新型麥胚活性肽對脂多糖誘導RAW264.7細胞的抗炎作用
2022-02-20 07:45:04陳元蓉楊高紅沈新春宋海昭王鑾鳳
中國糧油學報 2022年12期
陳元蓉,張 宸,楊高紅,沈新春,宋海昭,熊 玲,王鑾鳳,汪 芳
(南京財經大學食品科學與工程學院;江蘇省現代糧食流通與安全協同創新中心;江蘇高校糧油質量安全控制及深加工重點實驗室;糧食儲運國家工程實驗室,南京 210023)
巨噬細胞是調控炎癥反應的中心細胞,在炎癥的發生、維持和解決中起著關鍵作用[1]。在炎癥發生過程中,巨噬細胞主要通過抗原呈遞、吞噬和產生各種細胞因子和生長因子進行免疫調節。巨噬細胞對內毒素脂多糖(LPS)的識別激活了下游細胞內信號級聯,導致多種炎癥介質和細胞因子的產生[2],例如TNF-α、IL-1β、IL-6、IL-8和IL-12等[3]。TNF-α、IL-1β和IL-8通過協同誘導其他細胞因子、趨化因子和黏附分子的產生來放大炎癥反應。IL-6與其受體結合后,通過參與抗凋亡基因的表達,從而防止T細胞凋亡,最終擴大了炎癥反應。這些促炎細胞因子的mRNA表達在炎癥反應中被上調。然而,IL-10作為抗炎細胞因子,可以抑制促炎細胞因子的合成,從而調節炎癥反應[4]。
近年來,越來越多的研究表明食源性多肽具有良好的抗炎活性,其比藥物的副作用低,且呈現出對靶組織的高特異性,具有低毒高效的特點,其通過調節細胞因子的釋放,從而減輕炎癥細胞因子誘導的炎癥,在治療慢性炎癥性疾病方面具有明顯的優勢[5, 6, 7]。Hao等[8]從大豆中分離得到的Lunasin肽在LPS誘導的RAW 264.7 巨噬細胞炎癥模型中顯示出較高的抗炎活性,并顯著抑制了炎癥細胞因子的分泌。Gao等[9]研究發現鱘魚蛋白衍生的3種肽在LPS刺激的RAW 264.7巨噬細胞炎癥模型中,可有效抑制炎癥細胞因子IL-1β和IL-6的釋放。Ji等[10]研究表明小米谷醇溶蛋白肽(MPP)通過抑制LPS刺激的RAW 264.7巨噬細胞中炎癥細胞因子TNF-α、IL-6和IL-1β的產生,進而顯著減輕炎癥反應。Ren等[11]研究發現來源于榛子蛋白酶水解物的多肽LDAPGHR在LPS誘導的RAW264.7細胞炎癥模型中表現出抗炎作用,其通過抑制IL-1β、IL-6和TNF-α的分泌量來發揮抗炎作用。隨著越來越多的食源性多肽的抗炎活性被發現,從食品中尋找發現新型食源性抗炎活性肽將對炎癥性疾病具有重要意義。
小麥胚芽中麥胚蛋白約占30%,是一種完全蛋白,含有人體必需的8種氨基酸,被稱為“人類天然營養寶庫”[12]。生物活性肽通常由3~20個氨基酸組成,在母體蛋白質中不起作用。然而,它們可以通過酶水解、發酵或食品加工等手段得以釋放。有研究表明,小麥胚芽可提高動物對急性和慢性炎癥的抵抗作用,并通過抑制血清中促炎細胞因子的分泌恢復良好的腸道抗炎環境,從而治療佐劑性關節炎[13]。