藏北高寒草地土壤真菌群落系統發育多樣性對放牧的響應
2022-02-15 03:33:06張豪睿
草地學報 2022年1期
張豪睿, 付 剛
(1.中國科學院地理科學與資源研究所生態系統網絡觀測與模擬重點實驗室拉薩高原生態系統研究站, 北京 100101;2.中國科學院大學, 北京 100049)
生物多樣性包括物種多樣性和系統發育多樣性等,是人類可持續發展的重要基礎[1]。土壤真菌作為與植物多樣性和生產力密切相關的土壤微生物,對調控植物多樣性和生產力有著十分重要的影響[2-3]。土壤真菌多樣性作為衡量土壤生態狀況的重要生物指標,不僅能夠反映陸地生態系統質量,而且可以對環境脅迫做出快速反應[4]。草地作為一種非常重要的陸地生態系統,放牧是其最重要的人類利用方式之一。因此,量化草地生態系統土壤真菌多樣性對放牧的響應規律對于草地生態系統放牧優化管理和生物多樣性保護等都非常重要。
青藏高原作為“世界第三極”,對氣候變化和人類活動的干擾尤為敏感[5]。青藏高原是歐亞大陸最大的草原單元,面積約為250萬平方公里,且孕育了多種不同類型的高寒草地,如高寒草甸和高寒草原化草甸等[6-7]。青藏高原豐富的草地資源使得放牧成為青藏高原高寒草地最重要的土地利用方式[8]。由于氣候條件和土壤條件等原因,青藏高原上形成了多種不同方式的放牧活動,如冷季放牧和暖季放牧等,從而形成了冷季牧場和暖季牧場等的區分。放牧活動將持續影響青藏高原草地生態環境及其生態系統服務功能,從而對野生動物保護(尤其是珍稀瀕危野生動物)和人類健康發展等產生重要影響。放牧不僅可以通過牲畜踩踏草地,改變高寒草地的土壤緊實度、透氣性和持水能力等物理性質[9],而且放牧所產生的牲畜糞便可以改變土壤的營養成分等化學性質[10]。放牧造成的土壤理化性質的變化則可以影響土壤真菌多樣性。有關放牧對青藏高原高寒草地的土壤真菌多樣性影響的研究已經開展。然而目前的研究并未得到一致的結論,如Che等[11]在青藏高原的放牧試驗研究表明,放牧沒有顯著改變土壤真菌群落α多樣性和群落組成。Yang等[12]在青藏高原一個高寒草甸的放牧試驗研究則表明冷季放牧和全年放牧均未顯著改變土壤真菌群落α多樣性,卻都改變了土壤真菌群落結構。此外,前人的研究主要關注土壤真菌群落物種多樣性,而對土壤真菌群落系統發育多樣性的研究較少。然而僅分析物種或系統發育多樣性不能完全解決生理限制、生物相互作用、地理障礙和進化過程如何相互影響當地群落結構的問題[13-15]。群落系統發育結構模式不僅反映了基于生態位的過程,如競爭和棲息地過濾,還反映了物種進化、生態差異和生物地理過程[16-18]。
