DNA甲基化修飾的定位分析方法研究進展
2022-02-18 09:38:06陳夢園游雪嬌袁必鋒馮鈺锜
分析測試學報 2022年1期
陳夢園,游雪嬌,袁必鋒,2*,馮鈺锜,2
(1.武漢大學 化學與分子科學學院,湖北 武漢 430072;2.武漢大學 公共衛生學院,湖北 武漢 430071)
DNA 是一種由磷酸脫氧核糖主鏈和含氮堿基腺嘌呤(Adenine,A)、鳥嘌呤(Guanine,G)、胞嘧啶(Cytosine,C)和胸腺嘧啶(Thymine,T)組成的天然聚合物,用來儲存、編碼、傳遞和表達遺傳信息[1]。除了正常堿基外,DNA 中還包含許多化學修飾[2-4]。這些修飾不會改變DNA 的序列,但會調節轉錄、細胞分化等生理過程,與疾病的發生密切相關[5-6]。其中,5-甲基胞嘧啶(5-Methylcytosine,5mC)作為DNA 中含量最高的甲基化修飾,存在于大多數植物、動物和真菌中,是最具代表性的表觀遺傳修飾之一[7-9]。
5mC是一種甲基化發生在胞嘧啶第五位碳上的修飾,該修飾參與多種生理過程,比如基因組印記、X 染色體失活和細胞分化等[10-13]。研究發現,異常的高甲基化可能會使正常細胞中的潛在抑癌基因沉默,而異常的低甲基化則可能會導致原癌基因激活,從而引起癌癥的發生[14-16]。Dnmt1(DNA methyltransferase 1)、Dnmt3a 和Dnmt3b 是哺乳動物細胞中常見的3 種5mC 的DNA 甲基轉移酶[17-18],它們以S-腺苷甲硫氨酸(S-Adenosylmethionine,SAM)為甲基供體,在DNA 胞嘧啶的五位碳上加上甲基基團構成。5mC 的形成過程如圖1所示[19]:DNA 甲基轉移酶通過具有活性的半胱氨酸殘基進攻胞嘧啶C6位形成中間體,該中間體的C5 位對SAM 親核進攻從而甲基化,隨后C6 位的半胱氨酸殘基離去得到5mC。
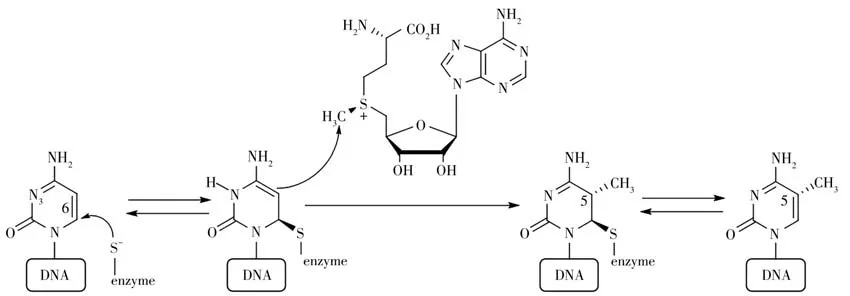
圖1 DNA甲基化過程示意圖Fig.1 Schematic illustration of DNA methylation
DNA 甲基化修飾在體內是動態平衡的,隨著5mC甲基化過程的發現,其去甲基化過程最近幾年逐漸清晰和完善。DNA 去甲基化的方式一般有兩種:被動去甲基化和主動去甲基化[20]。被動去甲基化是指在基因組DNA的半保留復制過程中,已存在的5mC含量因被動“稀釋”而下降,從而達到被動去甲基化的目的[20]。堿基切除修復(Base excision repair,BER)介導的DNA 主動去甲基化是目前研究得較透徹的生物途徑(圖2)[21]。在該DNA 去甲基化途徑中,5mC 在TET(Ten-eleven translocation)蛋白的作用下,逐步被氧化成5-羥甲基胞嘧啶(5-Hydroxymethylcytosine,5hmC)、5-醛基胞嘧啶(5-Formylcytosine,5fC)和5-羧基胞嘧啶(5-Carboxylcytosine,5caC)[22]。