Jeong等[14]研究發現,檸檬酸處理的小麥胚芽提取物(CWG)在LPS刺激的巨噬細胞模型中顯示出良好的抗炎作用,其通過抑制促炎細胞因子TNF-α的分泌、白細胞介素IL-6、IL-12和COX-2的合成來發揮抗炎作用。雖然已有研究通過酶解、發酵等手段得到麥胚活性肽[15],但通過體外模擬消化來得到麥胚抗炎活性肽鮮見報道,內肽酶水解可提高活性肽的生物利用度,避免其在胃腸道中進一步被消化。此外,胃腸道酶消化比單一酶消化產生的多肽效果更好[16]。基于此,本研究利用體外模擬胃腸道消化對麥胚蛋白進行酶解,通過超濾膜分離技術對蛋白酶解產物進行逐級分離,根據各級分離產物的抗炎活性篩選出具有較高抗炎活性的目標肽段,鑒定其氨基酸序列,并結合PeptideRanker數據庫活性預測結果篩選出抗炎活性最高的肽序列,進一步采用LPS誘導的RAW 264.7細胞體外炎癥模型檢測其抗炎活性,以期為麥胚活性肽的發現與分離純化提供研究基礎,并為麥胚蛋白應用于抗炎活性肽的開發與利用提供參考。
1 材料與方法
1.1 材料與試劑
小麥胚芽;小麥胚芽肽純度>98%。胰蛋白酶(2 500 U/mg)、胃蛋白酶(3 000 U/mg)、胰凝乳蛋白酶(250 U/mg);NO試劑盒和ELISA試劑盒(TNF-α、IL-1β、IL-6和IL-10);RAW 264.7細胞株;噻唑藍(MTT);胎牛血清(FBS)和RPMI Medium 1640培養基;地塞米松(DEX);所有分離用有機溶劑均為分析純。
1.2 儀器與設備
Labscale小型切向流超濾系統,H1850高速冷凍離心機,Thermo Scientific酶標儀,2XZ 旋片真空泵,MX-RD-E標準型旋轉混均儀,MZE多功能酶標儀,ZDDN-II全自動凱氏定氮儀器,Axio Observer A1熒光倒置顯微鏡,Easy-nLC 1200高效液相色譜儀和Q Exactive高分辨質譜儀,SCIENTZ-10N-A冷凍干燥機。
1.3 方法
1.3.1 麥胚蛋白的制備
小麥胚芽采用正己烷進行脫脂。將脫脂后的小麥胚芽,用粉碎機粉碎后過100目篩。稱取50.0 g小麥胚芽粉加入500 mL蒸餾水,稱取5.0 g NaCl用少量水溶解加入溶液中,調pH至9.0,用磁力攪拌器攪拌提取30 min,再于4 ℃,5 000 r/min離心10 min,取上清液,然后調pH至4.0,待沉淀完全后,二次離心10 min,棄去上清液,取沉淀,用蒸餾水溶解并調pH至7.0,透析脫鹽后,真空冷凍干燥得麥胚蛋白粉,于-20 ℃保存,備用[17]。參考GB 5009.5—2016《食品安全國家標準 食品中蛋白質的測定》,采用凱氏定氮法測定麥胚蛋白粉中的蛋白含量,小麥胚芽轉換系數為5.8。
1.3.2 麥胚蛋白體外模擬胃腸消化
