不同生境來源硝化細菌群對氨氮的去除性能
2022-02-20 17:09:24杜杭濤徐睿徐慧施文卿鄧皓元何俊龍朱琳
環境工程技術學報 2022年1期
杜杭濤,徐睿,徐慧,施文卿,2,鄧皓元,何俊龍,朱琳*
1.南京信息工程大學環境科學與工程學院,江蘇省大氣環境監測與污染控制高技術研究重點實驗室
2.中國科學院水生生物研究所
3.中國環境科學研究院
4.國家長江生態環境保護修復聯合研究中心
5.中國科學院生態環境研究中心,環境水質學國家重點實驗室
近年來,隨著城市人口的急劇增長和工農業生產的快速發展,造成污染源和外排污染物數量劇增,不少受納水體生態環境嚴重失衡[1]。氮作為水體中重要營養元素之一,當其含量超出一定水平后,則會引起水體富營養化,導致藍藻水華和赤潮現象頻發,對水生生物生存與人類健康構成巨大威脅。因此,如何高效脫氮成為了環境學科領域的研究重點[2-3]。在當前眾多脫氮技術中,微生物脫氮因效率高、費用低、安全性好等特點被廣泛運用于污水脫氮處理[4-5]。
微生物脫氮是利用硝化作用將氨氮轉化為亞硝態氮和硝態氮,最后由反硝化作用將硝態氮還原成氮氣從水體逸出的過程,從而達到從污水中脫去超標氮素的目的[6]。氮在污水中主要以氨氮、有機氮以及少量的亞硝態氮和硝態氮等形態存在。由于有機氮可被枯草芽孢桿菌、大腸桿菌、熒光假單胞菌、腐敗梭菌、變形桿菌等多種異養型細菌轉化為氨氮(氨化作用),且轉化效率很高,硝化過程則成為了微生物脫氮的關鍵,制約著后續反硝化過程的進行。因此,如何篩選高效硝化細菌是污水脫氮的關鍵。
硝化細菌是生物脫氨氮過程的主要功能菌群,可分為2 類:一類是將氨氮轉化為亞硝態氮的氨氧化細菌(ammonia-oxidizing bacteria,AOB);另一類為將亞硝態氮轉化為硝態氮的硝化細菌(nitriteoxidizing bacteria,NOB)[7-8]。目前,國內外眾多學者已對硝化細菌開展了大量的相關研究,但這些研究主要集中在硝化細菌脫氨氮效率及其影響因素等方面[9-14],而有關從何種生境篩選硝化細菌尚不清楚。筆者比較了沉積物、土壤自然環境和市售人為環境3 種生境硝化細菌,并利用分子生物學方法對各種群結構進行深入分析,以期為微生物脫氮工藝中硝化細菌菌種篩選提供參考。
1 材料與方法
1.1 硝化細菌富集
沉積物和土壤生境硝化細菌分別采集于云南省昆明市船房河沉積物和某菜地土壤,市售生境硝化細菌為天津日野南洋水族廠生產的硝化細菌菌液。富集培養液為人工配制,具體成分見文獻[15]。富集裝置為4 L 塑料桶,加入適量培養液后,用電磁式空氣壓縮機曝氣使水體DO 濃度大于2 mg/L。富集過程中,每2 d 換水1 次,每次換水前,先沉淀2 h,用虹吸法吸去上層水體,再加入等量的培養液。富集過程中溫度維持在(22±1)℃,pH 維持在8.0±0.2。
1.2 不同生境硝化細菌硝化速率
硝化速率測定在1 000 mL 錐形瓶中進行。為了利于硝化細菌附著,在每個錐形瓶中放置200 g 沸石。考慮到沸石對氨氮具有吸附性,本試驗硝化速率是通過測定反應器中硝酸氮濃度升高速率來實現的。硝酸氮濃度采用紫外分光光度法測定[16]。試驗前,為使得各反應系統穩定,加入沸石和等體積硝化細菌富集菌液后,在溫度為(22±1)℃,pH 為8.0±0.2條件下,用25 mg/L 氨氮濃度的人工配制污水曝氣培養,監測各反應器中的硝化速率,直至反應器的硝化速率穩定(約2~4 周)。每個生境類型硝化細菌設置3 個平行反應器。
