干旱脅迫對光叉委陵菜根系特征、結構和生理特性的影響
2022-02-15 02:48:48高鵬飛張靜范衛芳高冰郝宏娟吳建慧
草業學報 2022年2期
關鍵詞:植物
高鵬飛,張靜,范衛芳,高冰,郝宏娟,吳建慧
(東北林業大學園林學院,黑龍江 哈爾濱 150040)
干旱嚴重影響植物的生長與觀賞價值,甚至會導致植物的死亡。根系是植物吸收水分和養分的主要器官。有研究表明,經歷干旱后,植物根系總長度、總表面積、總體積、平均直徑和生物量等表型特征都會發生改變[1-2],根系細胞中的線粒體等細胞器的形態和數目也會出現變化[3]。干旱條件下,丙二醛(malondialdehyde,MDA)會在細胞中不斷積累,損傷生物膜系統[4-5],而細胞中的脯氨酸、可溶性蛋白等滲透調節物質含量會增加,提高細胞的保水能力[6-7],一些抗氧化酶如超氧化物歧化酶(superoxide dismutase,SOD)等也會增加活性,幫助植物抵御干旱脅迫[8-12]。因此,這些都可以作為判斷植物對干旱脅迫程度耐受性的重要指標。
東北地區因常年降水較少,在園林植被的選擇上具有一定的特殊性。光叉委陵菜(Potentilla bifurcavar.glabrata)是薔薇科(Rosaceae)委陵菜屬(Potentilla)多年生草本植物,能夠適應東北地區干燥的氣候條件,株型優美,花色美觀,根系發達,耐粗放管理,可以作為優良地被植物在園林中應用[13]。目前,對于委陵菜屬植物的研究多集中于資源開發利用、藥用和觀賞價值、引種馴化及花粉、葉表皮形態等[14-17],在生理方面尤其是干旱生理的研究較少。根系直接接觸土壤,當植物遭受干旱脅迫時,發達的根系是植物獲得水分和養分的重要保證。本研究通過自然控水的方法,測定干旱脅迫下光叉委陵菜的根系表型、結構特征以及根系生理指標變化,探討光叉委陵菜的耐旱能力臨界值,可以擴大其應用范圍,也可以為耐旱品種的選擇和城市綠化植物品種的豐富提供理論依據。
1 材料與方法
1.1 試驗材料
以東北林業大學園林學院苗圃地的光叉委陵菜多年生實生幼苗為研究對象,選取株高5 cm左右、生長周期一致的植株苗,緩苗處理2周待其生長。將生長正常且高度在15 cm左右的植株移植到直徑15 cm的花盆里,培養基質比例為營養土∶蛭石=3∶1,在植物培養室內進行種植,培養室溫度為25℃/18℃(晝/夜),光照強度為4000 lx,時間為16 h/8 h(光/暗)。
1.2 試驗方法