因此,放牧是否會影響青藏高原高寒草地土壤真菌群落系統發育多樣性以及是否會改變土壤真菌群落系統發育多樣性沿海拔的分布格局有待進一步研究;放牧對青藏高原高寒草地土壤真菌群落系統發育多樣性的影響是否與放牧季節和草地類型有關也有待于進一步研究。基于此,本研究依托2008年在藏北高原當雄縣設置的3塊配對的圍欄和自由放牧地,進行以下研究,(1)比較了冷季放牧和暖季放牧對土壤真菌群落系統發育多樣性的影響差異;(2)高寒草原化草甸和高寒草甸的土壤真菌群落系統發育多樣性對暖季放牧的響應差異;(3)對比分析了對照和放牧條件下的土壤真菌群落系統發育多樣性的海拔分布格局的差異。本研究的開展能夠為全球變化背景下的青藏高原土壤微生物多樣性保護和高寒草地生態系統優化放牧管理提供理論依據。
1 材料與方法
1.1 研究區域和試驗設計
研究區域位于西藏自治區拉薩市當雄縣。放牧是藏北高原重要的土地利用方式,為量化放牧對藏北高寒草地生態系統的影響,2008年設置了3個配對的圍欄和自由放牧試驗樣地(圖1),每個圍欄樣地約為20 m × 20 m。三塊樣地分別位于4 313 m(30°30′ N,91°04′ E),4 513 m(30°31′ N,91°04′E)和4 693 m(30°32′ N,91°03′ E)。這3個樣地包括冷季放牧的高寒草原化草甸(Alpine steppe meadow for winter pasture,ASMWP)、暖季放牧的高寒草原化草甸(Alpine steppe meadow for summer pasture,ASMSP)和暖季放牧的高寒草甸(Alpine meadow for summer pasture,AMSP)。圍欄內為封育樣地,禁止放牧。圍欄外為自由放牧樣地,放牧牲畜主要為牛羊等。ASMWP和ASMSP樣地的優勢物種分別為小嵩草(Kobresiapygmaean)、黒褐苔草(Carexatrofusca)和絲穎針茅(Stipacapillacea),而AMSP樣地的優勢種為小嵩草。三個樣地的土壤質地均為砂壤土,屬于高寒草甸土。在0~30 cm土壤深度內,ASMWP、ASMSP和AMSP的土壤全氮和有機碳分別為2.1 g·kg-1和19.8 g·kg-1,2.2 g·kg-1和24.0 g·kg-1,3.3 g·kg-1和43.7 g·kg-1[19]。為了方便、簡潔的表述兩個樣地間的群落周轉關系,我們利用“CC”來表示兩個對照樣地間的系統發育距離,C3C5代表對照條件下ASMWP和ASMSP間的系統發育距離;C3C7代表對照條件下ASMWP和AMSP間的系統發育距離;C5C7代表對照條件下ASMSP和AMSP間的系統發育距離。