1.2.1人工配制污水硝化速率
分別向各反應器中加入1 000 mL 25 mg/L 氨氮濃度的人工配制污水,在溫度為(22±1)℃、pH 為8.0±0.2、連續曝氣條件下培養,測定各反應器的硝化速率。
1.2.2生活污水硝化速率
實際生活污水取自昆明市蘭花溝河。將污水加入反應器前,過濾掉懸浮物雜質。取1 000 mL 預處理后的污水,分別加入各反應器中,用NaOH 溶液調pH 至8.0±0.2,在溫度(22±1)℃下曝氣運行,測定各反應器的硝化速率。
1.3 硝化細菌對不同環境因子的適應性
為了分析硝化細菌適應性,設置了各環境因子極端值。雖然在實際污水脫氮處理時這些極端條件不常見,但對于確定硝化細菌的各環境因子適應范圍,分析硝化細菌種群與功能差異性具有重要意義。
為研究硝化細菌對氨氮初始濃度的適應性,配制了氨氮初始濃度分別為5、50、100、150、200、250、300 mg/L 一系列人工污水。在溫度為20 ℃,pH為8.0,連續曝氣條件下,測定各反應器硝化速率。
為研究硝化細菌對溫度的適應性,將各反應器置于大容器全溫振蕩器(不使用振蕩功能)中,加入氨氮初始濃度為25 mg/L 人工配制污水,用導氣管連接氣泵對各反應器連續曝氣,調節pH 為8.0,測定5、10、20、30、40 ℃下各反應器的硝化速率。
為研究硝化細菌對pH 的適應性,在溫度為20 ℃,連續曝氣的條件下,用NaOH 調節反應器pH,測定各反應器在pH 為5.0、6.0、7.0、7.5、8.0、8.5、9.0、10.0、11.0 的硝化速率。
為研究硝化細菌對鹽度的適應性,在25 mg/L氨氮濃度的人工配制污水中加入NaCl,配制成鹽度梯度為0、5、10、15、20、25 g/L 的污水,在溫度為20 ℃,pH 為8.0 條件下,對各反應器連續曝氣,測定各反應器在各鹽度梯度下的硝化速率。
1.4 硝化細菌分子生物學分析
硝化細菌豐度采用MPN-PCR 計數法。從各反應器中取適量菌液,用Power Soil DNA Isolation Kit(MO BIO Laboratories,Inc.,Carlsbad,CA,美國)提取DNA,取定量DNA 進行梯度稀釋(1:10),然后對各梯度DNA 進行PCR 擴增(3 個重復),記錄各梯度下擴增條帶數目,由三管最大或然數表計算AOB、硝化桿菌(Nitrobacter)和硝化螺旋菌(Nitrospira)生物量。
利用引物amoA-1F 和amoA-2R 對amoA目的基因進行擴增,利用Gel Extraction Kit(Omega,美國)回收,連接到PEGMT-easy T 載體上后,轉化到Trans 5α 感受態細胞里,在Amp+LB 瓊脂平板培養基上培養,形成單菌落,挑選適量圓形白色菌落在500 μL Amp+LB 培養基里培養,委托中國科學院水生生物研究所分析測試中心測序,對測序結果進行系統發育分析。