根據Sangsawad等[18]的方法對模擬胃腸消化過程略做修改,用去離子水溶解麥胚蛋白凍干粉,攪拌均勻,形成8%的蛋白溶液,95 ℃水浴加熱30 min,調pH至7.0后使蛋白質完全變性。在模擬胃消化階段,用1 mol/L HCl溶液將pH保持在2.0,在37 ℃下用胃蛋白酶(0.4%,干基)消化懸浮液4 h。在模擬腸道消化階段,用1 mol/L NaOH溶液將pH保持在7.6,在37 ℃下用胰蛋白酶(0.3%,干基)和α-胰凝乳蛋白酶(0.1%,干基)消化6 h。消化過程中每隔1 h采樣,測定樣品水解度(DH)。消化10 h后,待消化液冷卻至室溫,5 000 r/min離心20 min,收集上清液,透析脫鹽后,于4 ℃下保存待進一步分離純化。
1.3.3 水解度測定
標準曲線的繪制:分別取0、100、200、300、400 μL絲氨酸標準液于5 mL試管中,加入去離子水補足至400 μL,加入3 mL OPA試劑,混勻,反應2 min,測定吸光度(以去離子水做參比)。以吸光度為縱坐標Y,絲氨酸濃度為橫坐標X,繪制標準曲線。
水解度的測定:取不同時間點的水解液1 mL,用去離子水稀釋100倍(使測定值在標準曲線范圍內),取稀釋后的水解液400 μL加入3 mL OPA試劑,反應2 min,測定吸光度。在同一條件下重復測定3次,根據測得的吸光度平均值從標準曲線上計算出CSerine-NH2(mmol/L),按公式計算水解度[19]。
式中:Wserine-NH2為每克蛋白質中含serine-NH2的量/mmol/g;x為樣品質量/g;P為樣品中蛋白質質量分數/%;V為樣品水解液體積/L;N為水解液稀釋倍數;h為水解過程中每克麥胚蛋白被斷裂的肽鍵數/mmol/g prot;htot為每克麥胚蛋白質所含的總肽鍵數(8 mmol/g);α、β分別用常數1.00、0.40表示。
1.3.4 雙縮脲法測定多肽含量
標準曲線的繪制:用5%的TCA依次配制0.0、0.2、0.4、0.6、0.8、1.0、1.2、1.4、1.6和1.8 mg/mL的Gly-Gly-Tyr-Arg四肽標準溶液10 mL。分別取6.0 mL不同質量濃度的標準溶液,加入4.0 mL雙縮脲試劑,混合均勻,靜置10 min,2 000 r/min離心10 min,取上清液于540 nm下測定OD值。以四肽標準溶液的質量濃度為橫坐標X(mg/mL),OD值為縱坐標Y,繪制標準曲線[20]。
取2.5 mL樣品溶液,加入2.5 mL質量分數10%的三氯乙酸(TCA)水溶液,于漩渦混合儀上混合均勻,靜置10 min,4 000 r/min離心15 min,將上清液全部轉移至50 mL容量瓶中,并用5%的TCA定容,搖勻。取6.0 mL溶液于15 mL離心管中,加入4.0 mL雙縮脲試劑(樣液:雙縮脲試劑體積比=3∶2),于漩渦混合儀上混合均勻,靜置10 min,2 000 r/min離心10 min,取上清液于540 nm下測定OD值,對照標準曲線求得樣品溶液中的多肽質量濃度C(mg/mL),計算樣品中多肽含量。通過四肽標品的測定,得到標準曲線方程為y=0.108 9x+0.040 4,R2=0.992 6。