利用“GG”表示兩個放牧樣地間的系統發育距離,G3G5代表放牧條件下ASMWP和ASMSP間的系統發育距離;G3G7代表放牧條件下ASMWP和AMSP間的系統發育距離;G5G7代表放牧條件下ASMSP和AMSP間的系統發育距離。C3G3,C5G5以及C7G7分別表示高寒草原化草甸冷季牧場、高寒草原化草甸暖季牧場和高寒草甸暖季牧場的圍欄和放牧處理間的系統發育距離。C_ASMWP和G_ASMWP分別表示圍欄和冷季放牧處理下的高寒草原化草甸樣地;C_ASMSP和G_ASMSP分別表示圍欄和暖季放牧處理下的高寒草原化草甸樣地;C_AMSP和G_AMSP則分別表示圍欄和暖季放牧處理下的高寒草甸樣地。
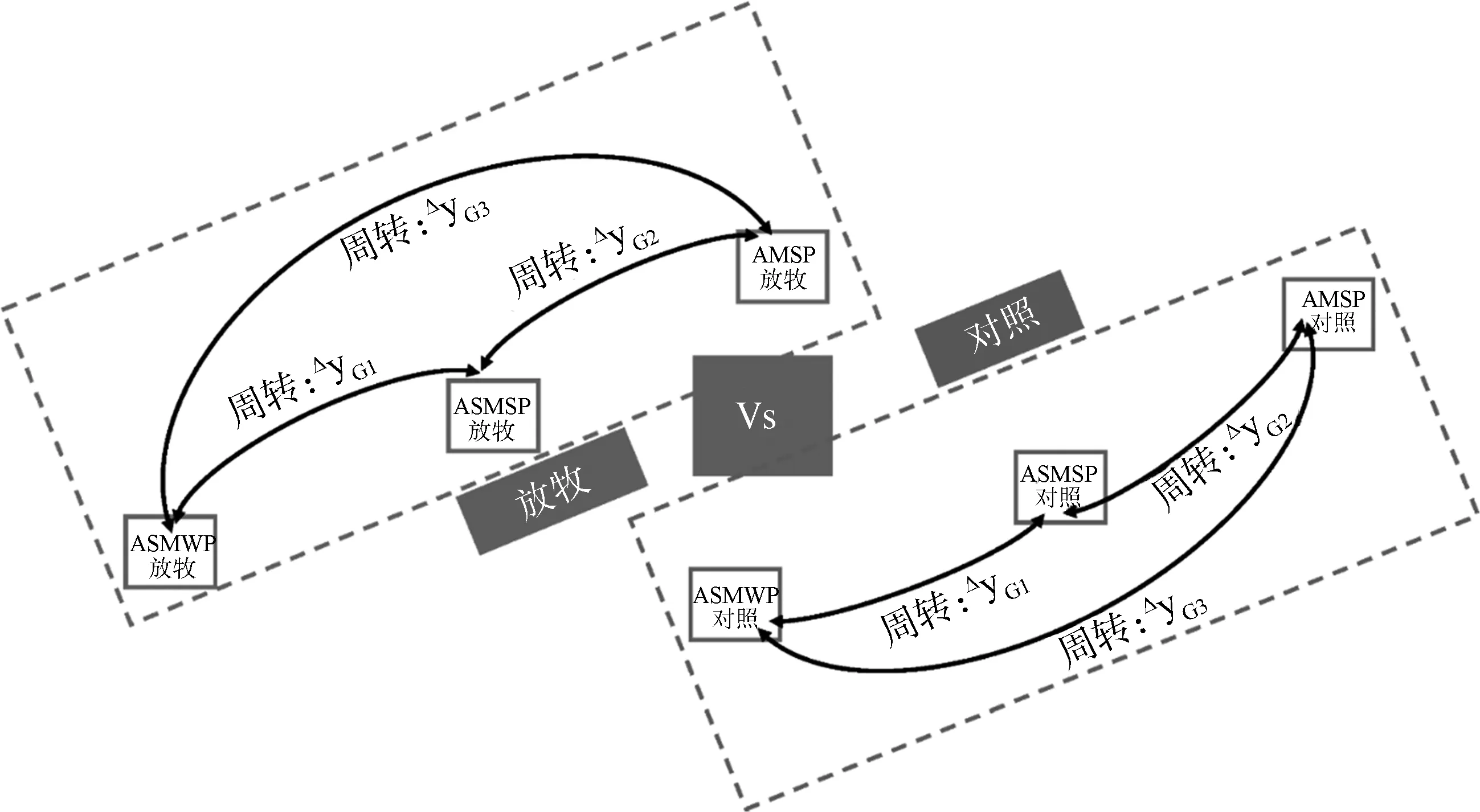
圖1 放牧樣地示意圖Fig.1 Schematic diagram of grazing plots注:ASMWP,冷季放牧的高寒草原化草甸;ASMSP,暖季放牧的高寒草原化草甸;AMSP,暖季放牧的高寒草甸Note:ASMWP,alpine steppe meadow for winter grazing;ASMSP,alpine steppe meadow for summer pasture;AMSP,alpine meadow for summer pasture
1.2 土壤樣品的采集和分析
在2018年8月,在每個海拔的圍欄外(自由放牧樣地)和圍欄內(圍欄封育樣地)各選取4個樣方(50 cm×50 cm)。在每個樣方內,使用直徑為3.7 cm的土鉆收集表層土壤(0~10 cm)。在每個重復樣方內采集3鉆土樣充分混勻后作為一個重復樣本。4個重復樣本不進行混合。一部分土壤樣品立即放入液氮中,帶回實驗室放置在-80℃的冰箱里保存,用于土壤微生物的測試分析。將其余的土壤樣品放置在-20℃的冰箱里保存,用于土壤常規理化性質(土壤含水量、銨態氮、硝態氮、速效磷和pH值)的觀測,其中土壤含水量采用烘干法觀測,銨態氮和硝態氮采用流動分析儀觀測[20]。
土壤真菌群落采用高通量測序,測序公司為廣州賽哲生物科技股份有限公司。樣品使用QIAGEN(Dusseldorf,Germany)試劑盒提取并純化DNA。用ITS1F和ITS2R引物擴增土壤真菌群落的ITS1F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)和ITS2R(5′-GCTGCGTTCTTCATCGAT GC-3′)。PCR反應體系為20 μL。PCR反應擴增條件為:98℃預變性1分鐘;27個循環進行聚合酶鏈反應擴增;最后一次擴增為72℃條件下10 分鐘。獲得的PCR產物送往Illumina HiSeq 2500平臺進行測序。詳細的DNA提取方法以及生物信息學分析詳見[7]。篩選OTUs中出現頻數最高的序列作為OTUs代表序列。用RDP Classifier方法與UNITE數據庫對土壤真菌OTUs代表序列進行物種注釋分析[17]。