由于Nitrobacter和Nitrospira是各生境主要的NOB 菌群[17-18],因此只對這2 個屬進行16S rRNA基因序列分析。Nitrobacter和Nitrospira的16S rRNA基因序列測定過程,除引物與AOB 的amoA基因測序不同外,其余步驟均相同。Nitrobacter和Nitrospira所使用的引物分別為FGPS 1 269/FGPS 872 和 NSR 113f/NSR 1264r。將測得的16S rRNA 基因序列分別進行基因型分析。
2 結果與討論
2.1 不同生境來源硝化細菌硝化速率差異
不同生境來源硝化細菌處理人工配制污水和實際生活污水時的硝化速率如圖1 所示。從圖1 可以看出,沉積物生境硝化細菌在處理人工配制污水和實際生活污水過程中,均表現出較高的效率,其硝化速率明顯高于土壤和市售生境。在處理人工配制污水時,沉積物生境硝化細菌的硝化速率達1.02 mg/(L·h),而土壤和市售生境硝化細菌的硝化速率僅為0.36 和0.56 mg/(L·h);在處理實際生活污水時,沉積物生境硝化細菌硝化速率為0.55 mg/(L·h),同樣顯著高于土壤和市售生境的0.18 和0.14 mg/(L·h)。不僅如此,與處理人工配制污水相比,3 種生境硝化細菌處理實際生活污水的硝化速率均有不同程度的下降。市售生境硝化細菌的硝化速率從0.56 mg/(L·h)降至0.16 mg/(L·h),降幅最大,達到71.4%;土壤生境硝化細菌次之,硝化速率從0.36 mg/(L·h)降至0.18 mg/(L·h),降幅為50%;沉積物生境硝化細菌降幅最小,硝化速率從1.02 mg/(L·h)降至0.55 mg/(L·h),降幅為46.1%。這表明,硝化細菌的活性易受外界環境條件的影響,污水水質的變化則會導致硝化速率的改變;并且,不同生境中的硝化細菌對環境條件的適應性不同,沉積物生境硝化細菌對環境的適應性最高,硝化速率隨環境條件的改變波動較小,土壤生境次之,市售生境適應性最低,硝化速率變化較大。這可能與它們的生存環境有關,沉積物生境受上覆水水質頻繁波動的影響變化較大,土壤生境較為穩定,而市售生境在人為調控作用下最為穩定。在這些不同生境條件的長期影響下,沉積物生境硝化細菌群落結構組成及其對環境的適應性要高于土壤和市售生境。
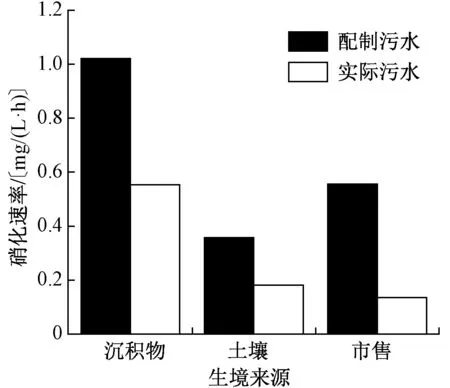
圖1 不同生境來源硝化細菌硝化速率差異性Fig.1 Differences in nitrification rates of nitrifying bacteria from different habitats
2.2 不同生境來源硝化細菌對環境因子適應性差異
2.2.1氨氮初始濃度
不同生境來源硝化細菌硝化速率對氨氮初始濃度的響應見圖2。從圖2 可以看出,沉積物和土壤生境硝化細菌的硝化速率隨氨氮初始濃度的升高變化趨勢相似,均呈先上升后下降的趨勢。主要原因是:氨氮作為硝化反應的底物,當其濃度升高時,促進硝化反應的進行,硝化速率隨之增大;當氨氮濃度超過一定水平后,游離氨濃度上升,對硝化細菌有一定的毒害作用,硝化細菌活性受到抑制,硝化速率隨之降低[19-21]。相比之下,市售生境硝化細菌的硝化速率隨氨氮初始濃度升高逐漸下降,表明稍高的氨氮初始濃度就會對市售硝化細菌活性產生抑制作用,該類硝化細菌對氨氮耐受力較差。