1.3.5 超濾膜分離
選用分子截流量為3 ku和1 ku的超濾膜,采用Labscale小型切向流超濾系統對酶解物進行分離,分別得到3個組分WGPH-Ⅰ(>3 ku)、WGPH-Ⅱ(1~3 ku)和WGPH-Ⅲ(<1 ku)。具體操作步驟:用0.1 mol/L,pH 7.6的KH2PO4-K2HPO4緩沖液將樣品配成質量濃度為0.1%的溶液,用0.45 μm的纖維素膜過濾除去不溶物。調節泵的壓力控制流速并在4 ℃下進行超濾。將超濾后不同分子量的組分脫鹽并冷凍干燥,得到不同分子量的多肽組分,用于下一步的抗炎活性篩選。
1.3.6 Nano-LC-MS/MS測多肽序列
將肽粉溶于200 μL Nano-HPLC Buffer A中,混勻離心提取上清液,將上清液轉入0.22 μm旋轉過濾器中,收集濾液[21]。
制備C18膜填充柱;將揮干的多肽樣品重新溶解于Nano-HPLC Buffer A中,然后用40 μL甲醇過柱離心1遍,棄掉EP管底部液體,重復2次;40 μL Nano-HPLC Buffer A過柱離心1遍,棄掉EP管底部液體,重復2次;多肽樣品過柱離心1遍,取EP管底部液體再過柱1遍;40 μL Nano-HPLC Buffer A過柱離心1遍,棄掉EP管底部液體,重復2次;更換新的EP管,40 μL 洗脫相Buffer B過柱離心1遍,收集EP管底部液體,重復1次。脫鹽后將含多肽樣品的80 μL 洗脫相Buffer B進行揮干。
將揮干的多肽樣品重新溶解于Nano-HPLC Buffer A中。采用Nano-HPLC液相系統EASY-nLC1200進行分離。液相A液為0.1%甲酸-水溶液,B液為0.1%甲酸-乙腈溶液。100 μm×20 mm RP-C18色譜柱Trap column以100%的A液平衡。樣品由自動進樣器上樣并吸附到Trap column柱上,再經分析柱,75 μm×150 mm RP-C18色譜柱分離,流速為300 nL/min。樣本間用空白溶劑30 min流動相梯度清洗1次。酶解產物經毛細管高效液相色譜分離后用Q-Exactive質譜儀進行質譜分析。檢測方式為,使用前經標準校正液校正,母離子掃描范圍:350~2 000m/z,質譜掃描方式為信息依賴的采集工作模式下(DDA),每次全掃描后采集最強的20個碎片圖譜(MS2 scan),碎裂方式為高能碰撞解離(HCD),NCE能量28,動態排除時間25 s。MS1在M/Z 200時分辨率為70 000,AGC target設置為3e6,最大注射時間100 ms,MS2分辨率設置為17 500,AGC target設置為1e5,最大注射時間50 ms。
1.3.7 多肽合成
利用高效固相肽合成儀合成抗炎肽,并通過反相高效液相色譜(RP-HPLC)質譜(LC-MS/MS)聯用對其純度進行了驗證,合成的肽在使用前保存在-20 ℃。