1.3 統計分析
系統發育多樣性(Phylogenetic diversity,PD)、平均進化距離(Mean pairwise distance,MPD)以及平均最近分類距離(Mean nearest taxon distance,MNTD)分別由“picante”包的“PD”、“MPD”和“MNTD”計算得到(https://github.com/skembel/picante)。βMNTD是兩個不同群落中的最近親屬間的豐度加權平均系統發育距離,本研究中使用“iCAMP”包的“bmntd.big”函數得到。使用“vegan”包的“adonis2”函數進行多元置換方差分析(Permutational multivariate analysis,PARMANOVA)統計分析,以檢驗放牧和對照處理間的βMNTD的顯著性(https://github.com/vegandevs/vegan/)。使用“vegan”包的“monoMDS”函數對βMNTD進行非計量多維標度(Nonmetric multidimensional scale analysis,NMDS)分析(https://github.com/vegandevs/vegan/)。采用T檢驗統計分析了每個樣地間放牧處理和對照處理的PD,MPD以及MNTD的差異。對每兩個樣地間的環境因子的歐氏距離也進行了樣本T檢驗。通過單因素方差分析(One-way ANOVA)檢驗了海拔對PD,MPD以及MNTD的影響。所有的繪圖以及統計分析均在R.4.0.2軟件中實現。
2 結果與分析
2.1 放牧對土壤真菌群落系統發育α多樣性的影響
對照處理的PD,MPD以及MNTD在三個海拔間沒有顯著差異,放牧處理間也不存在顯著性差異(表1)。放牧處理并沒有顯著改變ASMWP,ASMSP以及AMSP的土壤真菌群落系統發育α多樣性(圖2)。
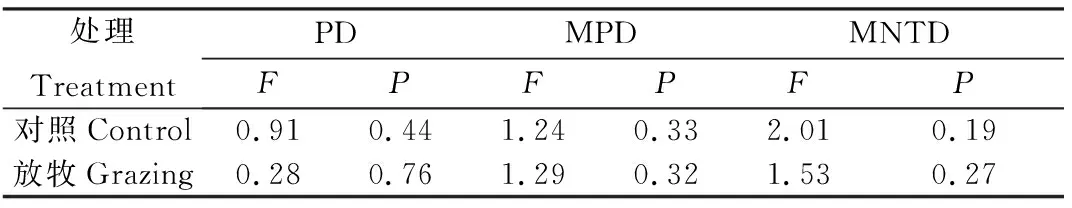
表1 三海拔間的土壤真菌群落系統發育α多樣性的方差分析Table 1 Variance analysis of phylogenetic α diversity of soil fungal community among three altitudes