沉積物生境硝化細菌耐受氨氮初始濃度的閾值為200 mg/L,低于此閾值時,硝化速率隨氨氮初始濃度增加快速上升,氨氮初始濃度每升高50 mg/L,硝化速率增加約0.15 mg/(L·h);當超過此閾值,硝化速率快速下降。土壤生境硝化細菌耐受氨氮初始濃度的閾值為150 mg/L,并且低于此閾值時,硝化速率隨氨氮初始濃度增加的變化趨勢不如沉積物生境明顯。這表明,沉積物生境硝化細菌耐受氨氮的能力高于土壤生境。

圖2 不同生境來源硝化細菌硝化速率對氨氮初始濃度的響應Fig.2 Nitrification rate response of denitrifying bacteria from different habitats to initial ammonium concentrations
2.2.2環境溫度
溫度對硝化細菌活性及其生長繁殖均有較大影響。大量研究表明,硝化細菌的適宜生長溫度為15~35 ℃,當溫度低于10 ℃時,硝化作用會受到抑制;高于20 ℃時,硝化細菌的活性較高,但超過38 ℃后硝化作用將會消失[22-23]。3 種生境來源硝化細菌的活性隨溫度升高的變化趨勢大體相同(圖3)。溫度低于10 ℃時,硝化速率較低;隨著溫度的上升,硝化速率逐漸增加;當溫度升高至30 ℃左右時,硝化速率最高;當溫度超過30 ℃后,硝化速率急劇下降。原因是硝化細菌的生物膜結構、蛋白質和核酸等活性容易受溫度的影響,主要體現在2 個方面:1)隨著環境溫度升高,硝化細菌體內酶活性增強,促使體內生化反應加快,硝化活性提高;2)當環境溫度超過一定范圍時,細胞中蛋白質、核酸等物質將受到不可逆轉的破壞,硝化細菌活性急劇下降,甚至出現菌體死亡現象[24]。然而,3 種生境硝化細菌的硝化速率隨溫度的變化幅度存在明顯差異,沉積物生境硝化細菌的變化幅度明顯高于土壤和市售生境。如當溫度從10 ℃升至20 ℃時,沉積物、土壤和市售生境硝化細菌的硝化速率增加幅度分別為0.70、0.24、0.34 mg/(L·h);當溫度從20 ℃升至30 ℃時,硝化速率增加幅度分別為1.33、0.47、0.02 mg/(L·h)。這主要是由硝化細菌生物量與種群組成的差異造成的。

圖3 不同生境來源硝化細菌硝化速率對環境溫度的響應Fig.3 Nitrification rate response of denitrifying bacteria from different habitats to temperature
2.2.3環境pH
適宜硝化作用的pH 為7.0~8.0,而適宜AOB和NOB 生長的pH 分別為6.0~7.5 和7.0~8.5[25-26]。當pH>8.0 時,AOB 的生長受到抑制,硝化活性降低,硝化作用進行緩慢;當pH<7.0 時,NOB 生長緩慢,硝化作用受到影響;pH 過高(> 9.0)或過低(<6.0),硝化作用則會變得十分微弱[27]。不同生境硝化細菌活性隨pH 變化趨勢相似,即pH 為7.0~8.0 時,3 種生境硝化細菌均表現出較高活性;當pH<7.0 和pH>8.0 時,硝化細菌活性受到抑制而降低(圖4)。pH 對硝化作用的影響主要通過以下3 種途徑實現:1)硝化細菌的活化與鈍化[28]。