2019年4月在東北林業大學園林學院植物綜合實驗室中進行試驗。干旱脅迫處理前植株正常澆水。試驗設定對照組和脅迫組,對照組每天正常澆水,脅迫組分為兩組,其中一組在干旱脅迫0,5,10,15,20,25 d時分別取樣測定根系表型特征及生理指標,在0,5,15,25 d時分別取樣觀察根系解剖結構和超微結構,另一組在干旱脅迫5,10,15,20,25 d時進行復水處理,并在復水5 d后對植物材料進行取樣,以測定各項生理指標。每個測定取樣3株(3次重復)植株。所取待測樣品放入液氮后保存于-80℃超低溫冰箱。
其中,土壤水分設置包括:自然狀態(田間最大持水量的70%~80%)、輕度干旱脅迫(田間最大持水量的60%~70%)、中度干旱脅迫(田間最大持水量的50%~60%)、重度干旱脅迫(田間最大持水量的40%~50%)[18-19]。根據預先進行的土壤持水量測定,自然失水狀態下的土壤在干旱0 d為自然狀態,5、10 d為輕度干旱脅迫,15、20 d為中度干旱脅迫,25 d為重度干旱脅迫。
1.3 測定項目與方法
1.3.1 根系表型特征的測定 取植株根系洗去多余的土與雜質,用Expression 11000 XL根系掃描儀掃描整個根系,用WinRHIZO軟件分析根系的總長、總表面積、總體積及根系直徑的數據。參照陳麗飛[20]、襲梅[21]的方法,剪取根系洗凈吸干水分,105℃烘干2 h,65℃下烘干48 h以上至恒重,測量其干重。
1.3.2 根系超微結構的觀察 取植株根系距根尖1 cm以上的部位裝在瓶中,取材大小為1 mm×3 mm,抽真空加入3%的戊二醛固定24 h以上,0.1 mol·L-1磷酸緩沖液沖洗3次,用1%鋨酸固定4 h,去離子水沖洗3次,再用梯度濃度乙醇進行脫水處理,丙酮過渡,用環氧樹脂812進行滲透包埋,一段時間后用LKB-5切片機(LKB Co,Switzerland)切片,醋酸雙氧鈾-檸檬酸鉛雙重染色。用H7650透射電鏡觀察。
1.3.3 根系生理指標的測定 參照王學奎[22]的方法,測定MDA、脯氨酸、可溶性蛋白的含量及SOD活性。
1.3.4 圖片與數據處理 用Photoshop軟件進行圖片組合,將測定的試驗數據采用Excel 2007軟件進行計算和制表,用SPSS 22.0進行方差分析,ANOVA檢驗差異顯著性。
2 結果與分析
2.1 干旱脅迫對光叉委陵菜根系表型特征的影響
光叉委陵菜的根系總長、根系體積、根系總表面積及根系平均直徑隨著干旱脅迫時間的延長逐漸增加,最大值均出現在25 d時,其中,5~15 d時增長速度較快,20、25 d時增長趨勢變緩。說明植物的根系在輕度和中度的干旱脅迫時能正常生長,在重度的干旱脅迫時植物的根系生長受到抑制。25 d時的根系總長、總表面積、總體積和平均直徑較0 d分別增加了118.45%、34.44%、80.00%、64.10%。根系干重在干旱前期大幅增加,在15 d時達最大值,最大變化率為38.30%,而在干旱后期則迅速減少,且低于0 d的干重(表1)。