2.2 對照和放牧處理間的土壤真菌群落βMNTD
對土壤真菌βMNTD進行NMDS和adonis分析發現,C_ASMWP與G_ASMWP,C_ASMSP與G_ASMSP,C_AMSP與G_AMSP之間在圖中明顯分開,P值均小于0.05(圖3a,表2)。多重比較的結果表明對照和放牧處理間的BMNTD在三個海拔間沒有顯著性差異(圖3b)。土壤速效磷、速效氮磷比和土壤銨態氮是影響對照和放牧間的βMNTD值的重要環境因子(圖4)。
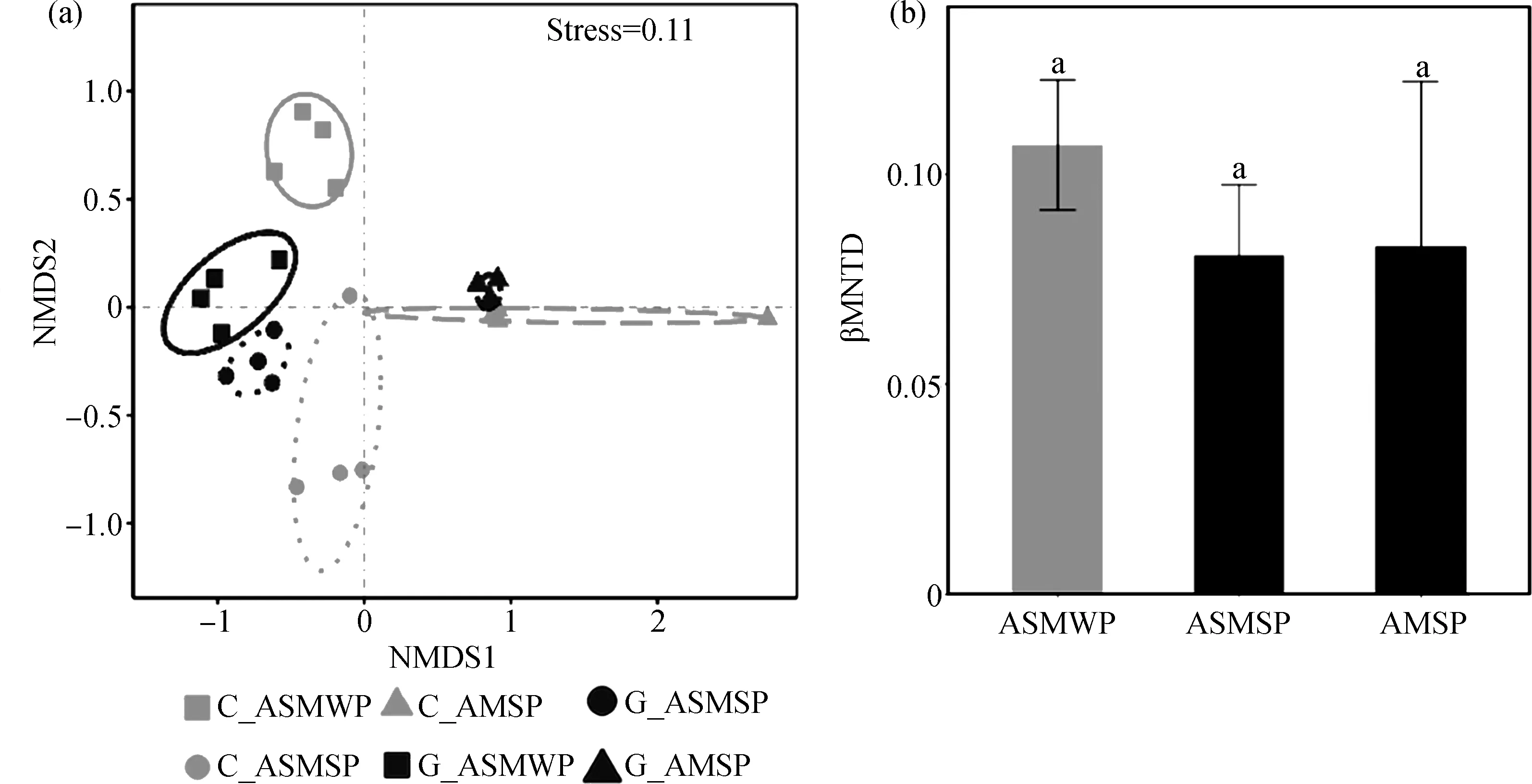
圖3 (a)土壤真菌系統發育β多樣性的非度量多維尺度分析,(b)三個海拔放牧和對照處理間的系統發育距離Fig.3 (a) Non-metric multidimensional scaling analysis for β-diversity of soil fungal community;(b) Phylogenetic distance between grazing and control treatments at three altitudes注:ASMWP,冷季放牧的高寒草原化草甸;ASMSP,暖季放牧的高寒草原化草甸;AMSP,暖季放牧的高寒草甸;C為對照處理,G為放牧處理Note:ASMWP,alpine steppe meadow for winter pasture;ASMSP,alpine steppe meadow for summer pasture;AMSP,alpine meadow for summer pasture;C,the control treatment and G,the grazing treatment
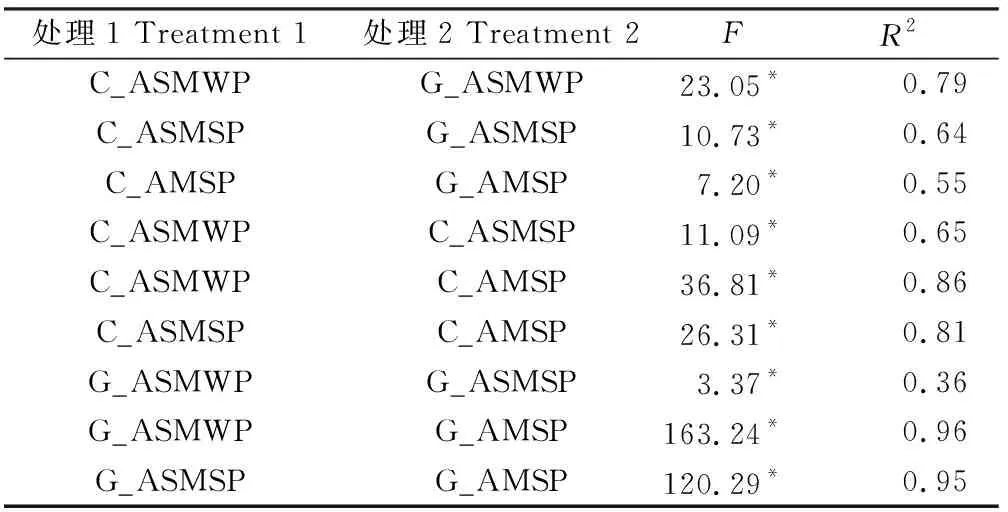
表2 基于βMNTD距離的Adonis檢驗Table 2 Adonis test based on βMNTD distance
2.3 放牧對土壤環境因子的海拔分布格局的影響
放牧后,ASMWP與ASMSP樣地間土壤含水量和硝態氮的差異減少,而速效磷的差異增加;ASMWP與AMSP樣地間土壤含水量、銨態氮、pH值以及速效氮磷比增加,而土壤硝態氮的差異減少;ASMSP與AMSP樣地間銨態氮及速效氮磷比增加,而土壤硝態氮的差異減少(圖5)。
2.4 放牧對土壤真菌群落系統發育β多樣性的海拔分布格局的影響