H+和OH-離子與硝化反應所涉及酶的可逆性結合,導致酶的失活與活化,從而引起硝化細菌活性的改變。2)對無機碳源的影響[29]。可溶性碳酸鹽作為硝化細菌的無機碳源,其含量高低容易受到pH 的影響。當pH 較低時,可溶性碳酸鹽轉化為CO2而逸出;當pH 較高時,可溶性碳酸鹽則轉化為難溶性碳酸鹽,很難被硝化細菌吸收利用。3)游離氨、亞硝酸[30]以及重金屬[31-32]的抑制作用。能為硝化細菌提供能量的氨氮和亞硝態氮,當以離子態NH3和HNO2存在時,對硝化細菌活性產生抑制作用。離子態NH3和HNO2濃度受pH 的影響較大。當pH 較低時,HNO2濃度較高;當pH 較高時,NH3濃度較高。因此,pH 過高或過低均會對硝化細菌產生抑制作用。
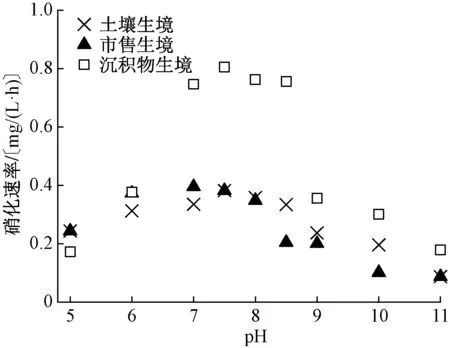
圖4 不同生境來源硝化細菌硝化速率對環境pH 的響應Fig.4 Nitrification rate response of denitrifying bacteria from different habitats to pH
雖然不同生境硝化細菌活性隨pH 變化趨勢相似,但在適宜的pH 范圍內,各生境硝化細菌的硝化速率存在明顯差異(圖4)。pH 為7.0~8.0 時,沉積物生境硝化細菌硝化速率明顯高于土壤和市售生境,且與2.1 節的研究結果一致。另外,沉積物生境硝化細菌的硝化速率在pH 大于8.5 時開始快速下降;而土壤和市售生境硝化細菌在pH 為8.0 時,硝化活性就已經開始表現出受抑制現象。這說明了沉積物生境硝化細菌耐受pH 的能力要稍高于土壤和市售生境。
2.2.4環境鹽度
當生活在淡水中的微生物受到含鹽廢水的沖擊時,一般能通過調節自身的滲透壓來保護細胞內的原生質。但鹽度過高時,微生物體內的水分子則會大量滲透到體外,導致微生物自身細胞因發生質壁分離而死亡。因此,污水鹽度的高低對微生物脫氮中硝化細菌的活性具有重要影響[33]。盡管各生境來源硝化細菌的活性均受鹽度的抑制,但沉積物生境硝化細菌表現出較高的耐鹽性(圖5)。當鹽度大于10 g/L 時,沉積物生境硝化細菌活性受到明顯抑制,硝化速率大幅度下降,鹽度為15 g/L 時硝化速率較鹽度為10 g/L 下降約54%。而土壤和市售生境硝化細菌的活性在鹽度大于5 g/L 時就已經開始受到明顯抑制,當鹽度升至10 g/L 時,硝化速率分別下降42.8%和65.2%。這可能是因為沉積物生境硝化細菌種群組成中存在某些耐鹽性菌類。

圖5 不同生境來源硝化細菌硝化速率對環境鹽度的響應Fig.5 Nitrification rate response of denitrifying bacteria from different habitats to salinity
2.3 各生境硝化細菌的MPN-PCR 計數