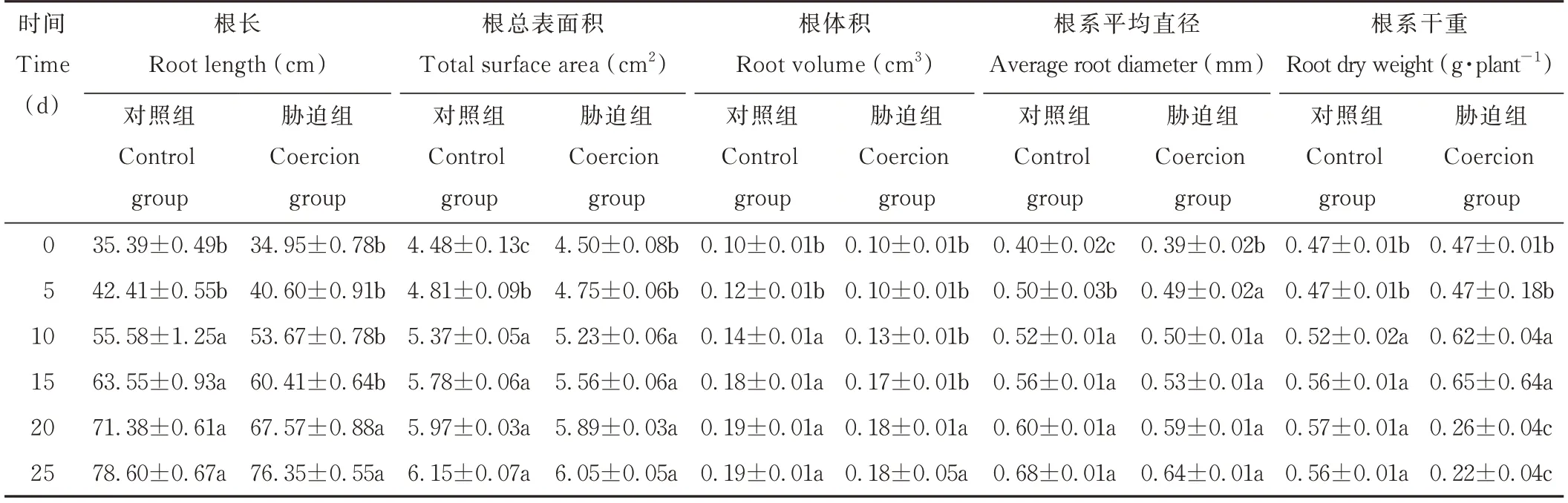
表1 干旱脅迫對光叉委陵菜根系表型特征的影響Table 1 Effect of drought stress on root characteristics of P.bifurca var.glabrata
2.2 干旱脅迫對光叉委陵菜根系超微結構的影響
0 d自然狀態下(圖1A),光叉委陵菜正常生長,根系的細胞結構完整,細胞壁較厚且完整;線粒體多為圓形或長橢圓形,沿細胞壁分布,內嵴結構明顯;此時細胞中不存在淀粉粒。5 d輕度脅迫下(圖1B),細胞壁較正常狀態下變薄;線粒體數量呈下降趨勢,形狀與0 d自然狀態下一致,內嵴結構明顯;淀粉粒出現,數量較多,充滿在細胞內。15 d中度脅迫下(圖1C),細胞壁厚度進一步變薄;線粒體明顯縮小,內嵴結構不明顯;淀粉粒的數量減少,但是體積開始變大,分布在細胞中。25 d重度脅迫下(圖1D),細胞壁變得更薄,并出現損壞的現象;線粒體消失;淀粉粒的體積與15 d中度脅迫時相比更大。液泡在光叉委陵菜的根系結構中始終沒有出現,這可能與植物的品種或者與植物的生長周期和狀態有關。
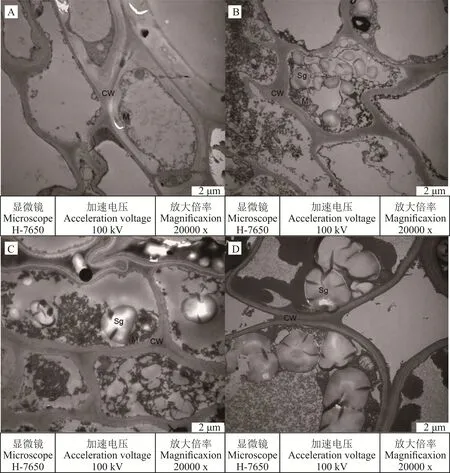
圖1 干旱脅迫對光叉委陵菜根系超微結構的影響Fig.1 Effect of drought stress on root ultrastructure of P.bifurca var.glabrata
由表2可以看出,細胞壁的厚度呈現逐漸減小的趨勢,5 d輕度脅迫、15 d中度脅迫和25 d重度脅迫下的厚度相較于前一脅迫程度分別減少了57.14%、8.33%、9.09%,其中5 d輕度脅迫相較于0 d自然狀態的變化程度最大,兩種狀態下的細胞壁厚度存在顯著差異(P<0.05)。對照時沒有淀粉粒的存在,5 d輕度脅迫時淀粉粒出現,其個數在15 d中度脅迫時顯著降低(P<0.05),25 d重度脅迫下淀粉粒數量較15 d中度脅迫時增加但不顯著(P>0.05),但淀粉粒平均直徑一直顯著增加(P<0.05)。線粒體的個數顯著減少,25 d重度脅迫時線粒體消失(P<0.05)。
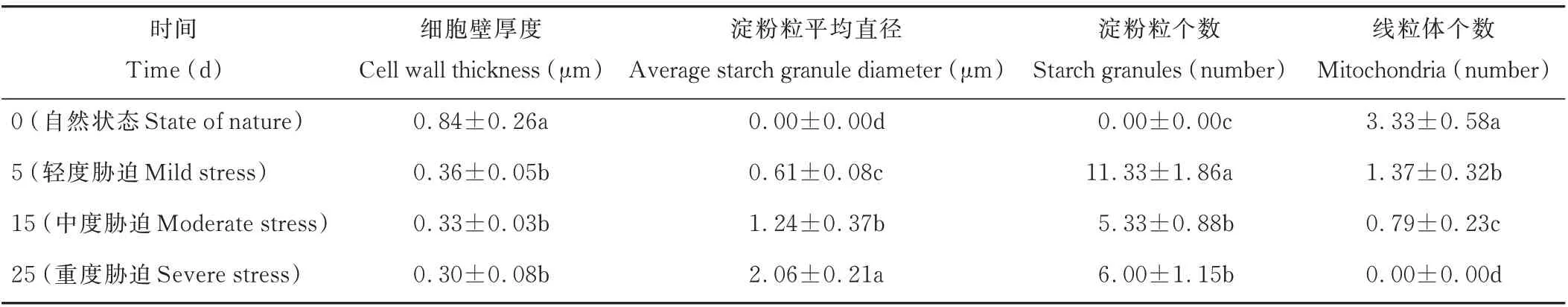
表2 干旱脅迫對光叉委陵菜超微結構細胞器的影響Table 2 Effect of drought stress on ultrastructure organelles of P.bifurca var.glabrata
2.3 干旱脅迫對光叉委陵菜根系生理指標的影響