胸腺嘧啶DNA 糖苷酶(Thymine DNA glycosylase,TDG)能將5fC和5caC上的堿基切除,形成一個無堿基位點;隨后無堿基位點通過BER途徑被修復成正常的胞嘧啶,從而實現DNA 的主動去甲基化[21,23]。除此之外,還有另外兩種可能的途徑:(1)5mC 經TET 蛋白氧化生產的5hmC 經AID(Activation-induced cytidine deaminase)和APOBEC(Apolipoprotein B mRNA editing enzyme,catalytic polypeptide-like)蛋白脫氨成5-羥甲基尿嘧啶(5-Hydroxymethyluracil,5hmU),5hmU 隨后被TDG 或SMUG1(Single-stranded-selective monofunctional uracil DNA glycosylase 1)蛋白切除,形成無堿基位點,再由BER 途徑修復成正常胞嘧啶;(2)TET 蛋白氧化5mC 生產5fC 和5caC,5fC和5caC可以直接脫醛基或脫羧基生產未修飾的胞嘧啶(圖2)。目前已證實在哺乳動物基因組中確實存在直接脫醛基[24-25]和脫羧基[21,26-27]途徑,這些研究為后續脫醛基酶和脫羧酶的鑒定奠定了基礎。
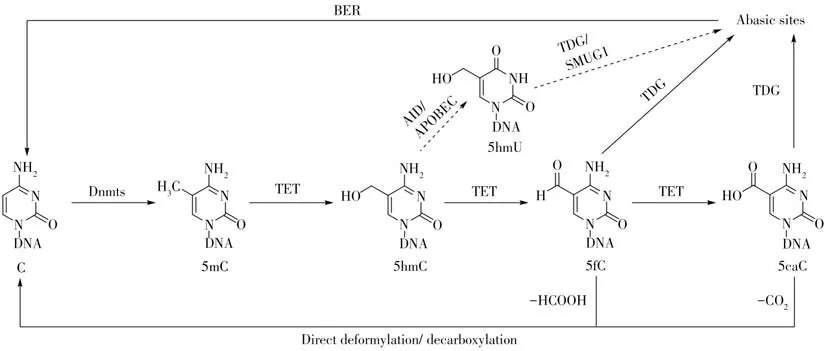
圖2 DNA主動去甲基化機制Fig.2 Mechanism for active DNA demethylation
5mC在生物體內發揮著重要生理功能,其甲基化的異常與各種疾病密切相關。研究DNA 甲基化和去甲基化過程中的相關蛋白和調節因子,能促進對發育和細胞穩態的理解,為臨床應用提供參考[28-29]。為了深入探索DNA 甲基化修飾的功能,需要了解其在基因組中的具體位置信息。近些年高通量測序技術的進步推動了DNA甲基化修飾領域的研究進展。本文總結了基于高通量測序技術的5mC定位分析方法的研究和應用情況(表1)。
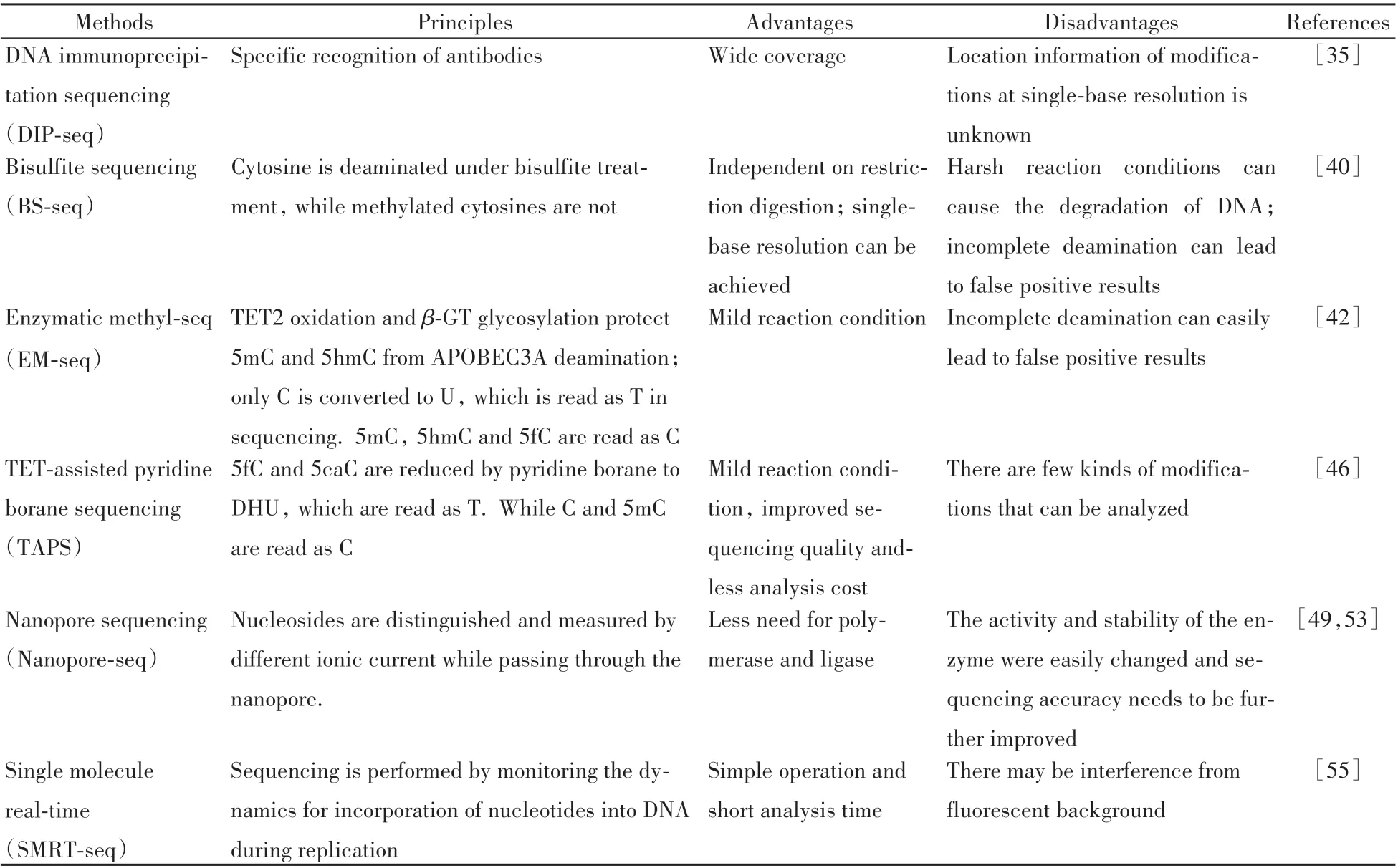
表1 基于高通量測序對5-甲基胞嘧啶(5mC)進行定位分析的方法Table 1 Summary of analytical methods for localizing 5-methylcytosine(5mC)based on high-throughput sequencing technology
1 5mC的定位分析方法
1.1 免疫沉淀富集介導的定位分析
由于核酸修飾的豐度通常較低,故將含有堿基修飾的片段進行富集有助于測序。Ruike等[30]證明使用特異性抗體富集含有堿基修飾的DNA 片段可以加強對核酸修飾的定位分析。基于此,研究者開發了多種基于免疫沉淀富集的方法,結合高通量測序,對基因組DNA甲基化進行定位分析,即DNA免疫沉淀測序(DNA immunoprecipitation sequencing,DIP-seq)。常用兩種方法對甲基化DNA 進行免疫沉淀:甲基化DNA 免疫共沉淀測序(Methylated DNA immunoprecipitation and sequencing,MeDIP-seq)[31]和甲基化DNA 特異性結合蛋白富集甲基化測序(Methyl-CpG binding domain protein-enriched genome sequencing,MBD-seq)[32-33]。