基于硝化細菌MPN-PCR 產物的電泳圖譜(圖6),經由三管最大或然數表[34]計算得出,沉積物、土壤和市售生境AOB 的生物量分別為1.1×107、3.5×108、6.3×107個,Nitrobacter生物量分別為5×104、1.9×105、5×104個,Nitrospira生物量分別為5×104、2.3×104、7.5×103個。進一步分析得知,土壤生境硝化細菌生物量(AOB 和NOB 的總生物量)最高,分別是沉積物和市售生境的31.8 和5.6 倍。但在種群結構組成上,沉積物生境硝化細菌種群較為平衡,AOB 生物量約為NOB 的110 倍,而土壤和市售生境AOB 分別約為NOB 的1 600 和1 100 倍。另外,3 種生境NOB 的組成也存在差異(圖7),沉積物生境NOB 的Nitrobacter和Nitrospira較為均衡,各占約50%;而土壤和市售生境NOB 主要為Nitrospira,分別占各自NOB 的89.2%和87.0%。

圖6 不同生境來源硝化細菌MPN-PCR 產物的瓊脂糖凝膠電泳圖譜Fig.6 Agarose gel electrophoresis for MPN-PCR products of denitrifying bacteria from different habitats
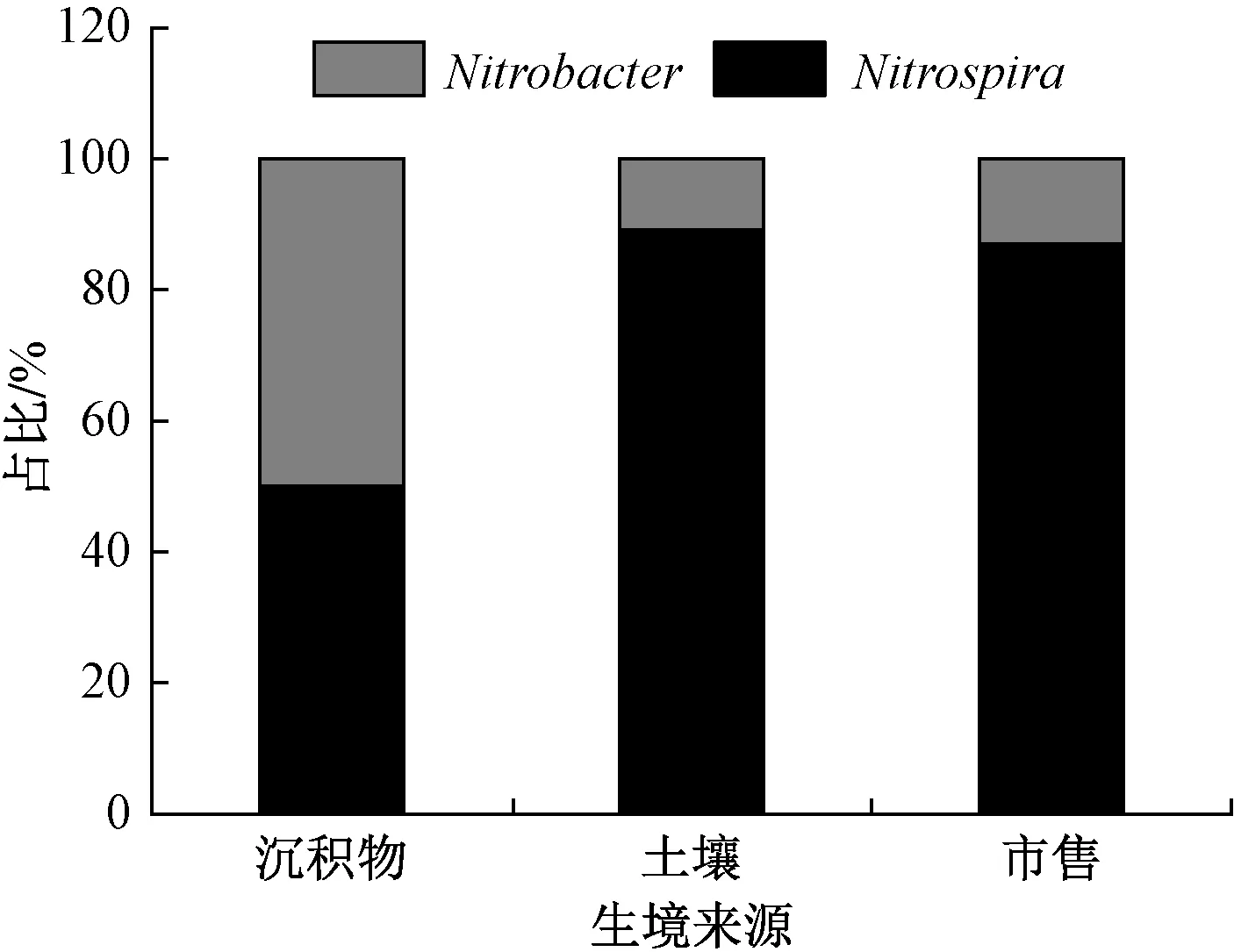
圖7 Nitrobacter 和Nitrospira 在硝化細菌中所占比例Fig.7 Percentages of Nitrobacter and Nitrospira in nitrifying bacteria
2.4 AOB 的系統發育分析
將測得的amoA基因序列構建NJ(Neighbor-Joining)系統樹,如圖8 所示。從圖8 可以看出,沉積物、土壤和市售生境AOB 均屬于Nitrosospira和Nitrosomonas,符合二者具有廣布性的特點[35]。土壤Nitrosospira和Nitrosomonas主要集中在系統樹中的Cluster 1 和Cluster 7,而沉積物和市售生境AOB 在系統樹中的位置較為分散。這表明土壤生境AOB 的特異性較高,即AOB 多樣性低于沉積物和市售生境,這可能是由生存環境穩定性差異引起的。由于上覆水水質頻繁波動的影響,沉積物生境硝化細菌的生存環境變異性要高于土壤生境,導致沉積物生境AOB 多樣性較高;市售生境硝化細菌的多樣性較高,有可能是其最初來源的自然生存環境條件的不穩定所致。雖然Nitrosospira和Nitrosomonas均存在于沉積物、土壤和市售等生境中,但其在各生境AOB 的相對組成上存在差異(圖9)。在沉積物生境中,Nitrosomonas是AOB 的主要組成部分,約占81.6%,為優勢屬,Nitrosospira僅占18.4%;在土壤和市售生境AOB 組成中,Nitrosospira與Nitrosomonas所占比例相當,分別占AOB 生物量的47.1%和52.9%、49.0%和51.0%。有報道稱[36],Nitrosomonas能耐受較低濃度的氧氣,即使在厭氧條件下,Nitrosomonas仍可達到總細菌生物量的1/4,這可能是在缺氧的沉積物環境中,Nitrosomonas在AOB 中占比高于土壤和市售生境AOB 的主要原因。

圖8 基于amoA 基因序列所構建的NJ 樹Fig.8 NJ tree of amoA genes
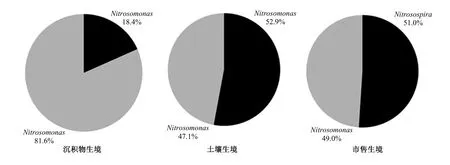