放牧導致了ASMWP與ASMSP樣地間的βMNTD的顯著降低,但是放牧和對照處理下ASMWP與AMSP以及ASMSP與AMSP樣地間的βMNTD未發生顯著變化(圖6)。對照處理下兩個海拔間(C3C5,C3C7以及C5C7)的土壤真菌β多樣性以及放牧處理間兩個海拔間(G3G5,G3G7以及G5G7)的土壤真菌β多樣性與環境因子的ABT分析結果表明,對照處理下土壤真菌β多樣性與速效氮磷比(20.3%)、銨態氮(19.5%)以及土壤含水量(17.0%)的距離矩陣相關性較大;放牧處理下土壤真菌β多樣性與土壤含水量(37.5%)、硝態氮(16.3%)以及速效氮磷比(13.6%)的距離矩陣相關性較大(圖4)。
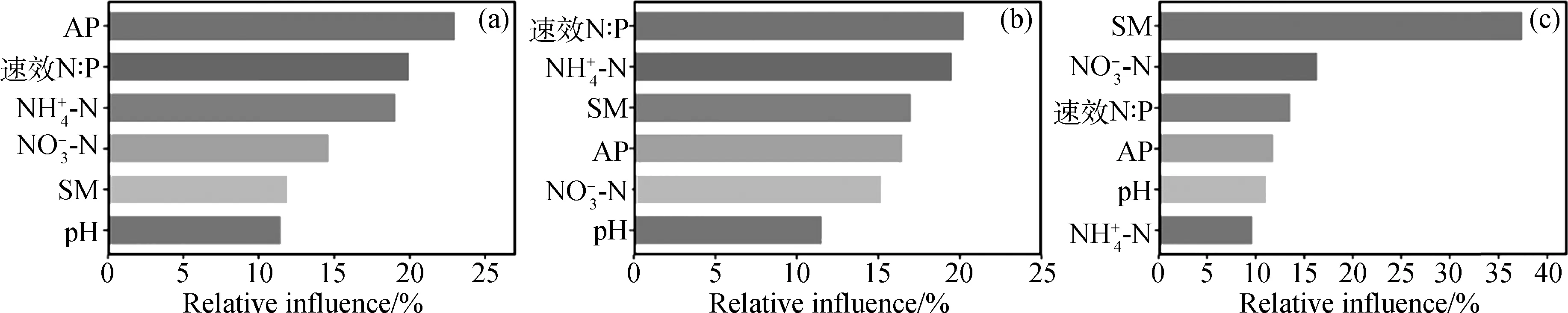
圖4 基于Aggregated boosted tree(ABT)分析的土壤真菌系統發育β多樣性與環境因子(兩個樣地間的基于歐式距離的距離矩陣)的關系Fig.4 Environmental drivers of phylogenetic β-diversity of soil fungal community composition according to the Aggregated boosted tree (ABT) analysis注:AP,速效磷;銨態氮;硝態氮;SM,土壤含水量;速效N∶P,(銨態氮+硝態氮)/速效磷。(a)為C3G3,C5G5以及C7G7之間的βMNTD與環境因子的關系,(b)為C3C5,C3C7以及C5C7之間的βMNTD與環境因子的關系,(c)為G3G5,G3G7以及G5G7之間的βMNTD與環境因子的關系Note:AP,available nitrogen;SM,soil moisture;Available N∶P,(ammonium nitrogen + nitrate nitrogen)/available P.(a) the relationship between β-MNTD and environmental factors among C3G3,C5G5 and C7G7,(b) the relationship between β-MNTD and environmental factors among C3C5,C3C7 and C5C7,and (c) the relationship between β-MNTD and environmental factors among G3G5,G3G7 and G5G7
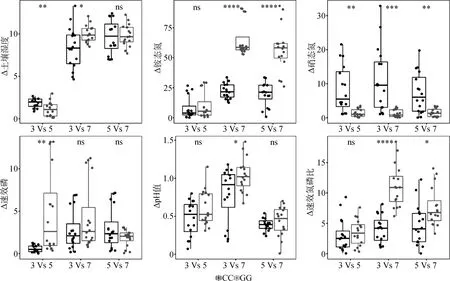