1.3.8 抗炎活性的測定
1.3.8.1 噻唑藍(MTT)法檢測RAW 264.7增殖活力
取對數生長期的RAW 264.7巨噬細胞,用10% FBS RPIM 1640培養液配制成3×104個/mL的細胞懸浮液100 μL于96孔板中,在細胞培養箱中培養12 h使細胞貼壁,另設置無細胞孔作為空白組。除去96孔板中培養液后,將多肽溶解于含10% FBS的RPIM 1640培養液中制成不同質量濃度多肽(20、80、320 μg/mL)培養液,再加到96孔板中,置于細胞培養箱中,37 ℃培養24 h后每孔加入10 μL的MTT溶液(5 mg/mL),培養4 h后吸去96孔板中溶液,加入50 μL的DMSO溶液,37 ℃恒溫振蕩20 min后在570 nm下測吸光度[22]。
1.3.8.2 Griess方法檢測LPS誘導的RAW264.7中NO含量
同1.3.8.1所述方法培養RAW 264.7巨噬細胞,加入不同質量濃度多肽(20、80、320 μg/mL)預保護2 h,然后加入LPS(1 μg/mL),于培養箱中繼續恒溫(37 ℃)培養24 h。從96孔板各孔中吸取50 μL上清液分別轉移到另一個96孔板各孔中,每孔依次加入NO檢測試劑盒中的Griess Ⅰ和Griess Ⅱ溶液各50 μL,混勻后。在540 nm下的檢測吸光度值,換算成NO摩爾濃度[23]。
1.3.8.3 炎癥因子的測定
根據試劑盒說明書使用ELISA試劑盒(IL-1β、IL-6、IL-10和TNF-α)檢測炎癥因子含量[24]。
1.3.9 統計學分析
所有實驗重復3次,實驗數據以平均值±標準差表示,數據分析使用SPSS17.0軟件進行student-t檢驗,P<0.05表示數據具有統計學差異。
2 結果與分析
2.1 麥胚蛋白體外模擬消化過程中水解度的變化
測得小麥胚芽中蛋白質質量分數為(28.47±2.65)%,其純度為(82.49±3.97)%。水解度(DH)表示蛋白質的降解程度,目前在評價水解效率方面應用廣泛。如圖1所示,麥胚蛋白在體外模擬消化階段其水解度整體呈上升趨勢;在胃消化階段(0~4 h),麥胚蛋白經胃蛋白酶水解后,其DH最終達到6.87%。有研究表明蛋白質經過胃蛋白酶的初步水解后其產物一般為分子量較大的多肽[25]。在腸道消化階段(4~10 h),DH在前2 h加速上升,到達11.99%并趨于穩定,其水解度最終為13.08%。小腸是人體的主要營養物質吸收部位,其中胰蛋白酶和α胰凝乳蛋白酶發揮著重要作用。胰蛋白酶作為內肽酶,可以沿底物蛋白的一級結構在氨基酸鏈的中間切割肽鍵。因此胰酶的加入大幅度提高了麥胚蛋白的DH,并產生大量的小分子活性肽。此外,由于消化液濃度的降低以及蛋白質底物和酶切位點的減少導致DH逐漸趨于穩定。

圖1 麥胚蛋白在不同消化時間的水解度的變化
2.2 超濾膜分離和多肽含量