圖9 amoA 基因克隆子中Nitrosospira 和Nitrosomona 占比Fig.9 The abundances of Nitrosospira and Nitrosomona in amoA gene clones
2.5 NOB 的16S rRNA 基因序列
將Nitrobacter和Nitrospira所測得的16S rRNA基因克隆子序列進行基因型分析,結果如表1 和表2所示。Nitrobacter基因型多樣性較高,114 個克隆子基因型可分為51 種,其中沉積物生境39 個克隆子有18 種基因型,土壤生境38 個克隆子有18 種基因型,市售生境37 個克隆子有21 種基因型。這表明在沉積物、土壤和市售這3 種生境中Nitrobacter多樣性相似,種類較多。基因型1、2、3 的Nitrobacter在這3 種生境中均存在,為廣布菌,其中基因型1 的Nitrobacter為優勢菌,克隆子數約占總克隆子數的28.1%,基因型2 的Nitrobacter為次優勢菌,占比為13.2%。沉積物、土壤和市售生境Nitrobacter結構組成上存在一定的差異。沉積物生境中,基因型1 和基因型2 的Nitrobacter占主要優勢;土壤和市售生境中,多種基因型的Nitrobacter并存。沉積物、土壤和市售生境中Nitrospira多樣性較低,94 個克隆子僅有8 種基因型。基因型1 占比最高,達91.5%,并且在3 種生境中均廣泛存在。這表明,基因型1 細菌是沉積物、土壤和市售生境中Nitrospira的廣布菌和絕對優勢菌,相比于其他基因型的Nitrospira,它在硝化反應中可能起著最為重要的作用。綜上,3 種生境的Nitrospira種群結構相似,都是由1 種主要的細菌(基因型1)和少量其他種類的細菌組成。

表1 各生境中Nitrobacter 的基因型分析Table 1 Genotype analysis of Nitrobacter from different habitats
硝化細菌的種群組成和豐度影響著硝化速率的大小[37]。在眾多生境中,NOB 的生物量是AOB 的3~30 倍,這是因為NOB 的底物是AOB 的產物,NOB 在生物量上的優勢能夠保證硝化反應的順利進行,避免由于積累對硝化細菌帶來的毒害作用[38-39]。本研究中,3 種生境AOB 生物量均高于NOB,這可能與實驗室內硝化細菌的富集培養條件與其自然生存環境條件的不同有關。在實驗室內富集時,硝化反應產物不能即時消除而不斷積累,對NOB 的生長產生抑制作用,造成NOB 生物量低于AOB;而在硝化細菌自然生存環境中,能通過反硝化過程和水生生物直接吸收等途徑來消除。因此,在本試驗中,由于生物量的不均衡,NOB 對各反應器的硝化效率起著決定性影響。大量研究表明[40-41],Nitrospira是污水處理系統、生物反應器以及淡水養殖塘中起主要作用的NOB。對Nitrospira克隆子基因型分析已知(表2),沉積物、土壤和市售生境Nitrospira組成結構相似,即由1 種優勢菌和少量其他種類硝化細菌組成,但生物量存在差異,即沉積物生境Nitrospira的生物量高于土壤和市售生境。有可能是該原因導致沉積物生境硝化細菌的硝化速率要高于土壤和市售生境,因此,在處理人工配制污水和實際生活污水時,沉積物生境硝化細菌的硝化速率明顯高于土壤和市售生境(圖1)。