2.3.1 干旱脅迫對光叉委陵菜根系MDA含量的影響 由圖2A可知,隨著干旱脅迫時間的延長,光叉委陵菜體內MDA含量持續顯著增長(P<0.05),5、10、15、20、25 d較0 d分別增長了35.90%、121.99%、152.84%、172.03%、286.30%。說明在干旱脅迫前期,植株抗旱能力較強,能夠有效地抑制MDA的產生,但后期,MDA含量急劇增長,可能是由于干旱脅迫程度的加深破壞了植株的干旱脅迫調節系統。復水后,同一脅迫時間下的MDA含量均有顯著下降(P<0.05),5、10、15、20、25 d脅迫時的MDA含量分別下降了22.70%、34.96%、29.59%、23.26%、15.38%,下降后MDA含量整體水平仍較高。25 d復水后,MDA含量仍保持在很高的水平,說明重度干旱脅迫對植株造成了較為嚴重的傷害。
2.3.2 干旱脅迫對光叉委陵菜根系脯氨酸含量的影響 如圖2B所示,當干旱時間延長時,光叉委陵菜根系脯氨酸含量呈增加的趨勢。干旱脅迫0~15 d期間,脯氨酸含量增長速度緩慢,不同天數的含量差異不顯著(P>0.05),但當脅迫時間超過15 d后,脯氨酸含量增長速度加快,含量差異顯著(P<0.05)。5、10、15、20、25 d較0 d分別增長了5.43%、8.93%、10.77%、61.62%、181.27%。復水使脅迫程度減輕,5、10、15、20、25 d復水后的脯氨酸含量比脅迫時的含量分別下降了0.12%、2.12%、2.22%、9.41%、1.52%,但變化都不顯著(P>0.05)。25 d復水后,脯氨酸含量仍保持在很高的水平,與復水前相比僅下降1.52%,說明在重度干旱脅迫下,植株受傷害程度較高,但其根部仍沒有死亡。
2.3.3 干旱脅迫對光叉委陵菜根系可溶性蛋白含量的影響 由圖2C可知,光叉委陵菜可溶性蛋白含量隨干旱脅迫時間的延長呈現出先上升后下降的趨勢。可溶性蛋白含量在0~10 d顯著升高(P<0.05),并在15 d達到最大值,20~25 d顯著降低(P<0.05),但仍高于0 d的含量。5、10、15、20、25 d較0 d分別增長了47.29%、69.27%、75.87%、62.56%、53.02%。這表明光叉委陵菜在應對干旱脅迫時可以產生大量的可溶性蛋白來調節細胞滲透壓,但是超過一定程度的干旱脅迫可能會對植物細胞造成損傷,使細胞內可溶性蛋白含量降低。復水使脅迫程度減輕,5、10、15、20、25 d復水后的可溶性蛋白含量均比5、10、15、20、25 d脅迫時的可溶性蛋白含量低,分別下降了0.18%、9.74%、4.10%、10.74%、11.12%。20和25 d復水后,可溶性蛋白含量顯著下降(P<0.05),但含量仍較高,說明可溶性蛋白在植物緩解較嚴重的干旱脅迫時仍發揮著較大的作用。