MeDIP-seq 是利用5mC 抗體富集含有5mC 的單鏈DNA 片段,將富集后的片段進行高通量測序的一種全基因組甲基化檢測手段[31,34]。類似地,基于抗體富集的測序方法也適用于DNA 中的其他修飾[35]。由于甲基化CpG結合(Methyl-CpG-binding domain,MBD)蛋白對甲基化序列有很強的親和性,能被用來對DNA 甲基化進行分析,因此研究者開發了一種將MBD 與高通量測序相結合的方法,即MBD-seq[33]。在MBD 蛋白家族中,MBD1、MBD2、MBD3 和MeCP2(Methyl-CpG-binding protein-2)4 種蛋白有相似的結構域,能識別雙鏈DNA 中CpG 雙核苷酸上的對稱甲基[35]。MBD2b(Methyl-CpG-binding domain protein 2b)作為MBD2 的一種亞型蛋白,與MBD3L1(Methyl-CpG-binding domain protein 3-like-1)蛋白的作用對調節甲基化DNA 復合物的形成至關重要,因此常用MBD2蛋白進行MBD-seq分析[32-33]。基于MBD蛋白的發現和應用,發展了一系列全基因組DNA 甲基化的檢測方法。除MBD-seq 外,還有MIRA-seq(Methylated-CpG island recovery assay sequencing)、MiGS(MBD-isolated genome sequencing)、MethylCapseq(Methylation DNA capture sequencing)等,均能用于5mC的定位分析[35-38]。
基于免疫沉淀的測序方法可以發現基因組中甲基化的區域,但無法實現甲基化修飾的單堿基分析[29]。抗體的高度特異性對測序結果的準確性至關重要。此外,商品化的抗體種類較少,且批次間抗體質量不一致,因此,基于免疫沉淀富集介導的5mC 定位分析對用來富集的抗體或者蛋白要求很高[35]。
1.2 亞硫酸氫鹽介導的定位分析
亞硫酸氫鹽測序法(Bisulfite sequencing,BS-seq)是指將DNA 甲基化修飾經亞硫酸氫鹽轉化后進行高通量測序的一種定位分析方法,已被廣泛應用于全基因組分析中[39]。在傳統的亞硫酸氫鹽測序方法中,C、5fC 和5caC 經亞硫酸氫鹽處理后脫氨變成尿嘧啶(Uracil,U),在測序過程中被讀成T;而5mC和5hmC 經處理后不脫氨,仍被讀成C(圖3)。將亞硫酸氫鹽處理和未處理的測序樣本進行比較可以發現甲基化的位點,但是該方法無法區分5mC 和5hmC[40-41]。因此,為了對5mC 進行定位分析,需要對傳統的亞硫酸氫鹽測序方法進行改進。