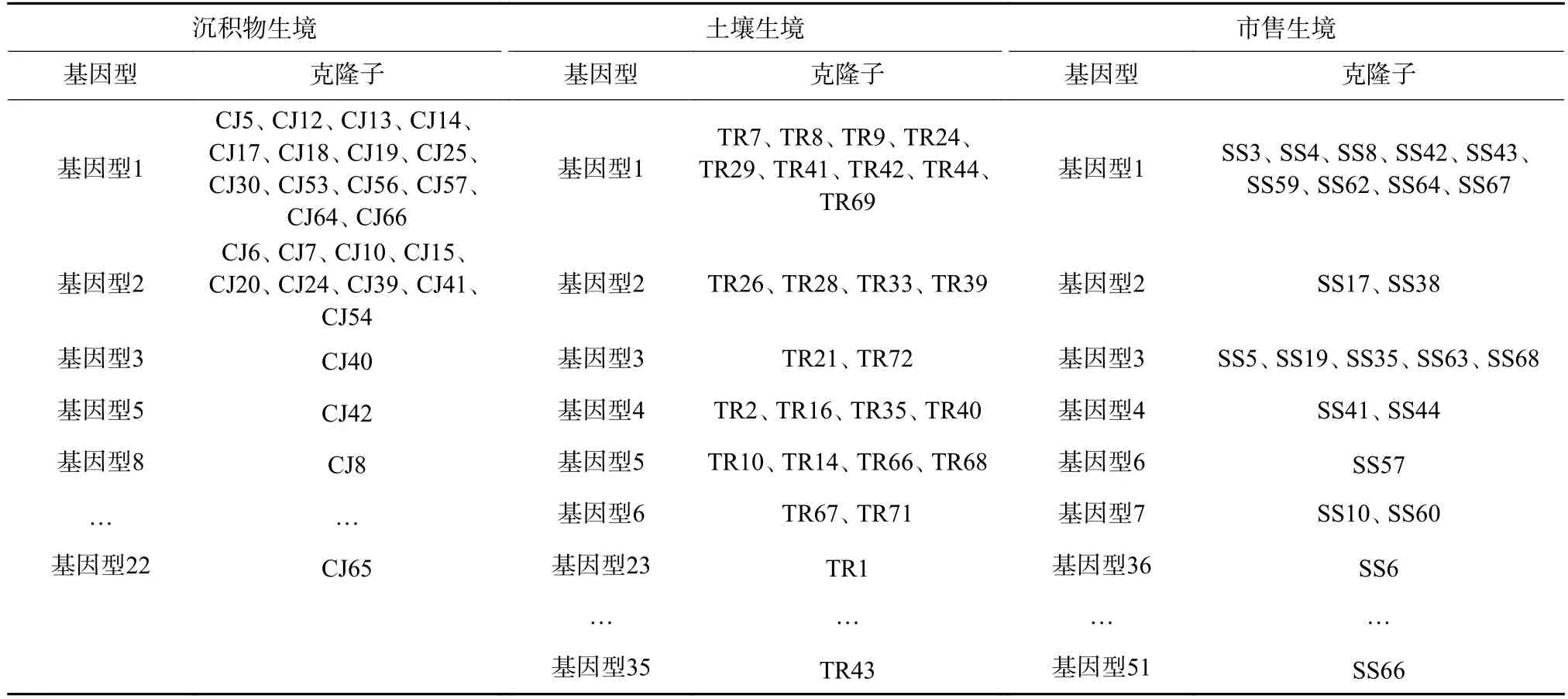
表2 各生境中Nitrospira 基因型分析Table 2 Genotype analysis of Nitrospira from different habitats
Nitrospira對游離氨的敏感性高于Nitrobacter[42]。當氨氮初始濃度較高時,游離氨濃度隨之升高,Nitrospira活性受到抑制。此時,Nitrobacter取代Nitrospira成為硝化過程中起關鍵作用的細菌,這就意味著Nitrobacter的種群組成和豐度影響著整個硝化反應的速率。由于土壤生境Nitrobacter生物量明顯高于沉積物和市售生境,理論上,土壤生境硝化細菌的硝化速率受氨氮初始濃度影響較小,即耐受氨氮的性能較好。但實際上,沉積物生境硝化細菌耐受氨氮的能力高于土壤和市售生境(圖3)。這可能是由各生境中Nitrobacter種群組成不同引起的。在沉積物生境中,基因型1 和基因型2 的Nitrobacter占比高于土壤和市售生境(表1)。據此推測,基因型1 和基因型2 的Nitrobacter在整個Nitrobacter屬中起著主要的作用。利用Blast 程序進行序列比對得出,基因型1 和基因型2 的Nitrobacter均為不可培養微生物,需采取其他方法做進一步研究。pH 主要通過引起硝化細菌的活化與鈍化,對無機碳源的影響,借助游離氨的抑制作用等多種途徑來影響硝化作用,但通過游離氨的抑制作用被認為是最主要的途徑[29]。沉積物生境硝化細菌對游離氨耐受能力高于土壤和市售生境,相應地,沉積物生境硝化細菌對pH 的耐受能力也高于土壤和市售生境,這與pH 對各生境硝化細菌影響的試驗結果(圖4)相一致。
在特殊環境條件的長期影響下,淡水水體中的部分硝化細菌種類具有耐受高鹽度的能力。以AOB 為例,人們已從污水處理廠和富營養化淡水中分離出耐鹽或適度嗜鹽的Nitrosomonas europaea和N.eutropha[43]。對amoA基因序列進行系統發育分析發現(圖8),土壤生境硝化細菌種群中存在耐鹽或適度嗜鹽的Nitrosomonas europaea和不耐鹽的Nitrosomonasoligotropha,沉積物和市售生境硝化細菌種群中只鑒定出不耐鹽的Nitrosomonas oligotropha,而沉積物生境硝化細菌的耐鹽性高于土壤和市售生境(圖5)。可能是沉積物生境硝化細菌種群中存在更為豐富的不可培養耐鹽度種類,需利用其他生物學方法做進一步分析。
3 結論
(1)沉積物生境硝化細菌的硝化速率高于土壤和市售生境,這主要是因為沉積物生境中Nitrospira生物量較高。
(2)沉積物生境硝化細菌耐受氨氮、pH 和鹽度等環境因子的能力高于土壤和市售生境,這是因為沉積物生境硝化細菌群落中存在Nitrobacter和可能存在耐鹽或適度嗜鹽種類(有待利用其他生物學方法進一步分析、確定)。
(3)微生物脫氮工藝所需的硝化細菌可考慮從沉積物中富集、篩選,以獲取硝化效率高且環境適應強的菌種。