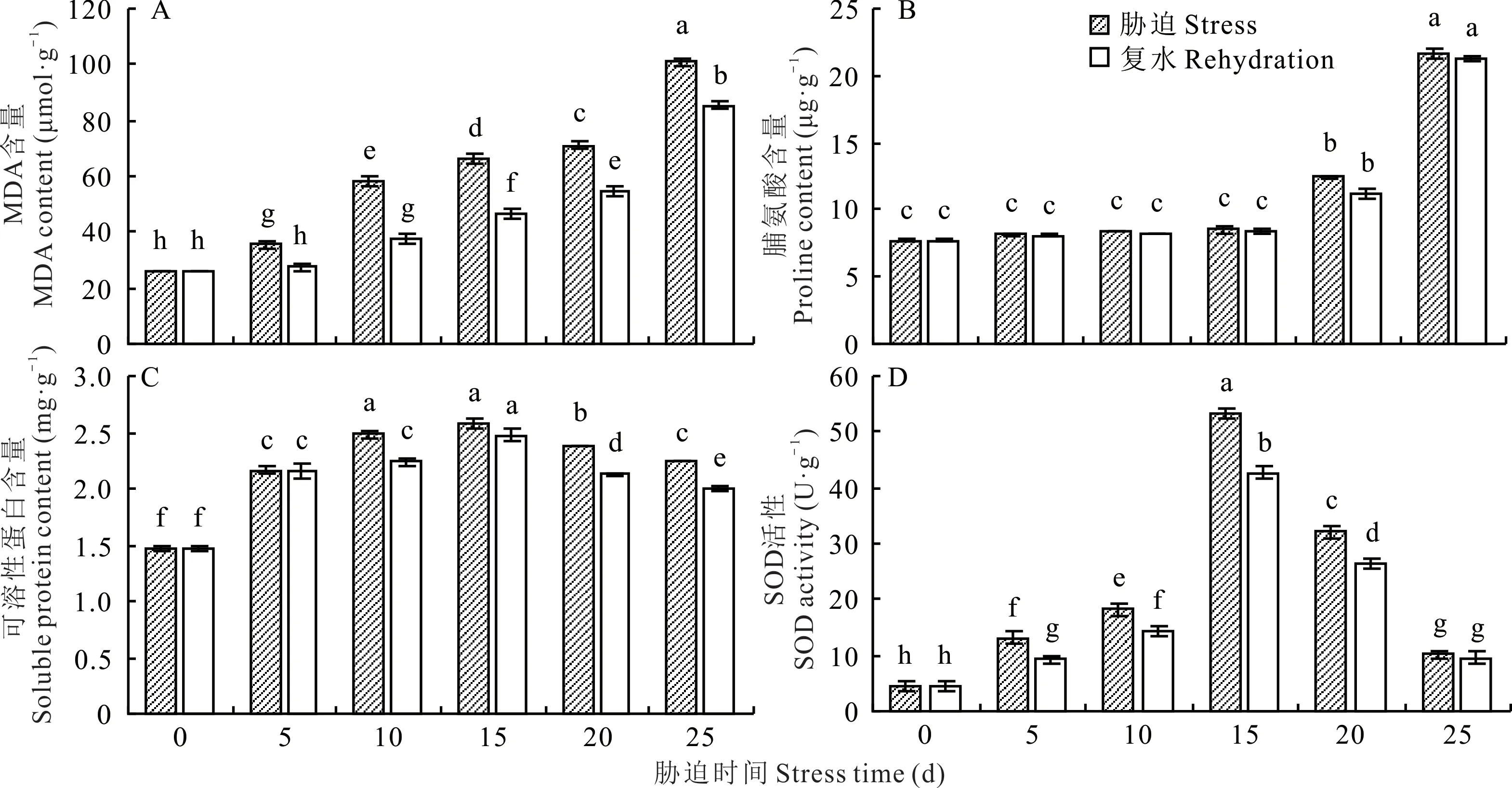
圖2 干旱脅迫對光叉委陵菜根系生理指標的影響Fig.2 Effect of drought stress on physiological index of root system of P.bifurca var.glabrata
2.3.4 干旱脅迫對光叉委陵菜根系SOD活性的影響 由圖2D可知,隨著干旱處理的時間延長,光叉委陵菜SOD活性在不同的處理時間下均發生顯著改變(P<0.05),并呈先上升后下降的趨勢,在15 d時達到最大值。5、10、15、20、25 d的SOD活性均高于0 d,分別增長了183.61%、290.47%、1049.25%、590.19%、117.47%。說明SOD活性在光叉委陵菜遭受干旱脅迫時較為敏感。復水在一定程度上減輕了干旱對植物造成的損傷,不同干旱天數復水后,SOD活性均有所下降,5、10、15、20、25 d復水后的SOD活性比5、10、15、20、25 d脅迫時低,分別下降了29.44%、20.86%、20.22%、17.65%、6.12%。干旱5~20 d復水后SOD活性均有顯著下降(P<0.05),但脅迫25 d時的SOD活性已下降到很低的水平,且復水后僅下降6.12%,下降不顯著(P>0.05),說明重度干旱脅迫已對光叉委陵菜的SOD酶系統造成破壞。
3 討論
3.1 干旱脅迫對植物根系表型特征的影響
根系是土壤-植株-大氣水循環過程中的關鍵一環,在干旱的環境中,根系的適應能力尤為關鍵,植物的生長發育水平以及對外界環境的適應能力可以通過根系的結構特征來體現[23]。在遭受脅迫時,植物往往通過維持較大的根系長度、總表面積、總體積、平均直徑和調節生物量等方面來維持自身的水分吸收和運輸能力,提高自身抗旱性。根系長度是評價根系吸收能力的重要指標之一,較長的根系可以吸收更多的水分,使植物有效抵御干旱[24]。本研究發現,當干旱脅迫的時間延長時,光叉委陵菜的根系長度與對照組相比有所降低,說明根系生長受到了干旱的影響,而Wang等[25]發現,干旱脅迫顯著降低了高羊茅(Festuca elata)的根系總長度,這與本研究結果一致。Pirnajmedin等[26]研究認為根系的體積、干重、總表面積等與植物的抗旱能力呈正相關。本研究中,光叉委陵菜根系的總表面積、總體積以及根系平均直徑在干旱脅迫過程中呈現增加趨勢,且在整個脅迫過程中的增長量與對照組相比較少但差異不顯著,說明其根系具有一定的抗旱能力,能夠抵抗一定程度的干旱脅迫,在受到脅迫時仍然會進行生長過程。對植物根系的干重進行比較分析發現,光叉委陵菜的根系干重先增加后降低,這可能是由于較嚴重的水分脅迫影響了植物的正常生長,根系被破壞,也可能是植物把碳水化合物的分配重心轉移到了莖葉上,使植株能更好地進行光合作用,增強競爭力[27]。