圖5 放牧和對照處理下樣地間環境因子的歐氏距離Fig.5 Euclidean distance of environmental factors between plots under grazing and control treatments注:CC為兩個對照樣地之間,GG為兩個放牧樣地之間。*為P <0.05,**為P <0.01,***為P <0.001,ns為P >0.05Note:CC between two control plots,GG between two grazing plots. * is P <0.05,** is P <0.01,*** is P <0.001,ns is P>0.05
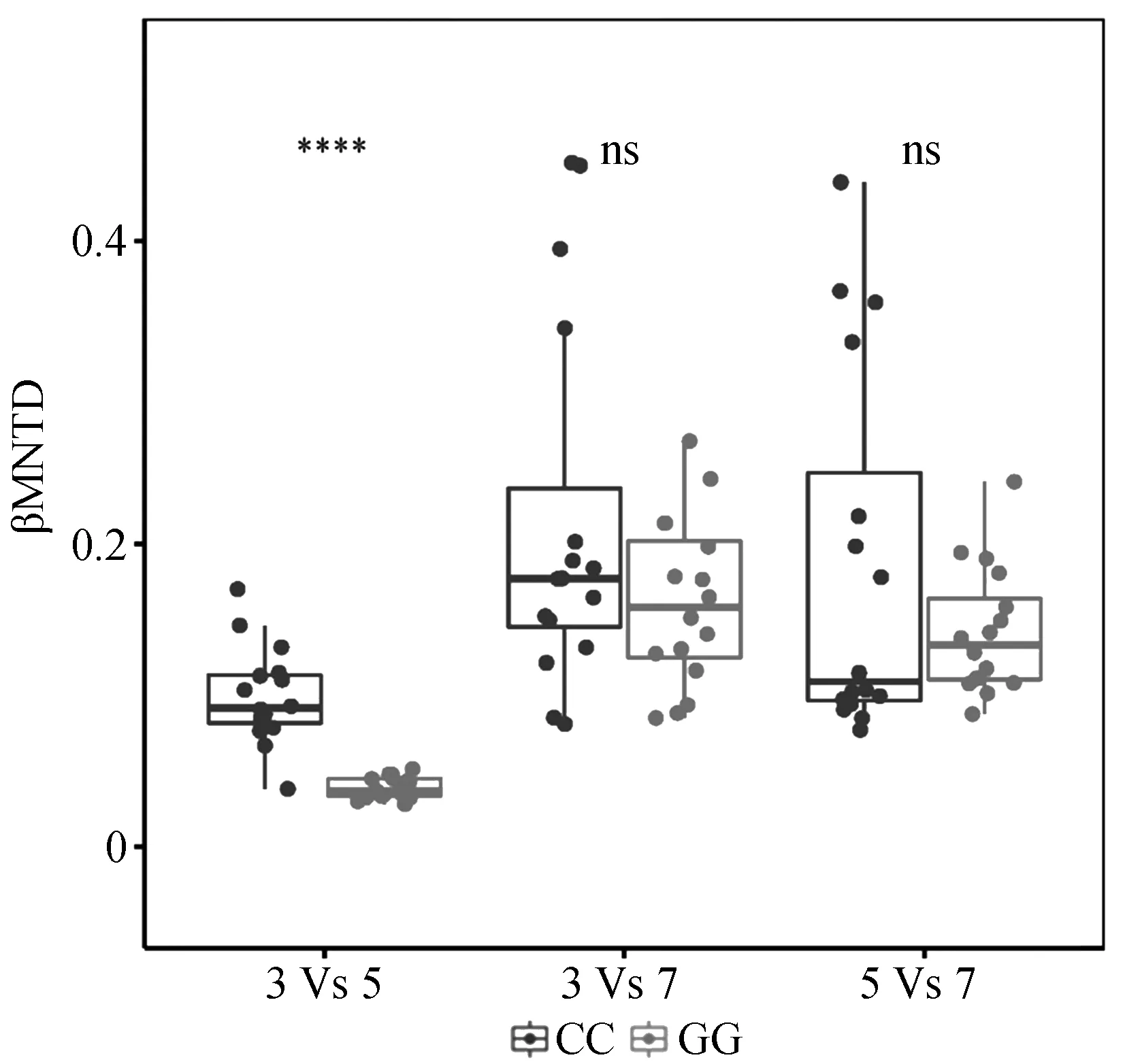
圖6 放牧以及對照處理下兩個海拔間的土壤真菌群落βMNTDFig.6 βMNTD of soil fungal community between two sites under grazing and control treatment注:CC為兩個對照樣地之間的βMNTD,GG為兩個放牧樣地之間的βMNTDNote:CC is βMNTD between two control plots,GG is βMNTD between two grazing plots
3 討論
3.1 冷季放牧和暖季放牧對土壤真菌群落系統發育多樣性的影響
冷季放牧和暖季放牧均未顯著改變土壤真菌群落系統發育α多樣性,與Zhang等[7]的研究一致,說明土壤真菌群落系統發育和物種α多樣性對冷季放牧和暖季放牧可能都不敏感。
雖然冷季放牧和暖季放牧都顯著改變了土壤真菌群落系統發育結構,但是它們的調控機制并不相同:冷季放牧所導致的土壤銨態氮、速效氮磷比和土壤含水量的變化可能是導致高寒草原化草甸土壤真菌群落系統發育結構變化的重要原因,而暖季放牧所導致的土壤硝態氮的變化可能是導致高寒草原化草甸土壤真菌群落系統發育結構變化的重要原因。我們之前的研究[7]表明,冷季放牧主要通過改變屬于土壤真菌子囊菌門和擔子菌門的某些關鍵種的相對豐度影響高寒草原化草甸土壤真菌群落物種組成,而暖季放牧卻主要通過改變擔子菌門的某些關鍵種的相對豐度影響高寒草原化草甸土壤真菌群落物種組成。因此,雖然冷季放牧和暖季放牧對高寒草原化草甸土壤真菌群落系統發育結構的影響強度沒有顯著差異(圖3),但是冷季放牧對高寒草原化草甸土壤真菌群落系統發育結構的影響可能主要與子囊菌門和擔子菌門的系統發育信息的變化有關,而暖季放牧對高寒草原化草甸土壤真菌系統發育結構的影響則主要與擔子菌門的系統發育信息的變化有關。這與Shi等[21]的研究結果并不一致,他們發現冷季刈割改變了叢枝菌根真菌群落系統發育α多樣性,但并未改變叢枝菌根真菌群落系統發育β多樣性。這可能是由于冷季刈割模擬冷季放牧只模擬了牛羊對牧草的啃食,而并未考慮牲畜的踩踏對土壤物理性質的改變和牲畜糞便對土壤化學性質的影響[22-26]。