氧化亞硫酸氫鹽測序法(Oxidative bisulfite sequencing,oxBS-seq)[40]和TET 輔助的亞硫酸氫鹽測序法(TET-assisted bisulfite sequencing,TAB-seq)[41]能實現5mC 的單堿基定位分析。在oxBS-seq 中,5hmC被KRuO4氧化成5fC,經亞硫酸氫鹽處理后脫氨讀成T,而5mC 依舊被讀成C[40],從而可在單堿基水平分辨5mC和5hmC(圖3)。
此外,還可以用傳統亞硫酸氫鹽測序的方法間接確定5mC 的位點(圖3)。在TAB-seq 中,首先在β-葡萄糖基轉移酶(β-Glucosyltransferase,β-GT)的作用下將葡萄糖基轉移至5hmC,形成的β-葡萄糖基-5-羥甲基胞嘧啶(β-Glucosyl-5-hydroxymethylcytosine,5gmC)不被TET 蛋白氧化,在測序時被讀成C;而5mC 經TET 蛋白氧化成5caC,在測序時被讀成T,從而可實現5hmC 的單堿基定位分析。將傳統亞硫酸氫鹽測序法確定的5mC 和5hmC 總位點減去5hmC 的位點,即可實現對5mC 位點的間接分析[41]。
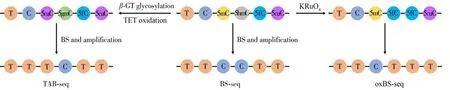
圖3 亞硫酸氫鹽測序原理圖Fig.3 Schematic diagram of bisulfite sequencing
亞硫酸氫鹽介導的定位分析方法存在兩個不足:一是該反應需要的條件苛刻,使得DNA 大量斷裂,不利于后續的高通量測序[42];二是未修飾胞嘧啶在人類總基因組胞嘧啶中所占比例超過95%,將這部分胞嘧啶轉化成胸腺嘧啶使得序列復雜度、測序質量以及覆蓋率降低,給后續分析造成較大挑戰[28]。此外,未修飾的胞嘧啶如果脫氨不完全,可能產生假陽性結果。因此,需要尋找更合適的方法對5mC進行定位分析。
1.3 酶介導的定位分析
為了克服亞硫酸氫鹽對DNA 的損傷,研究人員開發了一種比較溫和的方法,即酶促甲基化測序法(Enzymatic methyl-seq,EM-seq)[42-43]。該方法依賴于酶分析定位DNA 胞嘧啶的甲基化狀態,避免了過于劇烈的化學反應條件[42-43]。APOBEC3A(Apolipoprotein B m RNA editing enzyme catalytic polypeptidelike 3A)是細胞的抗病毒功能因子之一,可使單鏈DNA 的胞嘧啶殘基脫氨基轉化為尿嘧啶[42]。在單鏈DNA 中,APOBEC3A 能將C 和5mC 脫氨,在隨后的聚合酶鏈式反應(PCR)中二者均會被讀成T[42]。為了對5mC 進行單堿基定位分析,在APOBEC3A 脫氨前對5mC 和5hmC 進行選擇性保護,以在堿基水平上間接分析5mC[42]。具體來說,先使用β-GT 將5hmC 糖基化成5gmC,再用TET 蛋白將5mC 氧化成5caC,5gmC 和5caC 不被APOBEC3A 脫氨,在PCR 中依舊被讀成C,而C經歷脫氨被讀成T[44-45](圖4)。如果只將5hmC 進行糖基化保護,經APOBEC3A 處理后,只有5gmC 被讀成C,其余均被讀成T。將兩種方法相結合即可間接確定5mC的位點[42,44]。
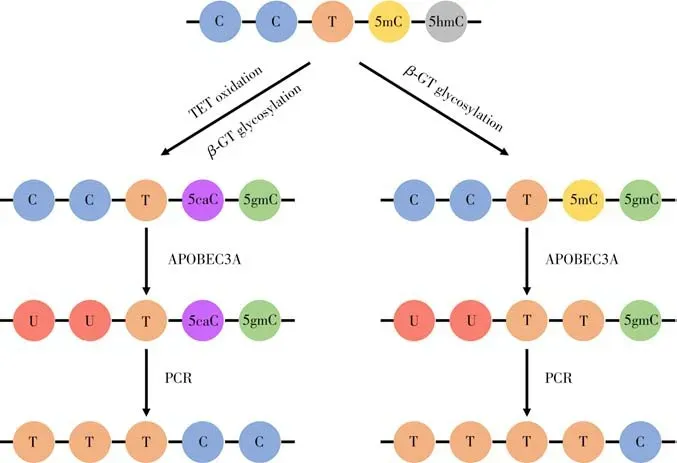