3.2 干旱脅迫對植物根系結構的影響
植株超微結構的變化規律是判斷其抗旱能力的重要指標[28]。本研究中,隨著干旱脅迫加重,光叉委陵菜根系的細胞壁變薄,淀粉粒的數量先增加后減少但體積不斷增加,線粒體數量減少且結構不清晰直至完全消失,液泡始終未出現。對于線粒體的變化,本研究結果與衛星等[29]結果一致。有研究表明,隨著干旱程度的加重,線粒體會發生內嵴、外膜降解和內腔空洞化等一系列結構變化[30],這印證了本研究的結果。線粒體作為三羧酸循環和氧化磷酸化的主要場所,在細胞中消失,說明重度脅迫下根系細胞已經難以維持正常的生理功能,這或許導致了根系結構的擴散。有研究發現在干旱脅迫時,淀粉粒出現較少[31],原因可能是干旱脅迫使得類囊體膜結構發生了異常,從而降低了光合作用,導致光合產物減少,最終導致根系細胞淀粉粒數量減少[32]。但是在本研究中,隨著干旱脅迫程度的增加,淀粉粒的數量先增后減,淀粉粒的體積不斷地增加,這可能與植物材料本身有關,也可能是因為隨著干旱脅迫程度的增加光叉委陵菜的根系通過增加淀粉粒的數量來緩解能量的短缺,從而抵抗遭受的逆境脅迫。
3.3 干旱脅迫對植物根系生理特性的影響
植物處在干旱狀態下時,會產生大量的O2-和H2O2等具有強氧化能力的活性氧(reactive oxygen species,ROS),發生膜脂過氧化反應產生MDA,氧化膜的有序性、結構性會降低,膜透性增大,最終影響植物細胞的代謝[33]。在本研究中,干旱脅迫時間的不斷延長,加劇了植物體內的過氧化反應,光叉委陵菜根系的MDA含量不斷上升,嚴重的膜脂過氧化作用使生物膜系統遭到破壞,各類細胞器膜逐漸斷裂、分解,這可能是導致線粒體消失的主要原因之一。復水能夠顯著降低MDA在植物體內的含量,緩解膜脂過氧化,這和肖凡等[34]的研究結果一致。完整且有序的膜系統可以保證細胞中各個區域的生理生化反應能夠有條不紊地進行[35],而膜脂過氧化作用導致的滲漏會破壞這種代謝分區,從而釋放或激活大量的水解酶,降低植物的抗旱性[36],因此隨干旱脅迫時間的延長,復水前后的MDA含量差值逐漸減小。
