卷曲為主(54.42%),與鐘琦等[29]報道一致,為抗病毒藥物研發與新冠肺炎患者診斷提供了參考。
抗原表位分析發現,N蛋白有15個T細胞表位和11個線性B細胞表位,其中I15-D216是其優勢B細胞表位。近期研究顯示,N蛋白存在較高頻率的R203K和G204R雙位點變異,該變異破壞了N蛋白整體結構穩定性和靈活性[30]。盡管N蛋白被認為是SARS-CoV-2疫苗開發和診療的重要靶點,但其持續進化的特點易產生傳染性更強的突變株,導致病毒與已有抗體親和力下降,給疫苗研發帶來巨大挑戰[3]。因此,需進一步進行免疫信息學分析,同時持續監測和追蹤SARS-CoV-2 N蛋白演變。
糖基化是重要的蛋白翻譯后修飾過程,高度親水的糖基對蛋白理化性質和生理功能具有重要影響。磷酸化是調節和控制蛋白活力和功能的最基本、最普遍,也是最重要的機制。本研究顯示,N蛋白有2個可能的N-糖基化修飾和23個O-糖基化修飾,有31個絲氨酸磷酸化修飾,22個蘇氨酸磷酸化修飾,以及4個酪氨酸磷酸化修飾。RAHMAN等[30]對N基因突變頻率較高的20例確診病例與感染率的關系進行了系統研究,發現N蛋白磷酸化修飾位點S250、S255、S310、S325、S327、T141、T247、T263、T265、T271和T362存在2~3種單點變異類型,分別為S250F/P、S255F/A/P、S310I/C/N、S325I/R/A、S327L/P、T141I/P/A、T247I/A、T263I/A、T265I/A、T271I/A 和T362I/K。此外,S193、S194、S201、S202、T205和S206等位點還存在不同形式的氨基端缺失變異類型。O-糖基化修飾位點S180是N蛋白突變形式最多的位點,具有6種氨基酸變異形式。以上變異可能影響SARS-CoV-2的傳播和侵染能力,但課題組并未發現N蛋白突變頻率與SARS-CoV-2感染率顯著相關。
N蛋白通過其N-末端結構域(N-NTD)與病毒RNA形成復合物,在病毒復制周期中發揮重要作用,使該結構域成為重要的藥物靶點[31-33]。本研究功能位點分析顯示,SARS-CoV-2 N蛋白含有1個RNA結合結構域,位于NTD A50-G175位,包含12個RNA結合位點,與KANG等[5]結果一致,發現N蛋白的N47-A50殘基具有高度靈活性且向外伸展,打開了RNA結合口袋,有助于同病毒基因組RNA高級結構結合。RAHMAN等[30]發現,N蛋白A55S、P67T、D81Y、A119V、P122L、D128Y、L139F和D144Y變異增加了NTD結構穩定性,而E62V、D103Y、A119S、A152S、A156S、L161F和P168S變異則降低了該結構穩定性。同樣,E62V、P67T、D144Y和P168S變異增加了N蛋白分子柔性,而A55S、D81Y、D103Y、A119S、A119V、P122L、D128Y、L139F、A152S、A156S和L161F變異則降低了RNA結合結構域分子柔性,進一步證明上述位點在功能上的重要性。本研究還顯示,CTD第P258-A359位存在1個二聚結構域,包含51個二聚相互作用位點。RAHMAN等[30]研究顯示,H300Y、T325I、S327L、T334I、D340N 改變增加了N蛋白穩定性,而Q289H、I292T、P344S、D348H改變則降低了分子穩定性。Q289H、I292T變異增加了分子靈活性,而H300Y、T325I、S327L、T334I、D340N、P344S和D348H則降低了分子柔韌性。提示這些結構域和功能位點在提高病毒轉錄和組裝效率方面起重要作用[34-35]。
多序列比對顯示,與SARS-CoV-2 N蛋白序列相似度最高的是BtRs-β-冠狀病毒/HuB2013和蝙蝠冠狀病毒Rp/Shaanxi2011(均為90.3%),其次是SARS冠狀病毒、SARS冠狀病毒PUMC02和SARS冠狀病毒PUMC03(均為89.8%)。此外,13條序列中完全相同的殘基占67.8%,提示N蛋白在進化上高度保守。進化分析顯示,SARS-CoV-2與蝙蝠非典型冠狀病毒YNLF_31C親緣關系最近,二者聚為一支,置信度為24,提示其可能有共同祖先。
N蛋白是SARS-CoV-2 IgM/IgG快速檢測試劑卡的核心原材料,目前已有多家公司在真核系統中完成了N蛋白表達。大腸桿菌原核表達系統以其安全性好、易放大培養、周期短等優點被廣泛用于基因工程藥物生產[36]。本研究構建了原核表達載體pET-22b-N,菌落PCR擴增和測序驗證結果顯示,載體構建正確。在大腸桿菌中表達N蛋白并研究其表達特性,結果顯示,經1 mmol/L IPTG誘導,N蛋白即可大量表達,且主要在沉淀中表達。從IgM/IgG快速檢測試劑卡成本考慮,大腸桿菌系統中N蛋白表達量高、速度快,將大大降低SARS-CoV-2篩查成本[37]。
本研究有助于全面揭示N蛋白的生物學功能,為開發和設計靶向N蛋白的快速診斷方法和抗病毒藥物提供了依據。