圖4 酶促甲基化測序原理圖Fig.4 Schematic diagram of enzymatic methyl sequencing
相對于亞硫酸氫鹽測序法而言,EM-seq具有條件溫和、可維持DNA完整性的優點[43],進一步擴大了應用范圍。然而,該方法同樣存在脫氨不完全造成假陽性、測序質量較差的問題[46]。
1.4 吡啶硼烷介導的定位分析
Liu 等[46]開發了TET 輔助吡啶硼烷測序法(TETEM-seqassisted pyridine borane sequencing,TAPS):利用TET 酶將5mC 和5hmC 氧化成5caC,再通過吡啶硼烷將5caC 還原形成二氫尿嘧啶(Dihydrouracil,DHU),DHU 在之后的測序中被讀成T,從而實現5mC 和5hmC 從C 到T 的轉變[35]。該方法可以用來定位分析基因組中的5mC 和5hmC,但無法將二者區分[46]。為此需要改進TAPS 方法以實現5mC 的定位分析。基于此,Liu 等[28,46]開發了β-葡萄糖基轉移酶阻斷的TAPS(TAPS withβ-glucosyltransferase blocking,TAPSβ)和化學輔助吡啶硼烷測序(Chemical-assisted pyridine borane sequencing,CAPS)。
在TAPSβ中,將葡萄糖基引入5hmC 使其糖基化成5gmC 后,進行TET 氧化和吡啶硼烷還原反應,該過程中5mC經歷了從5mC到5caC再到DHU,最后被讀成T的過程,而5gmC不參與氧化和還原過程,依舊被讀成C[28,35]。除了還原5caC 外,吡啶硼烷也能將5fC還原成DHU,基于此開發了CAPS方法。使用KRuO4將5hmC 氧化成5fC,再經吡啶硼烷還原成DHU,測序時被讀成T,而5mC 不參與任何一步反應,依舊被讀成C[28](圖5)。
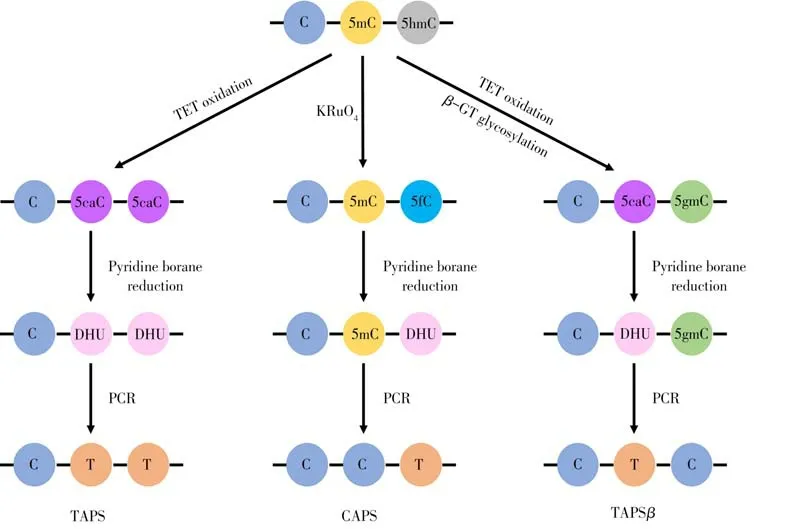
圖5 吡啶硼烷測序原理圖Fig.5 Schematic diagram of TET-assisted pyridine borane sequencing
與亞硫酸氫鹽測序和酶促甲基化測序方法相比,TAPS 方法顯著降低了DNA 的損傷,提高了測序質量,避免了未修飾胞嘧啶的轉化。此外,也降低了分析的成本[28,46-47]。TAPSβ是一種不依賴亞硫酸氫鹽、堿基分辨率高、能定量5mC 特異性位點的分析方法[46],但是,該方法中吡啶硼烷的還原效率為90%多,可能會造成測序結果存在一定的誤差。
1.5 納米孔測序定位分析
納米孔測序已經成為一種有前景的DNA甲基化修飾分析方法[48-49]。不同的核苷通過納米孔會產生不同的電流,根據電流的變化可以分辨正常核苷和修飾核苷[50]。目前,納米孔測序已成功用于區分DNA中的5mC和正常胞嘧啶[50-51],從而實現5mC的單堿基定位分析。此外,該方法也可用于DNA分子中5hmC的定位分析[48,52]。
與其他高通量測序方法相比,納米孔測序具有無需擴增、可減少聚合酶和連接酶的使用等優點[49],但該方法目前仍然處于發展階段,需要進一步優化維持酶活性和穩定性的條件[53]。