超濾分離是分離提純的過程,其根據物質之間相對分子質量的不同達到分離的目的。本實驗利用超濾膜對麥胚消化酶解液進行分離純化,以去除分子量大于3 ku的多肽雜質,最終得到純度較高的多肽溶液。最終分別得到3個組分WGPH-Ⅰ(>3 ku)、WGPH-Ⅱ(1~3 ku)和WGPH-Ⅲ(<1 ku),真空冷凍干燥得到凍干粉后測得3個組分的多肽質量分數分別為(4.03±0.33)%、(4.27±0.24)%和(5.68±0.06)%。不同組分肽段的含量、顏色和澄清度均不相同。大于3 ku超濾膜分離的肽段組分顏色最深,偏黃色,可能含有一些小分子物質存在。其他組分顏色為黃白色或白色,低分子量多肽的澄清度高于高分子量的澄清度。
2.3 麥胚抗炎活性肽的篩選
2.3.1 麥胚多肽(WGPHs)對RAW 264.7細胞活力的影響
通過MTT實驗探究麥胚多肽WGPH-Ⅰ(>3 ku)、WGPH-Ⅱ(1~3 ku)和WGPH-Ⅲ(<1 ku)在不同濃度下對巨噬細胞活力的影響,結果如圖2所示,與對照組相比,不同組分對RAW 264.7細胞的活力產生了一定的影響。當質量濃度在20~320 μg/mL時,3個組分對RAW 264.7細胞活力均無顯著影響(P>0.05),表明其在20~320 μg/mL質量濃度下對巨噬細胞無毒性。而當質量濃度達到640 μg/mL時,細胞活力顯著降低(P<0.01),表明過高濃度的麥胚多肽對細胞毒性較大。因此,綜合細胞活力實驗結果,選擇20、80、320 μg/mL 3個質量濃度進行后續實驗,進一步探究其抗炎作用。
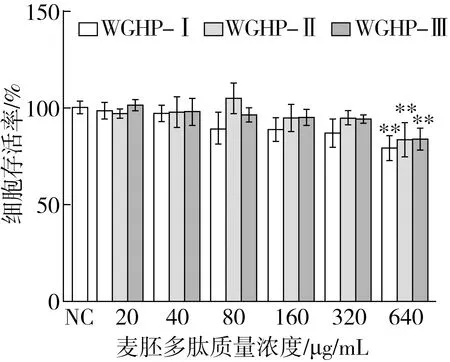
注:**表示與LPS模型組相比具有極顯著差異(P<0.01)。
2.3.2 麥胚多肽(WGPHs)對LPS誘導的RAW 264.7細胞NO分泌量的影響
NO是一種重要的跨膜分子信號,能夠損傷機體周圍組織,當炎癥發生時,巨噬細胞會分泌大量NO。已有報道表明LPS刺激RAW 264.7細胞可以急劇增加NO的釋放量,造成細胞產生炎癥反應[26]。因此,本研究采用LPS誘導的巨噬細胞炎癥模型分析不同多肽組分的抗炎活性。通過Griess法檢測麥胚多肽(WGPHs)對LPS誘導的巨噬細胞NO分泌量的影響。結果如圖3所示,與空白組相比,LPS誘導的RAW 264.7細胞NO分泌量明顯升高(P<0.01),其釋放量增加了5.98倍,表明該模型成功。麥胚多肽WGPH-Ⅰ和WGPH-Ⅱ干預組NO濃度與模型組相比無顯著差異;WGPH-Ⅲ干預組則顯著抑制了NO的釋放,表現出較好的抗炎活性。WGPH-Ⅲ組分的分子量(<1 ku)低于WGPH-Ⅰ和WGPH-Ⅱ組分,表明分子量較小的多肽其抗炎活性可能更高。該結果與Torresfuentes等[27]的研究相似。因此,選擇WGPH-Ⅲ組分進行后續質譜鑒定以及抗炎活性的進一步驗證。

注:##表示與正常組相比有極顯著差異(P<0.01),*表示與LPS模型組相比具有顯著差異(P<0.05),**表示與LPS模型組相比具有極顯著差異(P<0.01),下同。