干旱脅迫下,植物會增加細胞內滲透調節物質,以維持細胞內的膨壓,保證植物進行正常生理代謝活動[37]。脯氨酸和可溶性蛋白作為理想的滲透調節物質,可以通過主動積累來降低細胞滲透勢[38]。干旱脅迫前期,可能是因為光叉委陵菜抗氧化酶作用較大,能有效地清除細胞中的ROS,降低膜脂過氧化作用對細胞造成的影響,因此脯氨酸含量僅有略微升高,而干旱15 d后,較嚴重的干旱脅迫破壞了植物的抗氧化酶系統,此時脯氨酸迅速大量積累以調節細胞滲透壓,緩解干旱對植物的傷害,李鵬等[39]的研究中也得出了類似的結論。植物在逆境脅迫下,會將體內的不溶性蛋白轉化為可溶性蛋白,增強滲透調節能力[40]。本研究中,可溶性蛋白含量的最大值出現在15 d,呈現先上升后下降的趨勢,這與陳明濤等[41]的研究結果一致。脅迫前期蛋白質含量增加,可能是由于膜損傷釋放出了蛋白質,蛋白酶活性升高使蛋白質水解,而水解所得的氨基酸又用于合成如抗氧化酶和脫水蛋白等響應干旱脅迫的蛋白質,導致光叉委陵菜根系內可溶性蛋白含量進一步提高[42]。而在脅迫后期,較嚴重的干旱脅迫可能抑制了蛋白質的合成并加速了蛋白質的降解,使可溶性蛋白含量降低[43]。
一般研究認為,抗氧化酶的活性與植物的抗旱性密切相關[44-45]。SOD是植物體內的關鍵抗氧化酶,可以降低膜脂過氧化作用,有效清除ROS,從而保護膜結構,通常利用其活性變化情況來反映植物的逆境適應能力[46]。干旱脅迫前期,植物體內的SOD活性逐漸增加,增強了細胞清除ROS的能力,但隨著脅迫繼續進行,不斷積累的ROS加劇了膜脂的過氧化程度,細胞有限的抗氧化能力使得ROS的形成和清除之間的動態平衡被打破,細胞內環境逐漸紊亂,抗氧化酶系統受到影響,SOD的活性隨脅迫時間的延長逐漸下降。季楊等[47]的研究也得出了類似的結果。復水后,植物的干旱情況得到緩解,細胞內的膜脂過氧化作用減弱,因此SOD活性均較復水前有所降低。結果表明,光叉委陵菜的抗氧化防御系統具有一定的潛力去應對重度干旱脅迫對其造成的傷害。
4 結論
干旱脅迫對光叉委陵菜根系總長度、總表面積、總體積和根系平均直徑的影響較小,但根系干重隨干旱脅迫時間的延長呈現波動趨勢,0~15 d上升,20~25 d顯著下降(P<0.05)。
當干旱脅迫時間延長時,根系MDA的含量逐漸增加,復水后含量降低顯著(P<0.05);根系脯氨酸的含量一直上升,20~25 d顯著增加(P<0.05),復水后含量較復水前上升不顯著(P>0.05);可溶性蛋白的含量先升高后降低,20~25 d復水后含量較復水前上升顯著(P<0.05);SOD活性在0~15 d顯著上升(P<0.05),20~25 d顯著下降(P<0.05),復水后不同脅迫時間的SOD活性幾乎都顯著下降(P<0.05),但25 d復水后SOD活性基本無法恢復。
綜合以上結果,光叉委陵菜能耐20 d的自然干旱,具有較強的抗旱性,可以作為東北地區的抗旱植物。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