3.2 不同類型高寒草地土壤真菌系統發育多樣性對放牧的響應
本研究發現暖季放牧對高寒草原化草甸和高寒草甸的土壤真菌系統發育α多樣性都無顯著影響,這與Zhang等[7]的研究一致,表明高寒草甸和高寒草原化草甸的土壤真菌群落系統發育和物種α多樣性對暖季放牧的響應可能都不敏感。
我們之前的研究表明暖季放牧引起的高寒草原化草甸的土壤真菌群落的物種組成的變化主要是由于擔子菌門的變化造成的,而暖季放牧引起的高寒草甸的土壤真菌群落的物種組成的變化則主要與子囊菌門的變化有關[7]。因此,雖然暖季放牧導致的高寒草原化草甸的土壤真菌群落系統發育結構的變化與暖季放牧導致的高寒草甸的土壤真菌群落系統發育結構的變化沒有顯著差異,但是這并不表明暖季放牧導致的兩種類型高寒草地的土壤真菌群落系統發育結構的變化是完全相同的:暖季放牧導致的高寒草原化草甸的土壤真菌群落的系統發育結構的變化可能主要與擔子菌門的系統發育結構有關,而暖季放牧導致的高寒草甸的土壤真菌群落的系統發育結構的變化則可能主要與子囊菌門的系統發育結構的變化有關。暖季放牧增加了高寒草甸的土壤銨態氮含量,而減少了高寒草原化草甸的土壤硝態氮含量[7]。因此,暖季放牧導致的高寒草原化草甸和高寒草甸的土壤真菌群落系統發育結構的變化與暖季放牧導致的高寒草原化草甸和高寒草甸的土壤速效氮的不同變化有關。
3.3 放牧改變了土壤真菌群落系統發育β多樣性的海拔分布格局
ASMWP和ASMSP樣地間的βMNTD由于不同季節放牧的影響而顯著降低,即放牧使得兩個海拔間的土壤真菌群落的系統發育結構變得更相似。這可能與對照和放牧處理下土壤真菌群落系統發育β多樣性的環境調控因子不同有關:對照條件下土壤速效氮磷比、銨態氮以及土壤含水量與土壤真菌群落系統發育β多樣性的相關性較大,而放牧條件下土壤含水量成為影響土壤真菌群落系統發育β多樣性的主導因子,且放牧使得兩個海拔間的土壤含水量變得更為相似(歐氏距離減少49.1%)。因此,放牧引起的土壤含水量的相似性的增加是造成兩個海拔間系統發育結構變得更相似的重要原因。
4 結論
本研究依托藏北高原的三個配對的自由放牧和圍欄試驗樣地發現:放牧均沒有顯著改變ASMWP,ASMSP和AMSP的土壤真菌群落系統發育α多樣性;放牧均顯著改變了ASMWP,ASMSP和AMSP的土壤真菌群落系統發育結構,但放牧對土壤真菌群落系統發育結構的影響并不完全一致;放牧改變了土壤真菌群落系統發育β多樣性的海拔分布格局。