2.4 Nano-LC-MS/MS鑒定WGPH-Ⅲ(<1 ku)多肽序列
采用Nano-LC-MS/MS對WGPH-Ⅲ的多肽組分進行結構鑒定,其色譜主要出峰時間集中在7~90 min,對應的流動相中乙腈質量分數為50%~67%,表明WGPH-Ⅲ多肽呈現非極性,推測多肽中可能含有一定量疏水性氨基酸。
通過Mascot 2.3軟件分析得到與小麥胚芽蛋白質組中同源性最高的20個短肽。從350~2 000m/z中,選取峰面積前14的碎片進行二級質譜分析,共鑒定出14條多肽,其氨基酸序列及分子量如表1所示,這些肽均由7~12個氨基酸組成,且觀察肽序列可以看出Phe(F)、His(H)、Trp(W)、Cys(C)和Tyr(Y)等小分子疏水氨基酸殘基高頻率出現,說明其生物活性好。通過Peptide Ranker數據庫(http://distilldeep.ucd.ie/Peptide Ranker/)對每個肽進行評分,評價肽的生物活性概率。從預測結果可以得出,疏水氨基酸多且分子量小的肽活性較強。其中APEPEPAF(P1)、MDTSAPSPF(P2)、IDIPNGR(P3)和VDPAVPPK(P4)序列活性較強,因此,通過高效固相肽合成儀合成4條多肽,進一步驗證其生物活性,其二級質譜圖見圖4。
圖4 肽段的二級擬合質譜圖
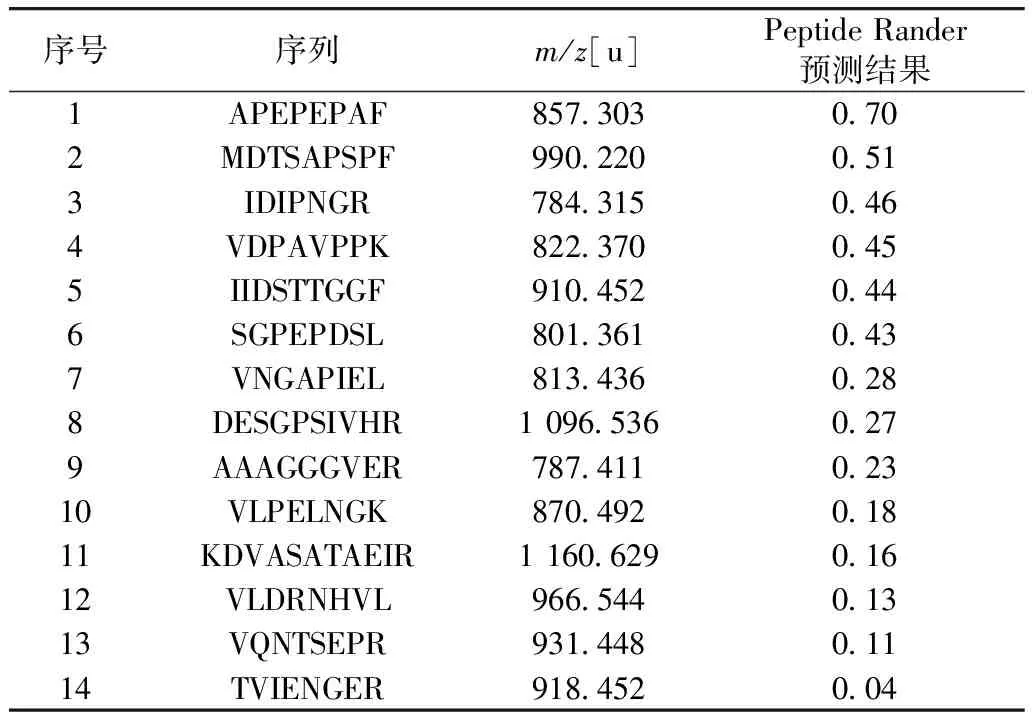
表1 目標肽段的鑒定及生物活性的預測
2.5 小肽P1、P2、P3和P4分別對LPS誘導的巨噬細胞NO分泌量的影響
通過Griess法檢測小肽P1、P2、P3和P4對LPS誘導的巨噬細胞NO分泌量的影響,結果如圖5所示,與空白組相比,模型組的RAW 264.7細胞NO分泌量極顯著升高(P<0.01)。與模型組相比,小肽組的NO濃度均明顯降低,且隨著濃度的增加,NO的分泌量逐漸降低,表明P1、P2、P3和P4都可以抑制RAW 264.7細胞NO的分泌,表現出較強的抗炎活性,且具有劑量依賴性。其分子機制可能是LPS激活巨噬細胞,誘導局部炎癥和抗體的產生,并影響體內iNOS的表達,從而影響NO的產生。其中小肽P1的抗炎活性最好,在炎癥性疾病的治療或預防中具有潛在意義。因此,選取P1(APEPEPAF)進行下一步研究。

圖5 小肽P1、P2、P3、P4對LPS誘導的巨噬細胞NO分泌量
2.6 小肽P1(APEPEPAF)對LPS誘導的巨噬細胞炎癥介質的影響
LPS是革蘭氏陰性細菌細胞壁的主要成分,能與Toll樣受體4(TLR4)結合,進而誘導巨噬細胞產生炎癥。當機體發生炎癥時,LPS的生成量會隨革蘭氏陰性菌數量增加而增加,影響炎癥性疾病的發生,且長期的炎癥會成為腫瘤發展的主要誘因[28]。經LPS刺激后,巨噬細胞會合成或釋放炎癥因子,阻斷炎癥因子是抑制炎癥的一個有效治療策略。采用Elisa試劑盒測定LPS誘導的巨噬細胞炎癥介質(IL-6、IL-10、TNF-α和IL-1β)的分泌量,并分析小肽P1對炎癥因子水平的影響,結果如圖6所示。