1.6 單分子實時測序定位分析
除納米孔測序外,單分子實時(Single molecule real-time,SMRT)測序作為另一種有前景的DNA測序方法,能直接對DNA分子上的5mC進行定位分析[54-55]。在單分子實時測序中,DNA聚合酶催化核苷酸進入DNA 中,通過監測每種核苷酸獨特的動力學特征,可以對DNA 中的不同堿基進行區分[29,48,55-56]。由于未修飾堿基和修飾堿基在核苷酸摻入時間上存在差異,該方法可被用于對5mC[55]、5hmC[57]、6-甲基腺嘌呤[58]和細菌硫代磷酸[59]的直接檢測。使用TET1將5mC氧化成5caC能增強動力學特征,提高對5mC的檢測能力[60]。
單分子實時測序方法的出現為微量樣品的分析提供了可能,具有測序通量高、成本低、時間短、操作簡單的優點,而如何降低熒光信號對背景噪音的影響是一個需要解決的問題。
2 結論與展望
目前已在生物體DNA和RNA中發現了多種化學修飾,其中,5mC作為DNA分子中的“第五堿基”,參與多種生理過程,如胚胎發育、轉錄異常、基因沉默。現有分析方法的開發推動了5mC的研究進展,破譯5mC 的位置信息有助于深入闡明5mC 的生物學功能,增強對生物生長發育、疾病發生等問題的理解。而對5mC生物學功能的研究需要靈敏度高、準確度高以及穩定性好的方法,這些方法的開發也將促進對應技術和軟件的改進與提升。
已報道的化學試劑或酶結合高通量測序對5mC進行定位分析的方法在選擇性標記DNA修飾方面具有良好的應用前景,但存在反應條件較為劇烈、易造成假陽性等局限性,這對精確定位5mC 來說是一個挑戰。未來可探索新的化學方法、開發新的化學試劑或采用更溫和的反應條件來提高5mC 的單堿基分辨率以及實現對單個5mC 位點的量化。使用第三代測序技術對5mC 進行定位分析是一個很有前景的方向,納米孔測序和單分子實時測序為直接對5mC 進行定位提供了可能。未來可通過研究如何維持酶的活性和穩定性以及降低熒光信號的干擾對納米孔測序和單分子實時測序進行改進。此外,現有技術精度、靈敏度的改進也可能促進對其它低豐度化學修飾的測序研究,甚至推進新修飾的發現。修飾位點的確定以及修飾種類的擴充有助于在基因層面加深對疾病的研究,從而對DNA甲基化修飾譜圖進行更加具體的闡述。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
兒童故事畫報(2019年5期)2019-05-26 14:26:14
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
Coco薇(2016年2期)2016-03-22 02:42:52
Coco薇(2015年1期)2015-08-13 02:47:34
小雪花·成長指南(2015年7期)2015-08-11 15:03:12
小雪花·成長指南(2015年4期)2015-05-19 14:47:56
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