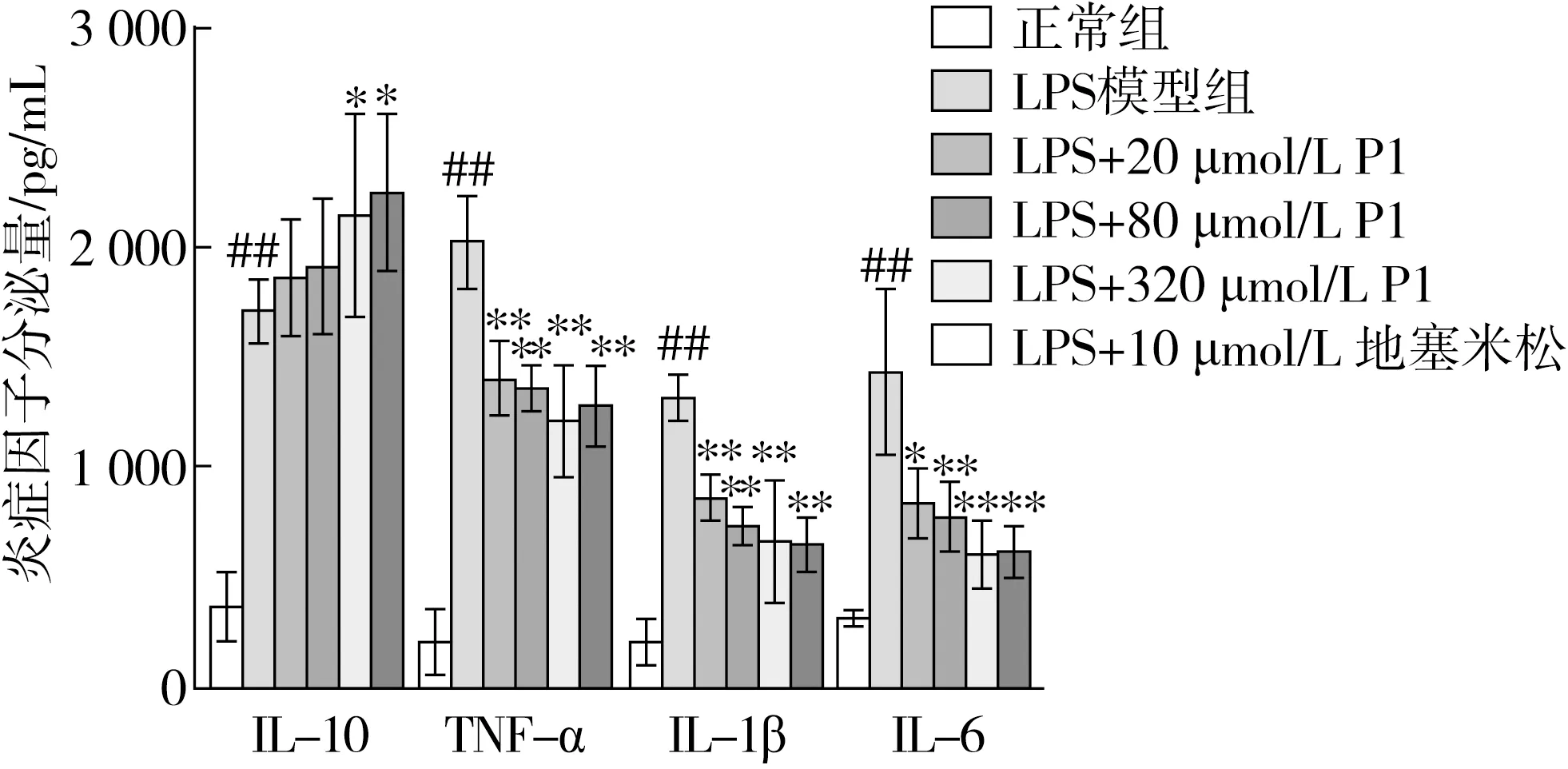
圖6 小肽APEPEPAF對LPS誘導的RAW264.7分泌炎癥因子的影響
與正常組相比,LPS誘導細胞后,其促炎因子IL-6、TNF-α和IL-1β的分泌顯著升高(P<0.05)。當小肽P1或陽性藥DEX干預后,與LPS組相比,促炎因子IL-6、TNF-α和IL-1β的分泌顯著得到抑制(P<0.01)。此外,與正常組相比,LPS誘導巨噬細胞后,其抗炎因子IL-10的分泌顯著升高(P<0.01),小肽P1或陽性藥DEX干預后均在不同程度上增加了抗炎因子IL-10的分泌,隨著小肽P1干預濃度的增加,其分泌量呈現出上升趨勢(P<0.01)。
通過檢測炎癥因子分泌水平的結果表明,小肽P1對促炎因子IL-1β、IL-6和TNF-α的分泌具有抑制作用,對抗炎因子IL-10的分泌有一定的促進作用。即小肽P1能夠抑制LPS誘導的巨噬細胞促炎因子IL-1β、IL-6和TNF-α的分泌,促進抗炎因子IL-10的分泌,呈現出良好的改善炎癥的作用效果,表明小肽干預后能在蛋白表達和基因轉錄水平上抑制細胞炎癥因子的表達,降低機體炎癥反應,從而發揮抗炎作用。
已有研究表明,多肽的抗炎活性與其氨基酸組成存在密切關系,Saisavoey等[29]和Zhao等[30]研究發現含有P、F殘基的多肽具有抗炎活性;此外,Liang等[31]發現玉米多肽—PPYLSP和FLPPVTSMG也呈現出較強的抗炎活性。由此推測,含有P、F、L等氨基酸殘基的多肽呈現出較好的抗炎活性;本研究中小肽P1(APEPEPAF)、P2(MDTSAPSPF)、P3(IDIPNGR)和P4(VDPAVPPK)的氨基酸序列中含有P、F等氨基酸殘基,其中小肽P1的P、F氨基酸殘基最多,且抗炎活性最強,小肽P1由于含有P、F等氨基酸殘基而呈現出較好的抗炎活性,說明小肽P1在預防或治療炎性疾病方面具有很大的應用潛力,可以作為一種新型的抗炎藥物用于疾病的預防和治療。
3 結論
采用堿溶酸沉法、體外模擬胃腸道消化法、超濾膜分離以及PeptideRanker數據庫活性預測,從麥胚蛋白消化產物中分離篩選得到14條新的生物活性肽,并進一步通過脂多糖(LPS)誘導的巨噬細胞RAW 264.7炎癥模型,篩選得到抗炎活性較強的4條抗炎活性肽,P1(APEPEPAF)、P2(MDTSAPSPF)、P3(IDIPNGR)和P4(VDPAVPPK),均能降低LPS誘導的巨噬細胞中NO的分泌量。其中抗炎活性最強的小肽P1,其氨基酸序列為APEPEPAF,分子量為856 u,該小肽能夠顯著抑制脂多糖誘導的巨噬細胞中NO及促炎因子的分泌,說明麥胚蛋白可作為抗炎肽的前體,通過內肽酶的酶解得以釋放,且安全性強、穩定性高,可以作為一種潛在的功能性食品加以開發利用。
